
Abstract
Clinically normal females exhibit higher 18F-flortaucipir (FTP)-PET signal than males across the cortex. However, these sex differences may be explained by neuroimaging idiosyncrasies such as off-target extracerebral tracer retention or partial volume effects (PVEs). 343 clinically normal participants (female = 58%; mean[SD]=73.8[8.5] years) and 55 patients with mild cognitive impairment (female = 38%; mean[SD] = 76.9[7.3] years) underwent cross-sectional FTP-PET. We parcellated extracerebral FreeSurfer areas based on proximity to cortical ROIs. Sex differences in cortical tau were then estimated after accounting for local extracerebral retention. We simulated PVE by convolving group-level standardized uptake value ratio means in each ROI with 6 mm Gaussian kernels and compared the sexes across ROIs post-smoothing. Widespread sex differences in extracerebral retention were observed. Although attenuating sex differences in cortical tau-PET signal, covarying for extracerebral retention did not impact the largest sex differences in tau-PET signal. Differences in PVE were observed in both female and male directions with no clear sex-specific bias. Our findings suggest that sex differences in FTP are not solely attributed to off-target extracerebral retention or PVE, consistent with the notion that sex differences in medial temporal and neocortical tau are biologically driven. Future work should investigate sex differences in regional cerebral blood flow kinetics and longitudinal tau-PET.
Introduction
Across post-mortem and observational studies, sex differences in levels of tau burden have consistently been found in both clinically normal and clinically impaired older adults. Post-mortem studies of sex differences in neurofibrillary tangle density have suggested a localization of these sex differences in medial temporal regions.1,2 In positron emission tomography (PET), clinically normal (CN) females exhibit greater 18F-Flortaucipir (FTP)-PET signal than males in medial temporal regions, particularly in females with elevated amyloid-beta (Aβ), which is characteristic of early tau deposition in neuropathological staging of Alzheimer's disease (AD). 3 CN females also demonstrate higher FTP-PET signal than males across many regions of the neocortex, regardless of Aβ accumulation, such as the rostral middle frontal, lateral occipital, and parietal regions.4 –6 This is paradoxical to what might be expected in CN based on recent FTP-PET staging models, as these regions are not expected to exhibit elevated tau until a progressed disease stage.7,8 As FTP-PET has applications in clinical trials of Alzheimer’s disease (AD)-modifying therapies, understanding the drivers of sex differences in tau-PET signal is critical for optimizing approaches to primary and secondary outcomes with this modality.9 –12 While an anti-Aβ therapy was recently FDA approved,13 –15 we are at a critical juncture in AD research where anti-tau therapies, as well as combination anti-tau and anti-Aβ therapies, are being explored. 16 At a broader level, elucidating heterogeneities in tau, and how sex may influence tau susceptibility and propagation, is necessary for understanding how tau-focused treatments may prevent or delay AD.
However, it remains unclear whether PET methodological factors, such as extracerebral tracer retention, variation in global morphometry, or signal spillover, contribute to the appearance of female susceptibility to higher levels of tau in neocortical regions. Recent literature on this topic suggests a strong female bias in off-target skull binding, which raises questions about the source of the signal driving sex differences in FTP in neocortical regions.17 –19 What has not yet been examined is the extent to which extracerebral signal may impact (i.e., attenuate or even nullify) sex differences in medial temporal and neocortical FTP signal that have been consistently reported in the literature.
AD-specific sex differences are consistent with the hypothesis that a biological effect may drive higher tau susceptibility in females. For instance, elevated FTP signal in females has been associated with faster rates of cognitive decline in females compared to males. 4 Furthermore, recent longitudinal tau studies have shown faster rates of tau accumulation in females, particularly if they carry APOEε4 or have abnormal levels of Aβ, in both FTP 20 and cerebrospinal fluid 21 studies. These studies provide validity that sex differences result in negative downstream consequences both pathologically and clinically. While the biological mechanisms responsible for female tau vulnerability have yet to be elucidated, it is important to acknowledge that methodological considerations may unduly influence sex differences in tau-PET imaging.
Components that may influence sex differences in FTP signal are radiotracer off-target binding, brain morphology, and spillover. Off-target FTP signal may exist in the skull in amyloid-negative individuals, particularly in females. 17 Females have also been found to exhibit greater tau-PET signal than males in the meninges and skull across FTP, 18F-RO948, and 18F-MK6240 radiotracers, each with variable potency.17,19,22 It is possible that sex-specific extracerebral signal may confound the magnitude of sex differences in cortical FTP signal. Sex differences in brain morphology and structure have also been demonstrated in the literature. Males generally have greater regional volume, gross brain volume, and surface area while females have thicker cortices. 23 Although regional volume or cortical thickness have not been found to affect sex differences in FTP,4,21 the influence of intracranial volume (ICV) is not yet known. Lastly, partial volume effects (PVEs) may pose additional complexities. Partial volume correction (PVC) of PET signal is a common approach to attempting to correct for signal spillover from one region to another. 24 Buckley and colleagues 4 observed that while non-PVC and PVC cortical FTP data both exhibited widespread sex differences, non-PVC sex differences were more potent, indicating a potential influence of PVE.
This work aimed to determine the contribution of confounding neuroimaging factors, such as off-target extracerebral FTP-PET tracer retention, ICV, and PVE, to sex differences in FTP-PET signal. We hypothesized that sex differences in neocortical regions of the brain with the most pronounced sex differences would remain even after adjusting for neuroimaging confounds.
