
Abstract
Age-associated cerebrovascular disease impacts brain tissue integrity, but other factors, including normal variation in blood markers of systemic health, may also influence the structural integrity of the brain. This cross-sectional study included 139 individuals between 40 to 86 years old who were physically healthy and cognitively intact. Eleven markers (total-cholesterol, high-density lipoprotein, low-density lipoprotein, triglyceride, insulin, fasting glucose, glycated hemoglobin, creatinine, blood urea nitrogen, albumin, total protein) and five derived indicators (estimated glomerular filtration rate, creatinine clearance rate, insulin-resistance, average glucose, and cholesterol/high-density lipoprotein ratio) were obtained from blood sampling. Diffusion tensor imaging was used to evaluate white matter tissue health. Blood markers were clustered into five factors. The first factor (defined as insulin/high-density lipoprotein factor) was associated with markers of integrity in the deep white matter and projection fiber systems, while the third factor (defined as kidney function factor) was associated with different markers of integrity in the periventricular and watershed white matter regions. Differential segregated associations for insulin and high-density lipoprotein levels and serum markers of kidney function may provide information about distinct mechanisms of brain changes across the lifespan. These results emphasize the need to determine whether therapeutic modulation of systemic health and organ function may prevent decline in brain structural integrity.
Introduction
Major insights in basic and clinical neuroscience have to great degree come through a perspective of the brain as a unique and somewhat independent organ from the more general health of the organism, with the idea that general health may only contribute to neural health in the case of disease. However, it is likely that normal variation in systemic physiology and end-organ function may contribute to overall brain health. Hints that normal inter-individual variation in peripheral and organ health may contribute to neural integrity have recently appeared in the literature demonstrating that variation in blood pressure is associated with brain tissue health in middle-aged and older individuals even in low-risk individuals and younger cohorts.1–3
Aside from a focus on the vascular system, very little is known about how normal variation in somatic physiological processes may impact brain health. Standard blood processing panels include markers of endocrine, cardiovascular, kidney, and hematologic dysfunction and ‘out of range’ values are indicative of disorders such as diabetes, metabolic syndrome, and cardiovascular disease, conditions that are all highly prevalent in older adults. Evidence of an important link between general health markers and cognition in non-disease populations comes from studies on the variation in insulin resistance and its related blood markers,4,5 as well as in glomerular filtration rate (GFR) and kidney filtration substances,6,7 and suggests that metabolic health markers identified in a simple blood test may also be related to brain health.
Our hypothesis was that physiological alterations in blood markers were associated with brain white matter integrity in normal healthy controls as measured with diffusion tensor imaging (DTI). This has been reported previously in patients with type 2 diabetes in whom regional alterations in regional DTI measures were found to be negatively correlated with HBA1c, fasting glucose, and serum creatinine. 8 In patients with end-stage renal disease (ESRD), increased serum urea levels were associated with altered white matter integrity. 9 In both cases, worse cognitive performance was associated with reduced white matter integrity.10,11 Recent work has also demonstrated an association between white matter integrity and insulin resistance, 12 serum cholesterol levels, 13 and kidney function 14 in individuals generally considered disease-free. The current work is much broader in scope examining several putative markers of systemic health. A factor analytic approach allows us to demonstrate the factor structure of the various markers examined and elucidate differential patterns of association among these physiologically and statistically distinct factors. This factor analysis is completely data driven, and there is no prior hypothesis as opposed to in our earlier studies in the same sample on insulin resistance 12 and in a different sample on serum cholesterol levels. 13 The current study demonstrates that several markers in fact correlate with insulin resistance to form a measure of general ‘metabolic’ health in the normal reference range which is strongly related to indices of white matter integrity.
Materials and methods
Study design and participants
A sample of 250 cognitively healthy middle-aged and older adults was recruited through the Massachusetts General Hospital, the local community, and local senior centers. These individuals form part of a longitudinal cohort to evaluate vascular contributions to brain aging. A total of 139 middle-aged and older adults (56 men/83 women) aged 40–86 years was selected for this cross-sectional study based on the availability of fasting venous blood sample and diffusion-weighted imaging MRI data. All datasets used were acquired within a span of three years. All participants were physically healthy, cognitively intact, and literate with at least a high school education. Participants were excluded if they had major neurologic or psychiatric illnesses, history of stroke, significant head trauma, brain surgery or substance abuse, unstable medical illness, cancer within the nervous system or contraindication for MRI scan. Participants with controlled hypertension, dyslipidemia, or type 2 diabetes were not excluded. One hundred twenty-five participants were Caucasian (89.93%), 12 were African American (8.63%), and two were Asian.
Standard protocol approvals, registrations, and patient consents
The study was approved (#2008P001486/MGH) by the Partners Healthcare Internal Review Board and followed the Ethical Principles and Guidelines for the Protection of Human Subjects of Research, generally known as the Belmont Report. All participants provided written informed consent to participate in this research.