Materials and methods
Participants
Data from 343 clinically normal participants (CN; 199 female, 58%; mean [SD] age = 74 [8.5] years; age range = 50–94 years) and 55 patients with mild cognitive impairment (MCI; 21 female, 38%; mean [SD] age = 77 [7.3] years; age range = 60–92 years) were obtained from the Harvard Aging Brain Study (HABS; 251 CN individuals) and Alzheimer’s Disease Neuroimaging Initiative (ADNI; 92 CN and 55 MCI individuals). The primary goal of this work was to interrogate findings from Buckley et al., 2020; therefore, the same sample was used in this analysis. In each cohort, participants were selected if they were CN or MCI at the time of FTP-PET imaging and had an MRI scan within one year of the PET scan. Sex was categorized based on self-report in both studies. Initial inclusion criteria for both HABS and ADNI have been previously published. 25 Procedures for this study were conducted under the ethical guidelines stipulated by the Partners Human Research Committee, which is the Institutional Review Board for the Massachusetts General Hospital and Brigham and Women’s Hospital. Each cohort was in accordance with the ethical standards of the responsible committee on human experimentation (institutional and national) and with the Helsinki Declaration of 1975, as revised in 2008. Written consent was obtained from all participants in each cohort. Investigators were blinded by anonymous participant identifiers while assembling and analyzing data.
Data availability
Data is available upon request for HABS at https://habs.mgh.harvard.edu/researchers/request-data/ and for ADNI at https://adni.loni.usc.edu/.
MRI
T1-weighted structural images obtained closest to each participant’s 18F-flortaucipir (FTP)-PET scan were processed using FreeSurfer v6.0. 26 (mean date difference between PET and MRI = 70 days) Cortical region of interest (ROI) parcellations were based on the Desikan-Killiany atlas 27 and subcortical ROI parcellations were based on the aseg atlas. 28 Segmentations were quality controlled, including skull stripping, white matter edits, and placement of control points to ensure data accuracy. 25
Tau PET imaging
Both HABS and ADNI use the FTP-PET tracer. FTP-PET acquisition parameters for each study are available online (HABS: https://habs.mgh.harvard.edu/researchers/data-details/; ADNI: https://adni.loni.usc.edu/methods/pet-analysis-method/pet-analysis/). Processing of FTP-PET imaging data for both the HABS and ADNI was completed using in-house processing pipelines as previously described. 4 To characterize regional FTP tracer retention, FreeSurfer defined standardized uptake value ratios (SUVRs) were created using a cerebellar grey reference region without partial volume correction. For any FTP data that were reported with PVC, we used the Geometric Transfer Matrix approach. 29 ROIs were expressed bilaterally.
Extracerebral tracer retention
Broad FTP tracer retention in skull, dura, meninges, and other structures outside of the cortex was quantified. Extracerebral FTP ROIs were created based on FTP signal adjacent to, but outside of, FreeSurfer-defined cortical regions. One extracerebral mask per participant was created using FreeSurfer’s skull and head extracerebral regions. Each mask was visually examined for any distortions and misalignments. Extracerebral FTP signal for a given cortical ROI was defined as mean FTP SUVR within the extracerebral mask at a distance of less than 12 mm from the ROI in question. We chose a radius at which the spill in at a given distance can be safely assumed to be negligible (i.e., the extracerebral signal should not have a notable effect on the cortical signal). Specifically, 12 mm was arrived at by empirically evaluating multiple distances with 12 mm being the optimal distance in both cohorts. As a sensitivity analysis, we investigated sex differences in extracerebral signal at additional radii (Supplemental Figure 1). Models using 28 regions were included in extracerebral analyses; regions that were not proximal to the extracerebral space were not included. Number of voxels in extracerebral ROIs and mean extracerebral to gray matter distances were also recorded. Extracerebral ROIs were defined as SUVRs referenced to cerebellar grey. An illustration of this process is presented in Figure 1.
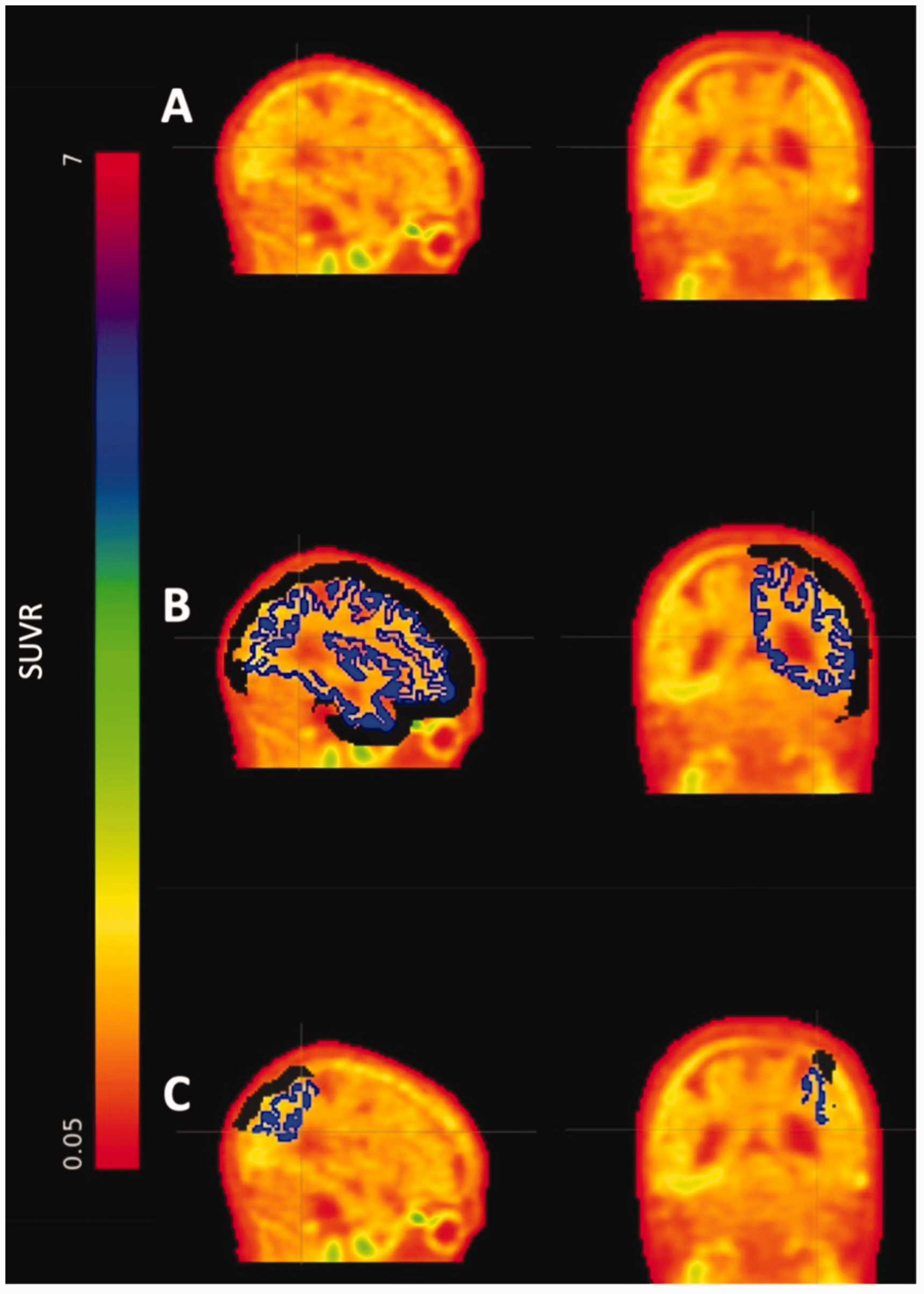
Extracerebral 18F-flortaucipir. (a) 18F-flortaucipir standardized uptake value ratio image. (b) Cortical parcellation in blue and 12mm extracerebral mask in black and (c) Cortical superior parietal parcellation in blue and example of an extracerebral ROI for the superior parietal region in black. ROI: region of interest.