Clinical procedures
Assessments included ascertainment of medical history as well as general medical, physical, and neurologic examinations. Overnight fasting venous blood samples were collected on the day of the MRI session for estimation of 11 markers: total-cholesterol, high-density lipoprotein (HDL), low-density lipoprotein (LDL), triglyceride, fasting serum insulin, fasting glucose, glycosylated hemoglobin A1C (HbA1C), creatinine, blood urea nitrogen (BUN), albumin, and total protein. Serum insulin was measured using electro-chemiluminescence immunoassay (Mayo Medical Laboratories, Andover, MA). Five indicators of systemic health – homeostasis model assessment of insulin resistance (HOMA-IR), average glucose level, estimated glomerular filtration rate (eGFR), creatinine clearance (CCL) and cholesterol to HDL ratio (Chol/HDL ratio) – were calculated and will be further referenced in the text as blood markers. The following formulas were used:



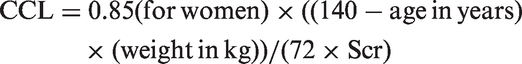
Factor analysis of blood markers
Factor analysis was performed to identify the latent structure of the blood markers. The resulting factors were deemed significant based on eigenvalue criteria (>1.0). Varimax orthogonal rotation was used and variables with absolute factor loadings at 0.40 or above were considered major contributors to a factor. Secondary confirmatory analyses included factor analyses without component rotation and without indicators including age in their calculation.
DTI acquisition and processing
MRI studies were performed on a 3 Tesla scanner (Siemens Trio, Erlangen, Germany). The diffusion-weighted images were obtained using single-shot echo planar imaging with a twice-refocused spin-echo sequence: 16 64 slices, repetition time/echo time = 7980/83 ms, 2 mm isotropic voxels with no slice gap, 60 directions at b = 700 s/mm2 with 10 volumes at b = 0 s/mm2, acquisition matrix = 128 × 128, flip angle = 90°, and total acquisition time = 8 min, 38 s. Data were preprocessed using the FSL Diffusion Toolbox (http://www.fmrib.ox.ac.uk/fsl). Diffusion volumes were eddy current and motion corrected with affine registration to the non-diffusion weighted volumes (b = 0 s/mm2). The diffusion tensor model was fit to the preprocessed DTI data, and the measures of white matter microstructure – fractional anisotropy (FA), mean diffusivity, axial diffusivity and radial diffusivity – were obtained for each individual. FA maps were registered and aligned into a template space using non-linear registration as part of the FSL Tract-based spatial statistics (TBSS) procedure. 17 A mean FA skeleton representing the centers of all tracts common to the group was computed and used to reduce partial-volume effects in the statistical analyses. Other DTI maps were also registered to the mean FA skeleton.
Statistical analyses
Voxel-wise general linear models (GLMs) were performed using FSL to examine regional associations between each of the significant factors and diffusion measures regressing out the following potential confounders: age, sex, the other factors, and motion parameters (translation and rotation 18 ). Due to the long scanning period, we also investigated the effect of scanning date. While there was a significant relationship between scanning date and some blood markers and diffusion measures, these associations were independent of the primary associations found between factor scores and diffusion measures and did not affect any of the study findings when added to the models (not shown). Results were corrected for multiple comparisons voxel-wise and threshold-free cluster enhancement 19 were applied to obtain statistical maps with corrected p-value < 0.05. The results were presented on the MNI152 template and expanded for visualization. The resulting significant clusters from the skeleton-restricted analysis were deprojected into each participant's native diffusion space as regions-of-interest (ROI) using the inverse transform created by TBSS. Average measures of white matter microstructure were extracted from these ROIs in native space and were used for (1) visualization of the data distribution in regions showing significance and (2) to examine whether associations were preserved in secondary factor analyses and secondary statistical models. Additional analyses were performed with ‘surrogate variables’ which had the highest loading of each factor and were considered representative of each factor. General extreme studentized deviate with α = 0.05 was used to detect outliers of independent variables, and results excluding outliers were also presented.
Results
Participants
Demographics, sample averages and reference values of blood markers and indicators.