Intracranial volume
The extent to which ICV accounted for sex differences in cortical FTP signal was investigated. As males have significantly higher mean ICV compared to females (t(396) = 4.70, p < .001), we matched ICV to sex using a caliper of 15,000 mm3. The resulting matched sample consisted of 284 individuals (Supplemental Figure 2), including 142 males (mean [SD] age = 76 [8] years; n [%] MCI = 25 [18]) and 142 females (mean [SD] age = 73 [8.6] years; n [%] MCI = 13 [9]).
Partial volume effects simulation
To explore the role of PVEs on sex differences in FTP signal, we simulated PVE values for each participant. 24 Group-level means of each 50 cortical and subcortical FreeSurfer-defined FTP SUVRs were estimated and assigned to each participant’s PET image. For example, the group-level mean of inferior temporal FTP SUVR was 1.34. This value was assigned to all voxels within the inferior temporal region for each participant. A 6 mm Gaussian kernel (standard smoothing kernel within HABS and approximation of the HR+ point spread function 25 ) was convolved across each image to simulate PVE, and regional means were extracted. As the same SUVRs were assigned to each participant before smoothing, these means were used to assess sex differences in PVE.
Statistical analyses
Multiple linear regression models across ROIs estimated the effect of sex on each measure of interest. We first examined sex differences in extracerebral FTP signal (M1). To account for local morphological factors, we additionally covaried for number of voxels or mean distance (M2A and M2B). We then examined if sex differences in cortical FTP signal survived after correcting for extracerebral FTP signal (M3). Vertex-wise brain maps were also created to explore spatial patterns of FTP sex differences after correcting for local extracerebral signal. Next, we investigated interactions of sex and extracerebral FTP signal on cortical FTP signal (M4). For ICV analyses, we used a pooled t-test to test if the matched sample had similar ICV across the sexes. We then estimated sex differences in cortical FTP signal (M5) in the matched sample. Lastly, we estimated sex differences in PVE-simulated values across cortical and subcortical ROIs (M6).
All models corrected for age at the time of FTP-PET imaging and cohort. Model assumptions were checked to validate the appropriateness of these parametric tests. Bonferroni correction was used to adjust for multiple comparisons (two-sided α = 0.0018 for M1, M2A, M2B, M3, M4, M5; two-sided α = 0.001 for M6). Statistics are reported in Supplemental Tables 2, 3, 4 and 5. False discovery rate correction was applied for vertex-wise analyses. Additional sensitivity analyses examined the influence of characteristic off-target FTP regions, specifically the vermis and choroid plexus. Sex differences in these two regions were characterized, and cortical FTP sex differences were estimated after adjusting for signal in the vermis or choroid plexus (Supplemental Table 6).
Results
Extracerebral tracer retention
Sample characteristics by sex are reported in Table 1, while those by cohort and sex are presented in Supplemental Table 1. Results of extracerebral FTP SUVR analyses are presented in Figure 2. We observed widespread sex differences in extracerebral FTP signal, with females exhibiting greater extracerebral signal than males in 21 out of 28 regions (M1). Correcting for number of voxels or mean distance did not attenuate these differences (M2A and M2B).
Sample characteristics by females and males.
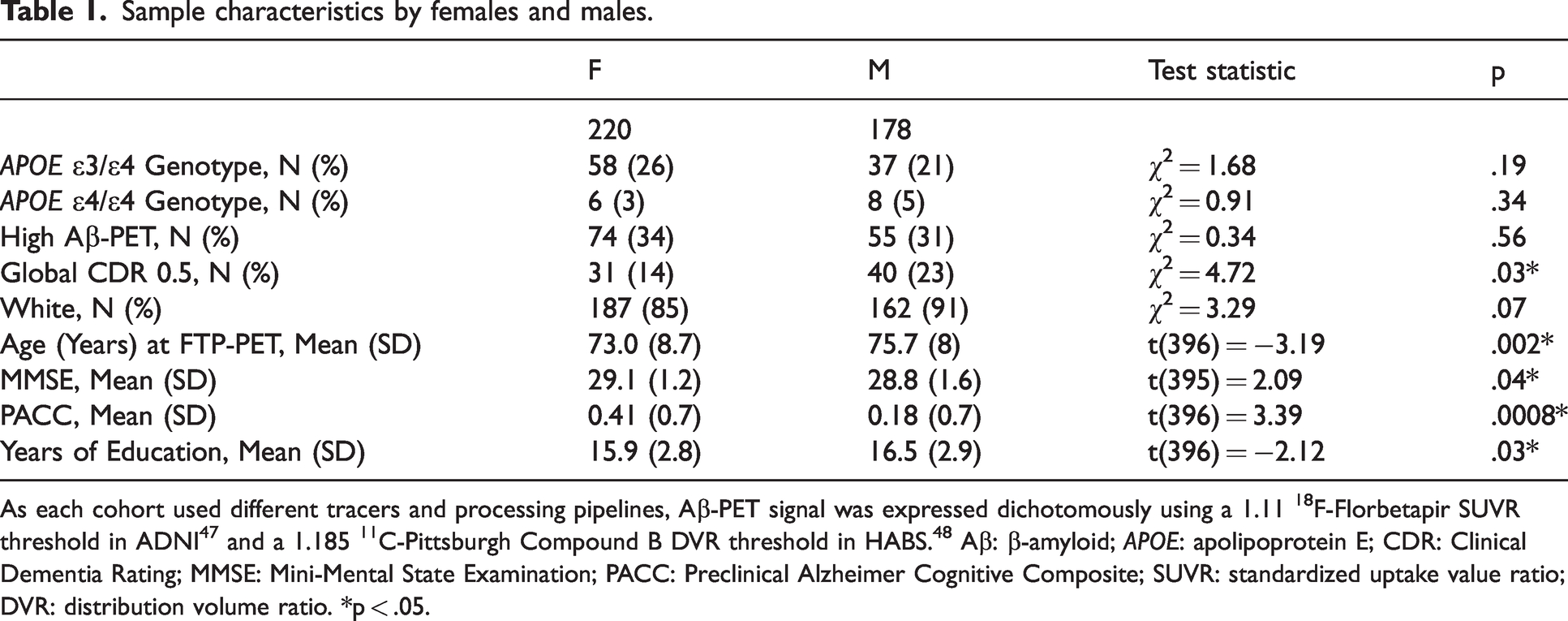
As each cohort used different tracers and processing pipelines, Aβ-PET signal was expressed dichotomously using a 1.11 18F-Florbetapir SUVR threshold in ADNI 47 and a 1.185 11C-Pittsburgh Compound B DVR threshold in HABS. 48 Aβ: β-amyloid; APOE: apolipoprotein E; CDR: Clinical Dementia Rating; MMSE: Mini-Mental State Examination; PACC: Preclinical Alzheimer Cognitive Composite; SUVR: standardized uptake value ratio; DVR: distribution volume ratio. *p < .05.
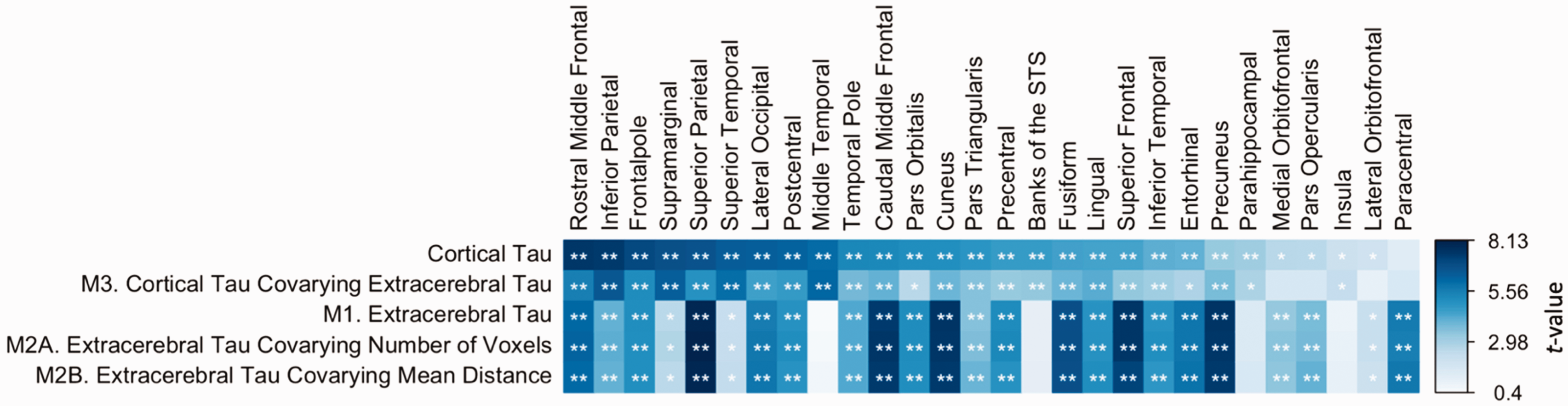
Sex differences in cortical and extracerebral tau. (Row 1) Effects of sex on cortical FTP. (Row 2; M3) Effects of sex on cortical FTP, covarying extracerebral FTP. (Row 3; M1) Effects of sex on extracerebral FTP. (Row 4; M2A) Effects of sex on extracerebral FTP, covarying number of voxels. (Row 5; M2B) Effects of sex on extracerebral FTP, covarying mean gray matter to extracerebral distance. (All) Cells are colored by sex effect t-values, with asterisks indicating significance. Positive t-values infer higher FTP SUVR in females than males. FTP: flortaucipir; STS: superior temporal sulcus; SUVR: standardized uptake value ratio. *.0018 < p < .05, **p < .0018.
When correcting for extracerebral FTP in models estimating sex differences in cortical FTP, the largest sex effects in cortical signal remained (M3). That is, females continued to exhibit significantly greater cortical FTP than males in regions such as the lateral occipital and inferior parietal lobe after adjusting for adjacent extracerebral signal. Vertex-wise sex differences in FTP correcting for local extracerebral signal showed widespread but slightly attenuated differences (Figure 3). We observed effect modification by extracerebral FTP in the association of sex and cortical FTP within the cuneus (p = .036) and paracentral lobule (p = .048; M4), but these findings did not survive multiple comparisons correction. Examining only CN participants did not significantly alter patterns of extracerebral findings.