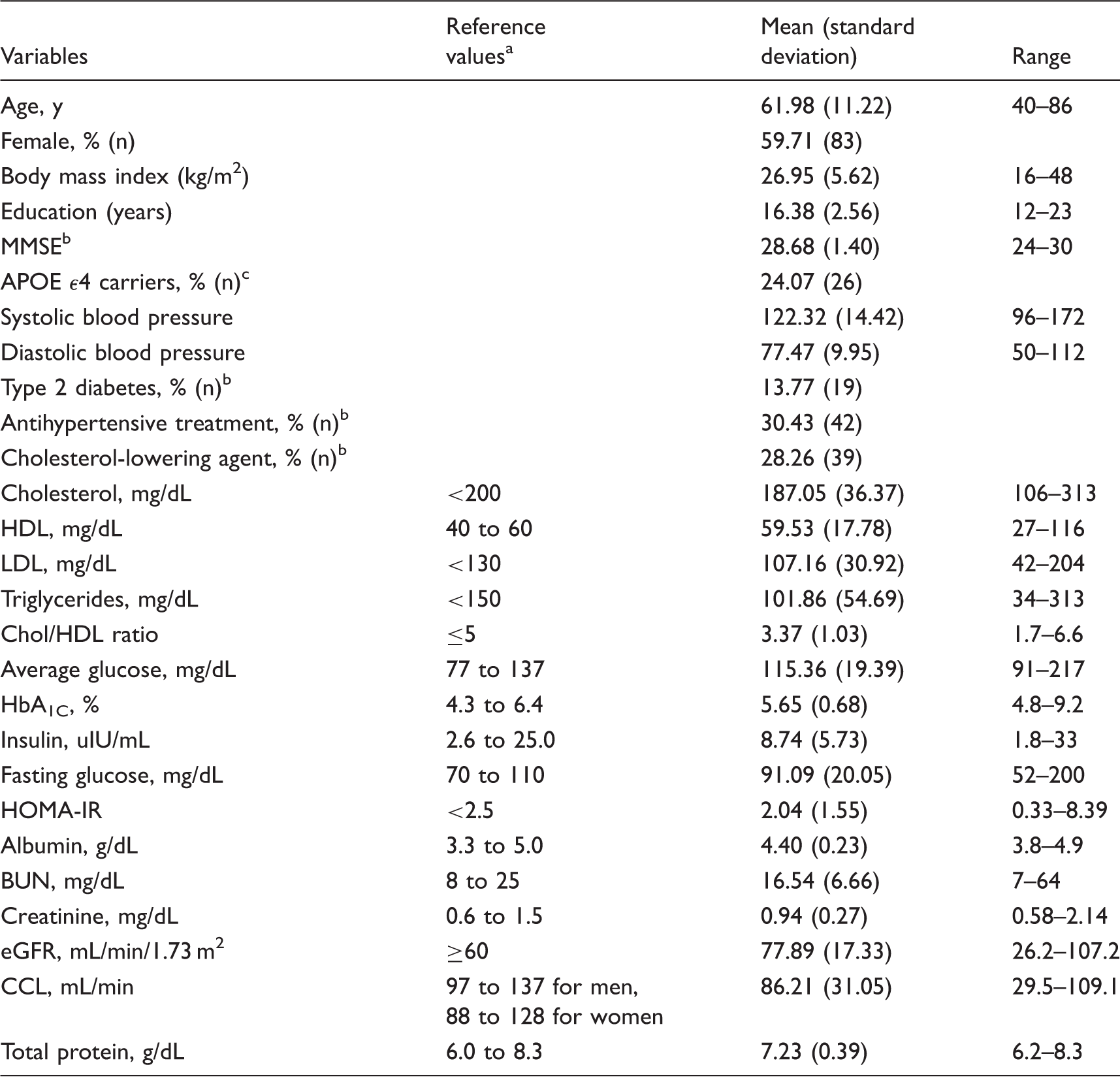
MMSE: mini-mental state examination; HDL: high-density lipoprotein; LDL: low-density lipoprotein; Chol: cholesterol; HbA1C: glycated hemoglobin; HOMA-IR: homeostatic model assessment of insulin resistance; BUN: blood urea nitrogen; eGFR: estimated glomerulus filtration rate; CCL: creatinine clearance.
From Massachusetts General Hospital core laboratory reference ranges (http://mghlabtest.partners.org/MGH-Core-Laboratory-Reference-Ranges.pdf) and the National Cholesterol Education Program 20 ; these values are provided for informational purpose only and may vary based on clinical practice, ethnicity, age, sex, and other factors.
Information missing for one participant.
Information missing for 31 participants.
Factor analysis of blood markers
Primary factor analysis of blood markers and indicators.