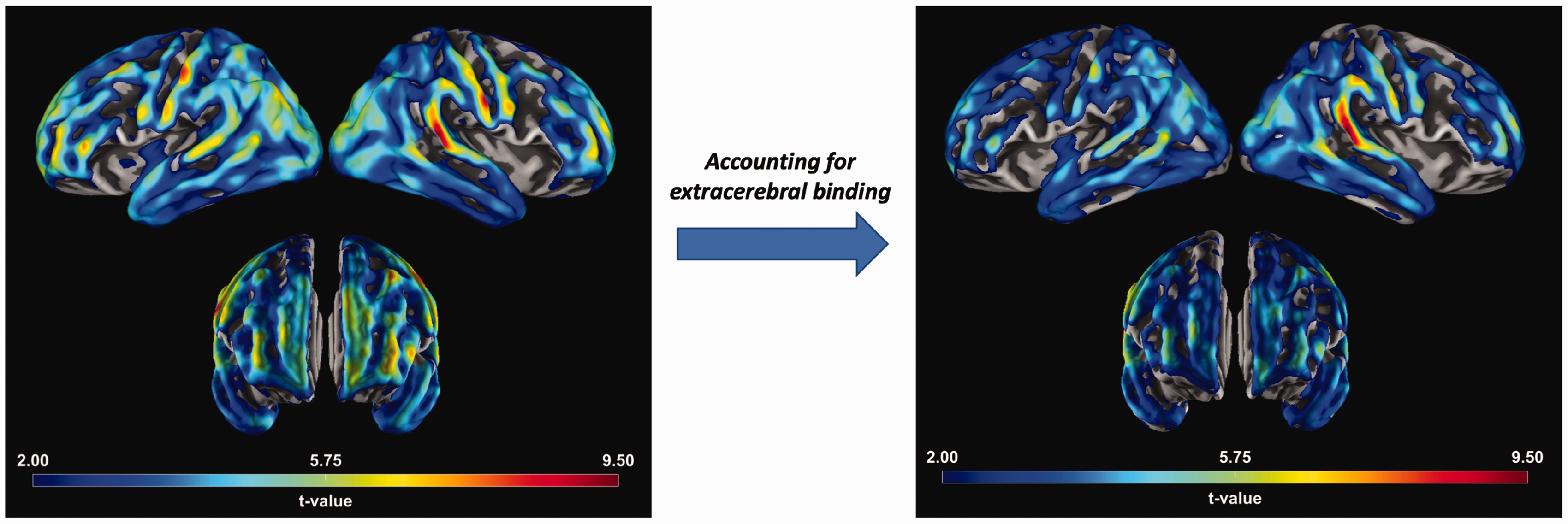
Vertex-wise sex differences. (Left) Sex differences in FTP. (Right) Sex differences in FTP correcting for local extracerebral retention. For example, vertices in the right inferior parietal cortex corrected for right inferior parietal extracerebral FTP retention. (All) Vertex maps of sex differences in FTP. Positive t-values infer higher FTP SUVR in females than males. FTP: flortaucipir; SUVR: standardized uptake value ratio.
Intracranial volume
The effect of sex on cortical FTP signal was estimated in an ICV-matched sample (Figure 4) to reduce confounding by gross morphology. There were no significant differences in ICV between males and females in the matched sample (t(282) = −0.40, p = .69). Sample characteristics in the matched sample resembled those of the full sample, as females had significantly better cognition than males based on the Preclinical Alzheimer Cognitive Composite (t(282) = 2.50, p = .01), Mini-Mental State Examination (t(262) = 2.03, p = .04), and Clinical Dementia Rating (X2(1) = 6.42, p = .01). Additionally, females in the matched sample were significantly younger (t(282) = −3.0, p = .003). Unlike the full sample, no significant difference in years of education was observed (t(282) = −1.60, p = .11). In the matched sample, the largest sex effects in cortical FTP signal remained significant (M5).

Cortical tau sex differences in intracranial volume-matched subsample. (Row 1) Effects of sex on cortical FTP in the full sample. (Row 2; M5) Effects of sex on cortical FTP in the matched subsample. (All) Cells are colored by sex effect t-values, with asterisks indicating significance. Positive t-values infer higher FTP SUVR in females than males. FTP: flortaucipir; ICV: intracranial volume; STS: superior temporal sulcus; SUVR: standardized uptake value ratio. *.0018 < p < .05, **p < .0018.
Partial volume effects
Sex differences in PVE were observed in both male and female directions (Figure 5). After correcting for multiple comparisons, females exhibited greater PVE than males in multiple cortical regions, while males exhibited greater PVE than females in the pallidum, putamen, and thalamus proper (M6).
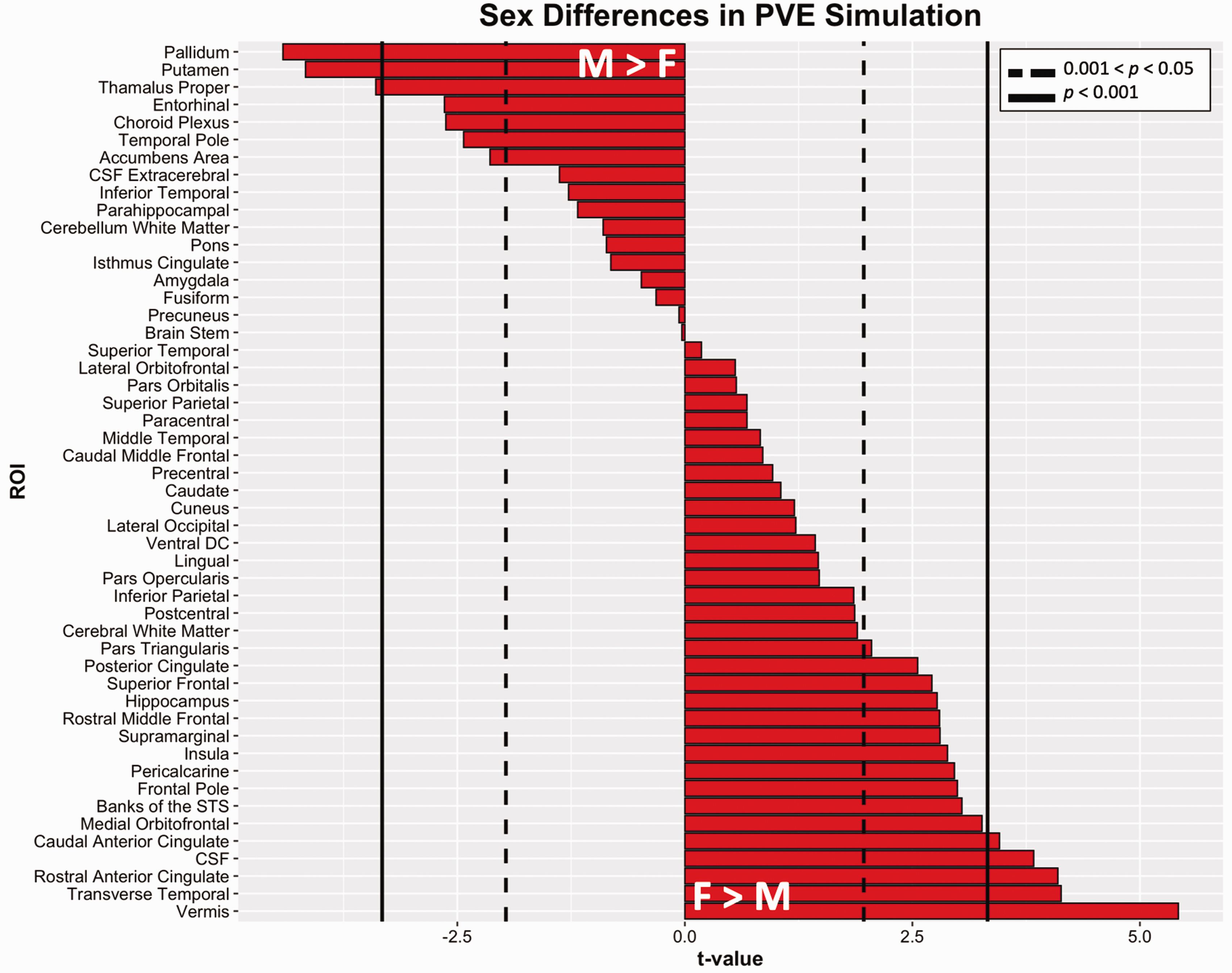
Sex differences in simulated partial volume effects. T-values from regressions of the form PVE∼sex + covariates (M6). Dotted lines indicate thresholds of uncorrected p = .05 and solid lines indicate Bonferroni corrected thresholds of p = 0.001. Positive t-values infer higher 18F-flortaucipir SUVR in females compared to males. CSF: cerebrospinal fluid; DC: diencephalon; PVE: partial volume effect; ROI: region of interest; STS: superior temporal sulcus; SUVR: standardized uptake value ratio.
We plotted standardized betas of PVC and non-PVC sex differences in eight cortical ROIs that we previously reported as having the largest sex effects (Figure 6). 4 Sex differences in PVE varied in size across the eight ROIs, indicating no consistent or pervasive effect of PVE on sex differences in cortical signal. Furthermore, in regions with large female-bias to PVE, such as the rostral middle frontal cortex, there was no inflation of sex differences in corresponding PVC cortical FTP signal compared to non-PVC.
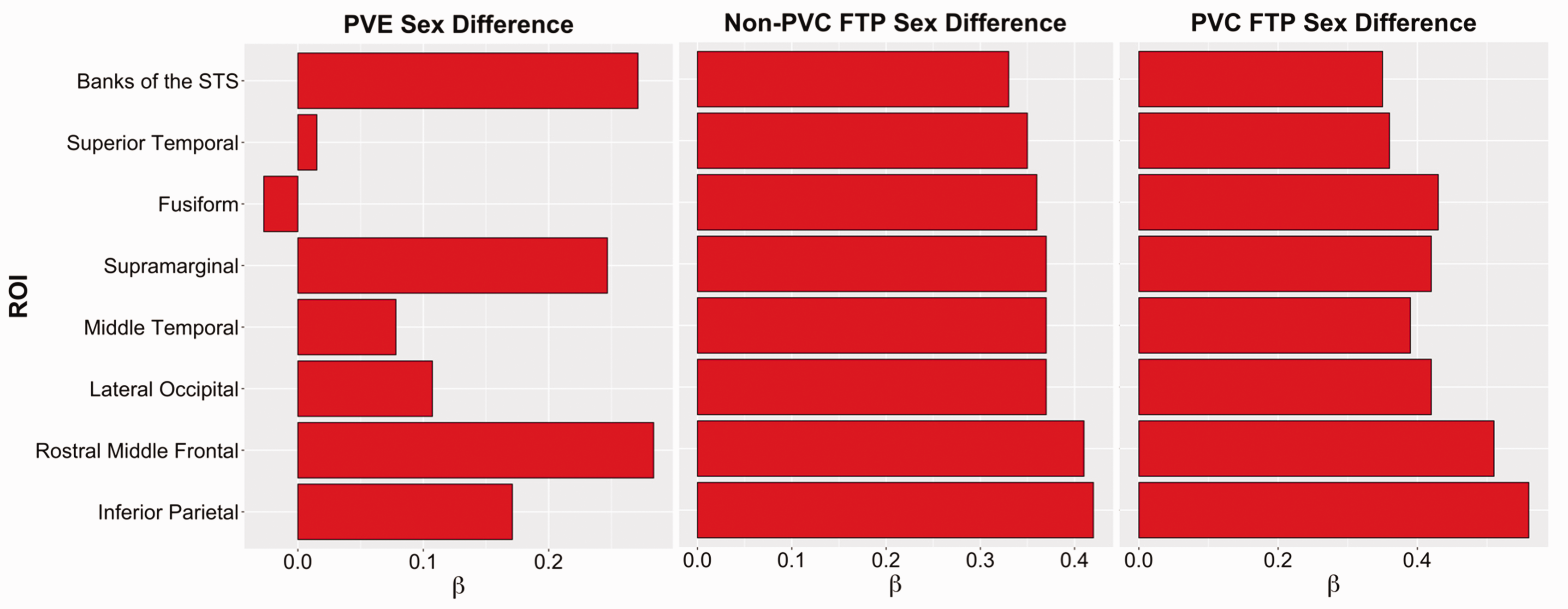
Partial volume effect simulation and cortical tau. Standardized β’s of sex effects in cortical FTP and PVE. Regions showed exhibited the strongest sex differences in cortical FTP. Positive betas indicate higher mean FTP SUVR in females than males. FTP: flortaucipir; PVC: partial volume corrected; PVE: partial volume effect; ROI: region of interest; STS: superior temporal sulcus; SUVR: standardized uptake value ratio.