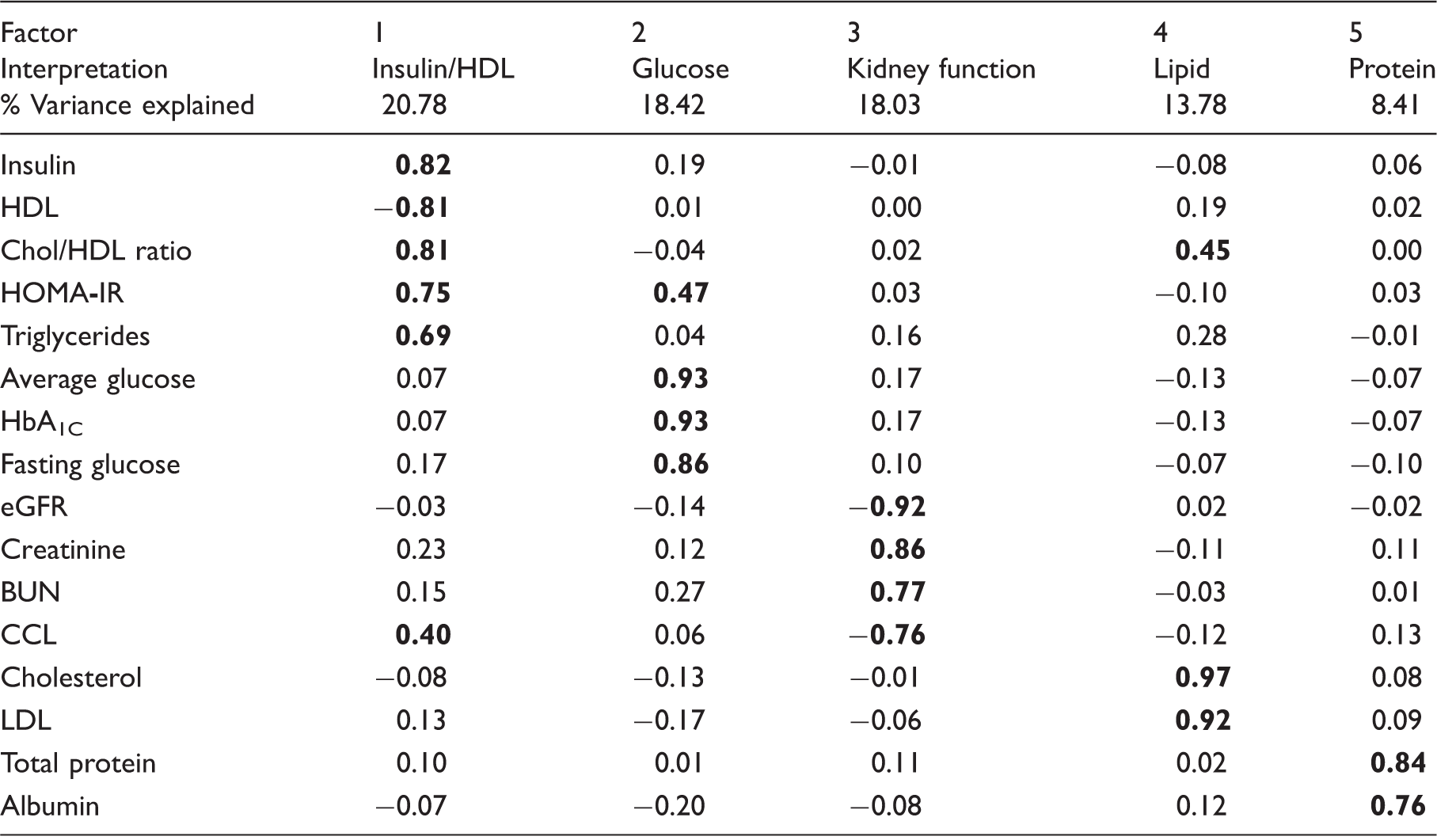
Note: Bolded values indicate an absolute loading value of 0.40 or above.
HDL: high-density lipoprotein; LDL: low-density lipoprotein; Chol: cholesterol; HbA1C: glycated hemoglobin; HOMA-IR: homeostatic model assessment of insulin resistance; BUN: blood urea nitrogen; eGFR: estimated glomerulus filtration rate; CCL: creatinine clearance.
An additional unrotated factor matrix was created to determine the potential influence of the rotation on the overall results. This procedure showed that 10 of 16 variables mainly converged into the first factor (Supplementary Table 1), which was less clearly defined then when using Varimax factor rotation. A more conservative set of factors that did not include variables with an age component (eGFR and CCL) was also created (Supplementary Table 2). This analysis resulted in the same five significant factors; though the variance explained by each factor differed slightly and therefore the factor equivalent to primary factor 3 (KFF) was the fourth factor in this secondary analysis (referred to below as secondary factor 4).
Voxel-wise associations between factors and white matter microstructure
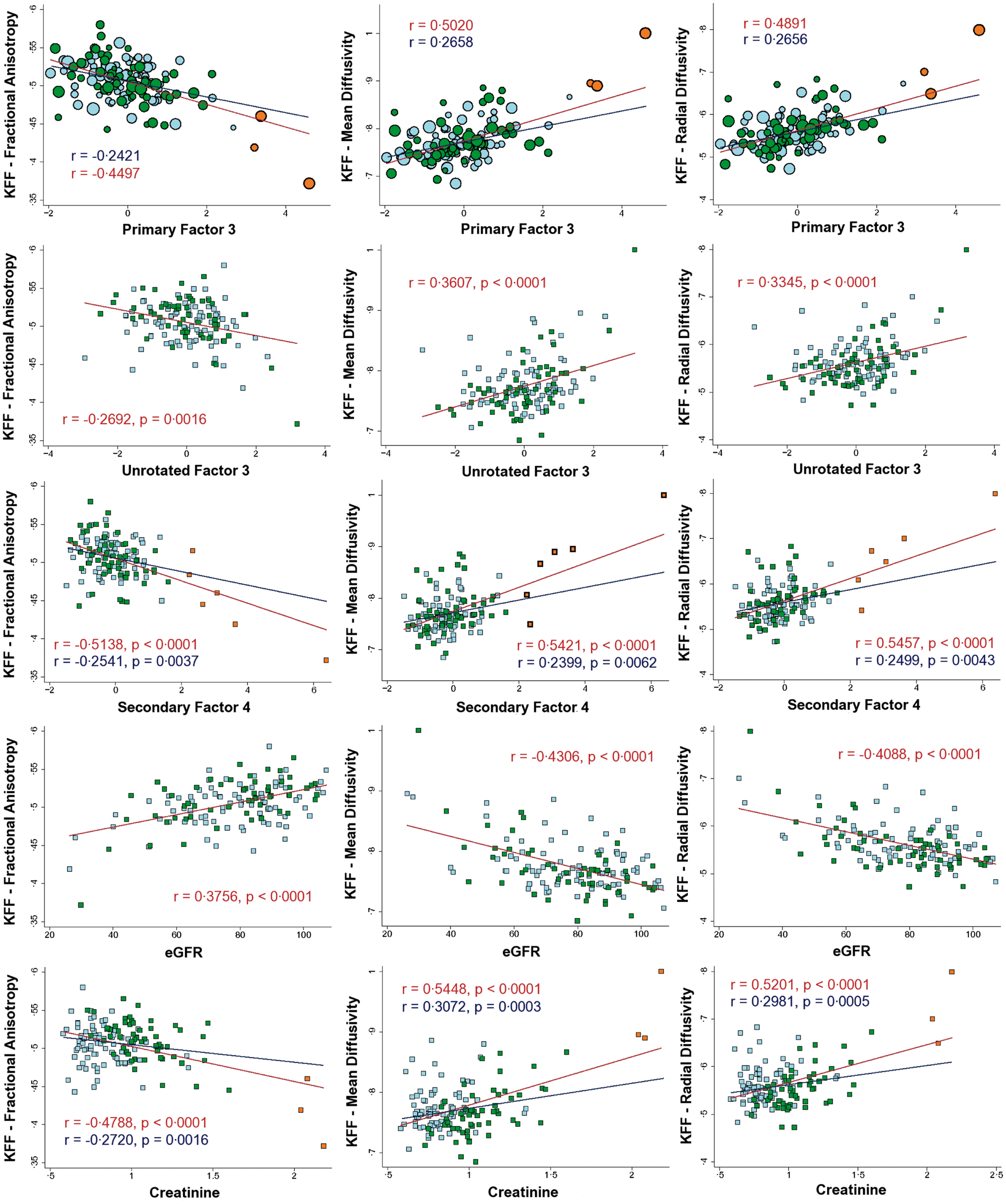
Factor 1 (IHF) and Factor 3 (KFF) showed associations that were almost completely segregated regionally and by diffusion metric. Specifically, IHF showed strongest associations with axial diffusivity that were qualitatively determined to be anatomically distributed within long projection fibers, whereas KFF showed strongest associations in radial diffusivity that seemed to be anatomically preferential to the deep periventricular white matter (see Figure 1 and its caption for more details). The overlap in associations was primarily limited to commissural fibers (mainly FA associations in the corpus callosum). While we did not explicitly correct for multiple comparisons across factors, both IHF and KFF were significantly associated with at least two diffusion metrics at p < 0.01 after voxel-wise correction for multiple comparisons. The results are shown at p < 0.05 to evidence the very low overlap in effects between both factors.
Association between primary factor scores and diffusion measures in all participants.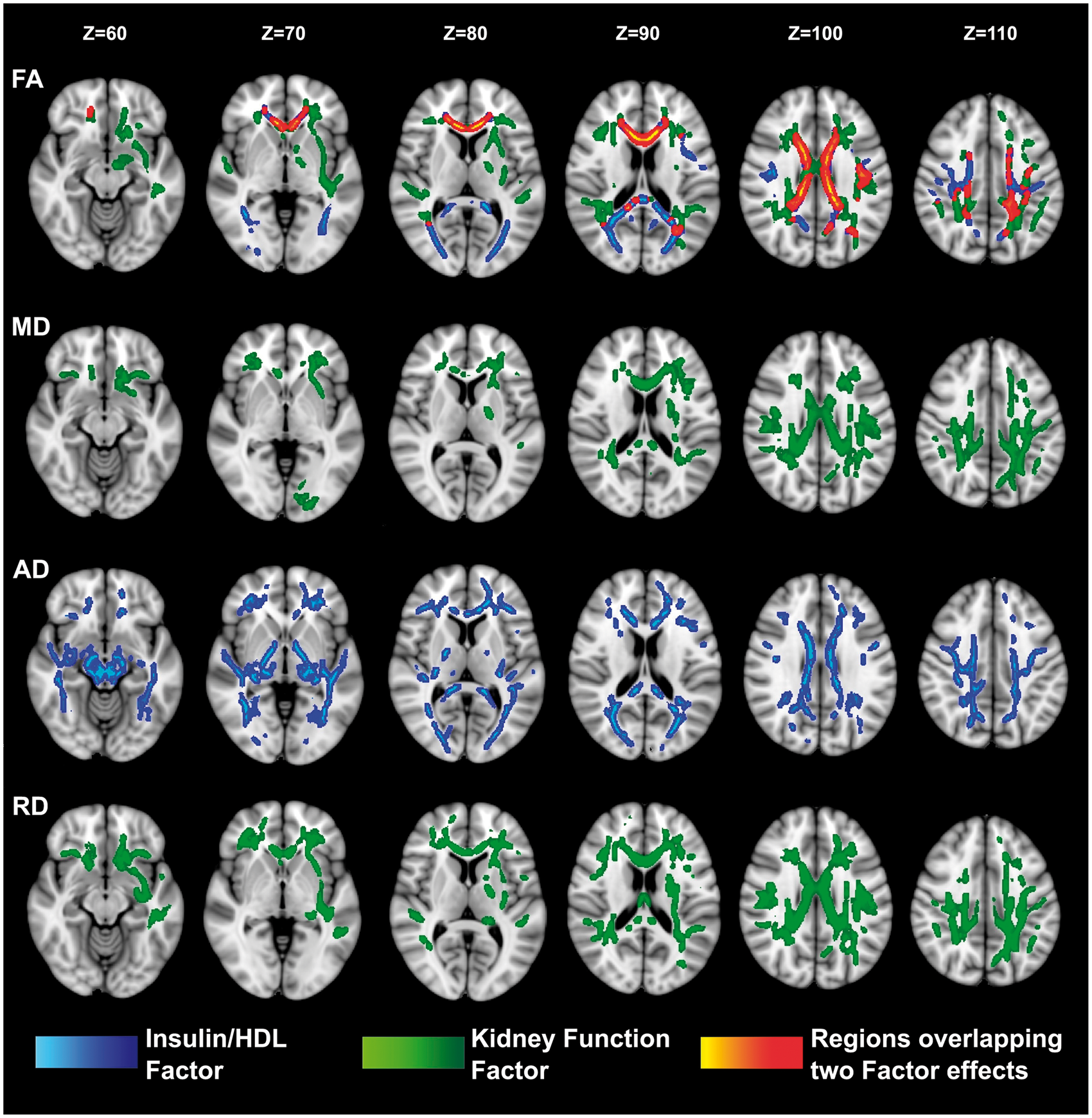
Secondary voxel-wise analyses using ‘surrogate variables’ (the single variable that had greatest loading on IHF and KFF, respectively) also showed the representative dissociation of their associations with measures of white matter microstructure as shown in Supplemental Figure 1.
Secondary/confirmatory analyses using ROI data
ROIs were extracted from the primary voxel-wise analyses to determine the distribution of the data and to confirm that the results were not substantially influenced by age-adjusted indicators or other aspects of the primary factor analysis. Correlations within ROIs remained significant, although reduced in power, for each of the secondary analyses as demonstrated in the scatterplots presented in Figures 2 and 3. Race and education were not significantly related to the factor scores or the extracted ROI data in our sample (Supplementary Figure 2). Additional analyses splitting participants into groups based on whether any of their factor scores diverged too far from the mean similarly showed that each factor was still associated with at least one diffusion measure in the healthiest (Supplementary Figure 3 and Supplementary Table 3).
Scatterplots of the associations between insulin/HDL factor (IHF) and diffusion measures. Scatterplots of the associations between kidney function factor (KFF) and diffusion measures.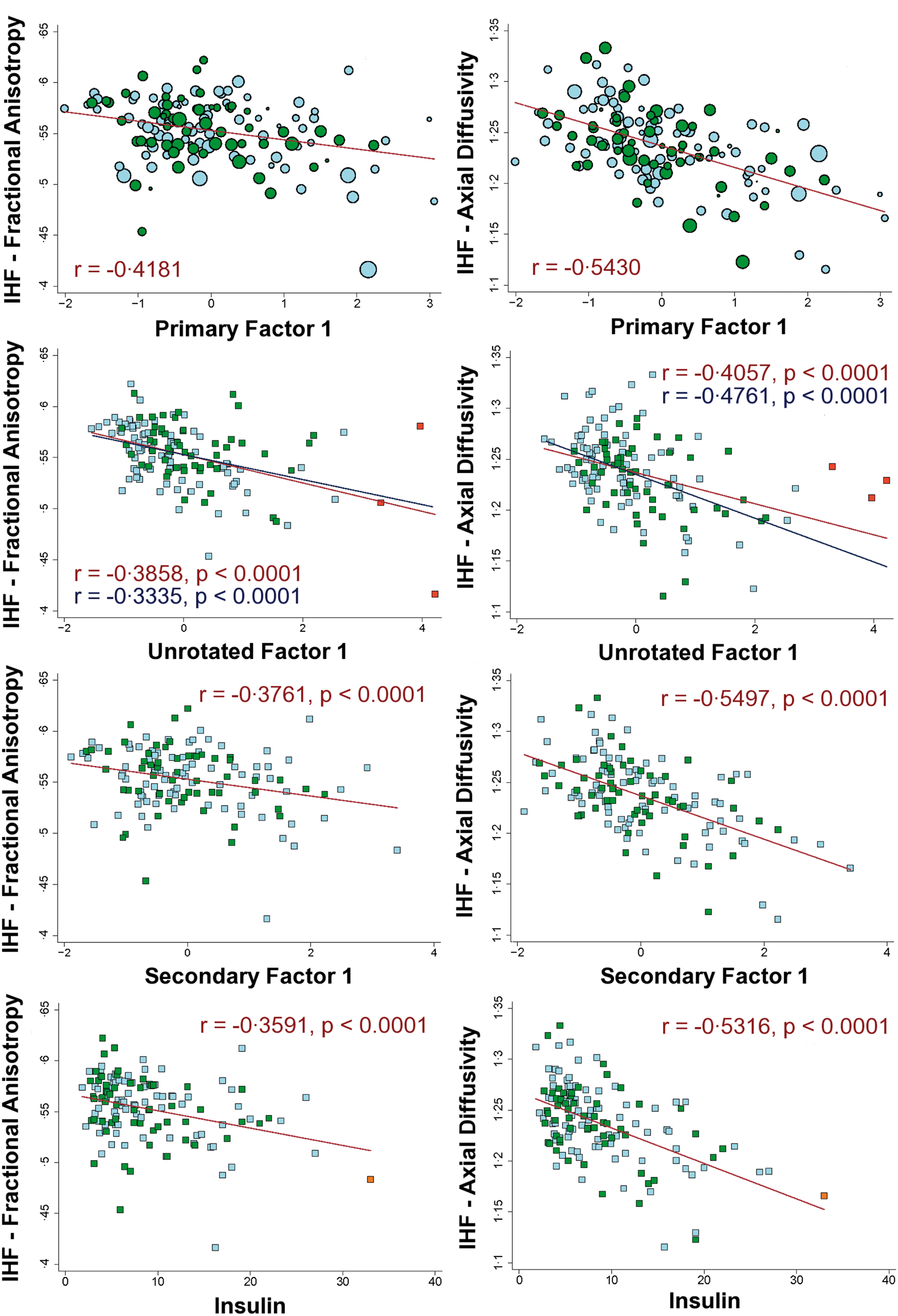
Discussion
The goal of this cross-sectional study was to assess the relationship between regional white matter microstructural integrity and interindividual variation across a diverse collection of standard normal physiological blood markers used clinically to monitor health. Widespread associations with brain tissue microstructure were found for markers of