Discussion
This study examined the influence of neuroimaging variables on sex differences in FTP-PET signal in predominantly CN individuals from two well-characterized cohorts of older adults. We previously reported greater FTP-PET SUVRs in females than males across the neocortex in both cohorts. 4 However, reports of off-target skull and meningeal binding, morphological variation, and PVEs of the FTP radiotracer have not yet been directly investigated as contributors to sex differences in FTP-PET signal.17,19,23,30,31 We therefore explored the independent roles these factors play in sex differences in FTP-PET signal. While sex differences were observed in each of these confounds, they did not explain sex differences in cortical FTP signal. As such, these findings suggest that underlying biological mechanisms remain a possible rationale for sex differences.
Females exhibited higher FTP signal than males across most extracerebral ROIs, consistent with previous findings,17,19,22 even after adjusting for local morphology such as number voxels or mean gray matter-extracerebral distance. Mechanisms underlying sex differences in FTP extracerebral tracer retention remain unclear. One explanation for off-target extracerebral retention is due to distinct FTP binding to leptomeningeal and skin melanocytes, neuromelanin- and melanin-containing cells, respectively. 31 FTP retention in leptomeningeal melanocytes may be most prominent in cells adjacent to the lateral ventricles. 32 Additionally, tracer metabolites retained in the meninges and surrounding skin may complicate cortical tau signal, however this has not been reported in FTP. Furthermore, cortical regions with the highest levels of extracerebral retention in this study did not coincide with such anatomical regions. Additionally, meningeal binding in FTP is less prominent than with tracers such as 18F-MK6240, 17 therefore we argue that female slanted FTP extracerebral retention may be less likely explained by off-target meningeal binding. Another suggestion relates to defluorination of the 18F-fluorine radioisotope. 19 A recent publication examining longitudinal changes in FTP skull binding suggested that binding is stable across time. The same skull binding phenomena was also true of 11C-Pittsburgh Compound B, although to a lesser extent, suggesting that defluorination does not entirely explain off-target binding in skull. The same study suggested that females with higher FTP skull binding exhibited lower bone density on CT scans, suggesting that bone loss may partially drive this off-target tracer retention phenomenon.
While work such as Pereira and colleagues 33 showed that sex differences in FTP exist after the removal of nuisance signal using data-driven methods, our findings extend those of previous studies by examining the direct impact of off-target FTP retention on sex differences in cortical regions. We observed that multiple regions with potent sex differences survived after correcting for extracerebral FTP signal, indicating that extracerebral retention may not confound cortical FTP sex differences. Moreover, only two weak interactions between extracerebral FTP and sex were observed, demonstrating that sex differences in cortical FTP are not solely present at higher or lower levels of extracerebral signal. Extracerebral findings were similarly observed in CN individuals, indicating that sex differences were not simply attributed to progressed clinical stage.
We previously reported that correcting for local cortical thickness did not attenuate sex differences in cortical FTP. 4 However, studies have not yet examined if sex differences are confounded by ICV. To address this question, we utilized ICV matching and observed that sex differences in cortical FTP remained in this matched group. This finding suggests that ICV does not confound sex differences in FTP signal.
Further, we previously observed that non-PVC sex differences were more potent than PVC sex differences, 4 suggesting potential PVE influence. We found sex differences in PVEs in both directions, such that females exhibited significantly greater PVEs in multiple cortical regions, whereas males exhibited greater signal in select subcortical structures. One potential explanation for the male-specific subcortical patterns could be attributed to off-target FTP binding in subcortical structures; 30 however, the male-specific direction remains unclear. As these structures are proximal to ventricles, and males have on average greater ventricular volume than females, 34 this may allow for more spillover effects in females and subsequently more male-directed subcortical signal. Subcortical structures such as the putamen and pallidum are surrounded by vessels and the Virchow-Robin spaces, fluid-filled perivascular spaces that appear throughout the brain parenchyma. Therefore, water versus grey matter tissue may be more of an issue for PVC. 35 These areas are associated with greater cerebrovascular and white matter abnormalities, which can be male biased. However, further investigation into the local morphological features of these areas is needed to fully elucidate these differences. We found female biased PVEs were largest in regions such as the cerebellar vermis, which is a site of great cerebrospinal fluid exposure and is vulnerable to age-related atrophy. Thus, it may be susceptible to greater PVE. 36 The transverse temporal region is locationally similar. As such, it is possible that male-biased PVEs are more vascular-associated, while female-biased PVEs may be more age-associated. The trend of PVE sex differences had varying magnitudes suggesting that the effects were neither robust nor systematic, and that PVEs could not solely explain sex differences in cortical FTP signal.
Although our findings are consistent with the hypothesis that sex differences in tau deposition are biologically driven, these factors have not been fully elucidated. There is growing interest in the interaction between declining estrogen during menopause transition and elevated amyloid levels in females.2,37,38 The use of hormone therapy is also of interest when investigating the impact of sex hormone fluctuation on AD pathology. Recent evidence suggests that CN individuals who have used menopausal hormone therapy show a lower FTP signal compared to non-users and males. 6 Cardiovascular risk factors are another possible contribution, as females are at a greater risk for small vessel disease, 39 which is a well-documented risk factor for AD. 40 Finally, there is a growing story involving the connection between sex-specific immune responses to AD pathology.41,42 Sex dimorphism in microglial activity may result in stronger associations between to amyloid and tau burden across the cortex.