metabolic health and markers of kidney function. Less optimal metabolic health was associated with decreased axial diffusivity in the deep white matter and projection fiber systems, while less optimal kidney function was related to increased mean and radial diffusivity in the periventricular and watershed white matter regions. Associations were statistically independent of age and sex and remained apparent when limiting the sample to only the healthiest individuals, outside of the clinical range of high risk or disease. The differentiation of these effects both on a regional basis and in terms of which diffusion measures are affected suggests distinct mechanisms by which these systemic markers affect the integrity of the brain white matter. These cross-sectional findings may therefore suggest complementary and dissociable influence of subtle variability in systemic physiology on the health of the central nervous system and may further suggest multiple unique routes of therapeutic intervention to explore towards the potential attenuation of neurological aging. Given the potential sensitivity of brain tissue to variations in these health parameters, the findings reported here may have substantial public health implications and may inform neurological practice in reducing risk for age-associated cognitive decline and dementia but may also have implications for general medicine overall. Future interventional studies should focus on optimizing these systemic markers in longitudinal care and determining whether improvement in brain integrity and delay in cognitive decline may be observed.
Previous work by our group2,12,13,21 and others1,22–27 have demonstrated cross-sectional associations between variation in health parameters and white matter microstructure. Our work extends these preliminary findings to evaluate associations with blood physiological parameters generally considered in “normal/healthy” ranges. Unexpectedly, we found not only that indices of metabolic health (IHF) and kidney function (KFF) had strong associations with regional microstructural integrity but also that these effects were distinctly unique. The IHF was more highly associated with regional changes in axial diffusivity in the deep white matter and projection fiber systems. Alterations in the KKF in contrast were more highly associated with regional changes in mean and radial diffusivity in the periventricular and watershed white matter regions, which are regions prone to white matter hyperintensities of vascular origin. The major overlapping effects were limited to findings in the commissural fibers (primarily in the corpus callosum). These fibers, particularly the anterior segments, are known to be particularly vulnerable to the aging process, 28 and these results may therefore elucidate this vulnerability through sensitivity to multiple mechanisms of deterioration. The significance of the differential sensitivity to diffusion measures is unclear. Axial diffusivity, primarily associated with the IHF, has been linked to axonal injury 29 ; however, like our previous study in an overlapping sample on insulin resistance 12 , poorer health was linked to reduced axial diffusivity, contrary to the increase in overall diffusivity observed with typical aging. 28 In contrast, radial diffusivity, primarily associated with the KFF, has been linked to demyelination30,31 and was increased as expected with poorer health. It is, however, clear that multiple processes contribute to variations in diffusion signal, and it is completely unknown from this work whether the associations are driven by variation in any specific histological parameter. Furthermore, it is possible that regions of crossing fibers such as the periventricular regions may be uniquely vulnerable to certain processes, and detection is similarly more sensitive for certain diffusion measures in crossing fibers compared to non-crossing regions. Future work will be necessary to disentangle the regional nature of the associations from the diffusional sensitivity.
This study also adds strong evidence to recent findings of associations between markers of kidney function and brain tissue health in the range of healthy variation. 14 This is in contrast to the effect of significant kidney dysfunction which is known to contribute to neurological complications in ESRD.32,33 By ESRD white matter alterations are widespread in the brain.9–11,34,35 Such changes were suspected to be due to severe disease, however, because renal function slowly decreases with aging as a normal biological phenomenon linked to cellular and organ senescence, 36 the results here suggest that even subclinically reduced renal function may also contribute to decline in white matter microstructure which is known to be a prominent effect of aging. Previous research suggests that retention of solutes that are toxic in high concentration, such as urea, creatinine, parathyroid hormone, and myoinositol may damage the brain in CKD. 37 Alternatively, it is possible that vascular compromise may contribute independently to kidney insufficiency and hypoperfusion and ischemic damage of brain tissue. 38 Greater small-vessel disease burden has been shown to be associated with greater renal impairment.39,40 Furthermore, the fact that the volume of white matter hyperintensity of presumed vascular origin was correlated with serum creatinine in individuals with CKD may support this notion. 41 The finding of dominant distribution of clusters associated with kidney factor in frontal and parietal periventricular white matter, regions also prone to white matter hyperintensities of presumed vascular origin, may also support an independent vascular mechanism to variation in both kidney function and white matter tissue health.
This study also demonstrates that a broader spectrum of overall health contributes to our prior result linking insulin resistance and white matter integrity. 12 Several blood markers were strongly related to insulin resistance and formed the IHF, such as HDL, insulin and triglycerides levels, which in turn were related to the same effects on white matter integrity reported in our previous study solely for insulin resistance. While the hypothesis of our previous study was correct in linking insulin resistance and white matter integrity, the current work demonstrates the importance of a more holistic and data-driven approach revealing that there is a more complex metabolic process that may underlie this association and involve both glucose and fat regulation. Such an approach is needed in future research, for instance to further understand the link between insulin resistance and disorders such as Alzheimer's disease.
There are several limitations to this work that are being addressed in ongoing studies. The results presented here are cross-sectional, and therefore no mechanistic causal inference about directionality can made. The findings do, however, provide suggestive links and follow-up work in longitudinal and interventional cohorts will clarify the role of each of the presented health factors on brain deterioration. While we did not find any effect of race on the results, another limitation is that the sample used in this study consists predominantly of white Caucasian middle-aged and older adults from the Boston metropolitan area and may not allow generalization of the findings to other populations. For instance, we did not find an association between white matter integrity and the blood factor strongly related LDL and total cholesterol, despite finding such an association previously in a completely distinct sample which was disproportionately African American compared to the current sample but also was partially enrolled dependent upon meeting the criteria of having at least one first-degree familial relative with a diagnosis of dementia. 13 The blood markers available for this work may also not reflect the ideal set and future work will require larger marker sets including metabolomic and proteomic data in large participant samples to better track down the primary systemic factors linked to brain health. Additionally, future research will also aim to investigate how potential redundancy between blood markers may affect the factor structure obtained, and will aim to account for other potential confounders such as hydration status not considered here. Finally, although several studies have demonstrated associations between white matter integrity measured by DTI and cognitive performance, we did not examine cognitive associations here as this topic requires an additional primary focus. Despite these limitations, the results present a novel framework for understanding the aging brain in health and disease and provide valuable information regarding follow-up work to understand potential pathophysiologic mechanisms contributing to neural variation with typical aging.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the NIH grant R01NR010827 using resources provided by NIH grants NS042861, NS058793 and by the Center for Functional Neuroimaging Technologies, P41RR14075, a P41 Regional Resource supported by the Biomedical Technology Program of the National Center for Research Resources (NCRR), NIH. This work also involved the use of instrumentation supported by the NCRR Shared Instrumentation Grant Program and/or High-End Instrumentation Grant Program; specifically, grant numbers S10RR021110, S10RR023401, S10RR019307, S10RR019254 and S10RR023043. JPC was supported by the Fonds Québécois de la Recherche – Santé and by the HST IDEA2 Program supported by the Peter C. Farrell (1967) Fund.
Acknowledgments
The authors would like to thank Paul J. Wilkens for his help with the diffusion data.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors' contribution
Chang-Woo Ryu: study concept and design, analysis and interpretation of data, statistical analysis, drafting/revising the manuscript for content; Jean-Philippe Coutu: study concept and design, analysis and interpretation of data, statistical analysis, drafting/revising the manuscript for content; Anna Greka: interpretation of data and revising the manuscript for content; H Diana Rosas: acquisition of data, interpretation of data and revising the manuscript for content; Geon-Ho Jahng: interpretation of data and revising the manuscript for content; Bruce R Rosen: interpretation of data and revising the manuscript for content; David H Salat: study concept and design, acquisition of data, interpretation of data and drafting/revising the manuscript for content.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.