A strength of this study is the large sample size obtained from the combination of both the HABS and ADNI cohorts. One limitation to this study is the small samples of under-represented individuals. Individuals from the HABS and ADNI cohorts tend to have higher education and socioeconomic status, as well as lower racial and ethnic diversity, restricting the generalizability of our findings. Additionally, this was a cross-sectional study, highlighting the need for future examination of longitudinal rates of tau accumulation in the context of off-target retention by sex over time. Recent studies suggest stable off-target skull retention, 19 so it is possible that baseline skull retention could be included as a variable in future longitudinal tau-PET studies. The impact of survival bias on sex differences in FTP-PET remains a potential rationale for the greater susceptibility to tau deposition in females. Some evidence suggests that incidence rates of dementia may be partially explained by the fact that older surviving female participants are different from the male participants due to female longevity or lower female comorbidly. 43 Furthermore, alternative methods for investigating extracerebral and related tracer retention exist. For instance, Fu and colleagues 44 estimated signal perpendicular to the cortical surface to create individualized estimates of skull tau at the perimeter of the cortical ribbon. Smith et al. 17 extracted structures within 5 mm from the outer surface of the CSF layer surrounding the brain to sample signal in the meninges and inner parts of the skull. It is important to acknowledge the specificity and objectives of measures unique to each study. In this study, the metric aimed to quantify broad extracerebral FTP signal rather than specifically targeting the perimeter of the cortical surface. Further, we did not examine the outcome of including extracerebral ROIs in the implementation of GTM PVC. The overall aim of this study was to directly examine proportion the variance explained by adjacent extracerebral FTP signal on the association between sex and cortical tau-PET, which would be unaccounted for shared variance in a GTM model. This is certainly an approach that can be explored in the future. There may also be other variables that we did not consider, such as regional cerebral blood flow kinetics (e.g., R145,46) motion distortion, or variation in time-activity curves. Lastly, we did not consider the implications of other tracers, such as 18F-MK6240 or 18F-RO948.
In summary, the present study investigated the influence of neuroimaging confounds on sex differences in FTP-PET. Substantial sex differences remained even after adjusting for extracerebral FTP retention, ICV, and PVE, supporting the notion of an underlying biological mechanism driving sex differences in tau vulnerability. These findings support the notion that sex differences in tau-PET signal should be a critical factor in clinical trial outcome reporting, particularly in future anti-tau therapy trials. CN females enrolled in prevention trials will likely exhibit higher tau burden than males 4 as well as faster accumulation rates. 20 These differences may impact estimates of intercept and change in trials assessing the reduction and slowing of tau propagation. The biological underpinning of sex differences in tau-PET is critical to understanding drug responses and may influence secondary endpoints of amyloid-specific trials13 –15 and the primary endpoint of tau-specific trials. 11 Hence, it is important to consider sex differences in therapeutic responses, including tau outcomes, and implement sex-specific enrollment criteria to garner a more comprehensive profile of drug efficacy. Further research is needed to elucidate the biological mechanisms underlying sex differences in neocortical tau in both observational and interventional studies of preclinical AD.
Supplemental Material
sj-pdf-1-jcb-10.1177_0271678X231196978 - Supplemental material for Contribution of extracerebral tracer retention and partial volume effects to sex differences in Flortaucipir-PET signal
Supplemental material, sj-pdf-1-jcb-10.1177_0271678X231196978 for Contribution of extracerebral tracer retention and partial volume effects to sex differences in Flortaucipir-PET signal by Matthew R Scott, Natalie C Edwards, Michael J Properzi, Heidi IL Jacobs, Julie C Price, Cristina Lois, Michelle E Farrell, Bernard J Hanseeuw, Emma G Thibault, Dorene M Rentz, Keith A Johnson, Reisa A Sperling, Aaron P Schultz and Rachel F Buckley in Journal of Cerebral Blood Flow & Metabolism
Footnotes
Funding
The Harvard Aging Brain Study is funded by the National Institute on Aging (P01AG036694) with additional support from several philanthropic organizations. Several co-authors’ contributions were also supported by career development awards: R.F.B. is supported by a K99/R00 award from NIA (R00AG061238) and an Alzheimer's Association Research Fellowship (AARF-20-675646). H.I.J. receives funding from NIH-NIA R01AG062559, R01AG068062, and R21AG074220. B.J.H. is supported by the Belgian National Science Foundation (FNRS grant #CCL40010417 and WelBio programme #40010035) and the Queen Elizabeth Medical Foundation. J.C.P. receives funding from NIA R01AG050436 and R01AG052414. Data collection and sharing for this project was funded by the Alzheimer's Disease Neuroimaging Initiative (ADNI) (National Institutes of Health Grant U01 AG024904) and DOD ADNI (Department of Defense award number W81XWH-12-2-0012). ADNI is funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering, and through generous contributions from the following: AbbVie, Alzheimer's Association; Alzheimer's Drug Discovery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen; Bristol-Myers Squibb Company; CereSpir, Inc.; Cogstate; Eisai Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly and Company; EuroImmun; F. Hoffmann-La Roche Ltd and its affiliated company Genentech, Inc.; Fujirebio; GE Healthcare; IXICO Ltd.;Janssen Alzheimer Immunotherapy Research & Development, LLC.; Johnson & Johnson Pharmaceutical Research & Development LLC.; Lumosity; Lundbeck; Merck & Co., Inc.;Meso Scale Diagnostics, LLC.; NeuroRx Research; Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer Inc.; Piramal Imaging; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Private sector contributions are facilitated by the Foundation for the National Institutes of Health (![]() ). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Therapeutic Research Institute at the University of Southern California. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California.
). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Therapeutic Research Institute at the University of Southern California. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California.
Acknowledgements
The authors thank the participants who volunteered their valuable time to these studies.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ contributions
All authors contributed to the study conception and design. MRS, NCE and RFB analyzed and interpreted the data, and drafted and edited the manuscript. MJP, HILJ, JCP, CL, MEF, BJH, EGT, APS, DMR, KAJ, RAS all contributed to the manuscript editing and interpretation of results.
Supplementary material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.