
Abstract
Apoptosis in the endothelium of major cerebral arteries may play a role in the initiation and maintenance of cerebral vasospasm after subarachnoid hemorrhage (SAH). We tested the therapeutic effect of caspase inhibitors on endothelial apoptosis and on cerebral vasospasm in an established dog double-hemorrhage model. Thirty-one mongrel dogs were divided into five groups: control; SAH; SAH treated with vehicle [DMSO]; SAH treated with Ac-DEVD-CHO [a specific caspase-3 inhibitor]; and SAH treated with Z-VAD-FMK [a broad caspase inhibitor]. The inhibitors (100 μM) were injected into the cisterna magna daily from Day 0 through Day 3. Angiography was performed on Day 0 and Day 7. Histology, TUNEL staining, and immunohistochemistry were conducted on basilar arteries collected on Day 7 after SAH. Positive staining of TUNEL, poly(ADP)-ribose polymerase (PARP), caspase-3, and caspase-8 was observed in the endothelial cells of the spastic arteries. Double fluorescence labeling demonstrated co-localization of TUNEL with caspase-3 and TNFα-receptor-1 (TNFR1). Ac-DEVD-CHO and Z-VAD-FMK prevented endothelial apoptosis and reduced angiographic vasospasm. The mechanism of apoptosis in endothelial cells involves TNFR1 and the caspase-8 and caspase-3 pathways. Caspase inhibitors may have potential in the treatment of cerebral vasospasm.
The molecular mechanisms responsible for cerebral vasospasm after spontaneous aneurismal subarachnoid hemorrhage (SAH) have been investigated, and it has been proposed that they may be mediated by oxyhemoglobin (Macdonald and Weir 1994;Weir et al. 1999), pro-inflammatory cytokines (Bavbek et al. 1998;Clatterbuck et al. 2003), protein kinase C (Laher and Zhang 2001;Nakayama et al. 2003), mitogen-activated protein kinases (Zhang 2001), microcirculation compromise (Dietrich and Dacey, Jr. 2000;Ohkuma et al. 2001), and several factors released from the endothelium, including endothelin-1 (Zuccarello 2001;Chow et al. 2002). Due to the complicated pathogenesis of cerebral vasospasm, selective pharmacology intervention targeting the molecular mechanisms of cerebral vasospasm has yet to be developed (Findlay 1997;Mayberg 1998).
Cell death, especially apoptosis after SAH, has been documented in neurons (Matz et al. 2000), smooth muscle (Ogihara et al. 2000), and endothelial cells (Meguro et al. 2001a). Endothelial cell death and detachment of the endothelium promote thrombus formation and trigger cell migration and proliferation (Smith et al. 1985;Clower et al. 1994;McGirt et al. 2002;Borel et al. 2003). Therefore, endothelial apoptosis may trigger, aggravate, and maintain cerebral vasospasm, either in the major arteries (Zubkov et al. 2002b) or in the penetrating arterioles (Zubkov et al. 2000b). Indeed, apoptotic changes in cerebral endothelial cells were identified in a patient who died after SAH (Zubkov et al. 2000a). A family of aspartate-specific cysteine proteases in the cytoplasm (known as caspases) play an important role in the development of apoptosis (Moskowitz and Lo 2003). Among the caspases, caspase-3 has been found to be a key factor in apoptosis (Porter and Janicke 1999;Nicotera 2002). Caspase-3 activation can cleave PARP and irreversibly commit cells to undergo the morphological features of apoptosis, including nuclear condensation and DNA fragmentation (Robertson et al. 2000;Ferrer and Planas 2003). At least two different initiator caspases, caspase-8 and −9, can be responsible for the activation of caspase-3 through distinct cellular signaling pathways (Mattson et al. 2000). Because caspases are major contributors to the apoptotic machinery in many cell types, selecting potent caspase inhibitors has emerged as a new therapeutic strategy (Mattson 2003;Ferrer and Planas 2003).
We hypothesize that inflammatory cytokines (Chyatte 1990;Fassbender et al. 2001;Dumont et al. 2003), especially TNFα, generated after SAH (Gruber et al. 2000;Fassbender et al. 2001;Tran Dinh et al. 2001), bind with TNFα-receptor-1 to activate caspase-8, which in turn activates caspase-3 to cleave poly(ADP)-ribose polymerase (PARP) and produce DNA fragmentation in endothelial cells (Victor and Gottlieb 2002).
MATERIALS AND METHODS
Thirty-one adult mongrel dogs (either sex, weighing 14–20 kg) were used. Dogs were randomly assigned to five groups: normal control [without SAH, n = 3]; SAH [without treatment, n = 6]; SAH treated with vehicle DMSO [n = 6]; SAH treated with Ac-DEVD-CHO [n = 7]; and SAH treated with Z-VAD-FMK [n = 7]. The DMSO and inhibitors were injected into the cisterna magna daily beginning on Day 0 and ending on Day 3. This study was semi-blinded, meaning that the investigators knew the normal and SAH groups since no treatment was applied, but they did not know the content of treatment in the three treated groups (DMSO, Ac-DEVD-CHO, and Z-VAD-FMK). Two dogs were used to test the possible vasoactive effect of the caspase inhibitors. In this additional study, an angiogram was obtained in normal dogs, and then caspase inhibitors were injected into the cisterna magna. The angiogram was repeated at 30 minutes and 2 hours after caspase inhibitor injection.
Experimental dog double-hemorrhage model
Dogs were pre-anesthetized with acepromazine (0.1–0.5 mg/kg), atropine (0.05 mg/kg), and xylazine (1.1 mg/kg), and anesthesia was maintained by isoflurane 1.25%, O2 2L/minute after being mechanically ventilated. The body temperature of the dogs was kept at 37°C with a heating blanket. The mean arterial blood pressure and blood gas levels were monitored through a catheter inserted into the femoral artery and were maintained within normal ranges. The flow of isoflurane was adjusted during surgery according to the values of the mean arterial blood pressure and blood gas to control the depth of anesthesia. Subarachnoid hemorrhage was induced according to the method of Varsos et al (Varsos et al. 1983) as described in our previous publications (Kimura et al. 2002a;Satoh et al. 2002;Kimura et al. 2002b;Kusaka et al. 2003). For cerebral angiography, the left vertebral artery was catheterized with a No. 4 French catheter via the right femoral artery. A baseline vertebrobasilar artery angiogram was obtained. The cisterna magna was punctured transcutaneously, and 0.4 mL/kg of CSF was withdrawn. An equal amount of arterial blood was withdrawn from the femoral artery and immediately injected into the cisterna magna within 1 minute to avoid possible blood clotting in the syringe. The dogs were then tilted at a 20° angle for 20 minutes with their heads down, in a prone position, to permit pooling of blood around the basilar artery.
The first blood injection was considered to be Day 0. On Day 2, the same blood injection procedure was repeated under identical anesthesia conditions and within the same mean arterial blood pressure and blood gas levels without performing angiography. The angiogram was repeated on Day 7 before all dogs were sacrificed.
Caspase inhibitors
Two caspase inhibitors were purchased from BIOMOL. Ac-DEVD-CHO is a highly specific caspase-3 inhibitor (BIOMOL). Z-VAD-FMK is a reversible, cell-permeable, broad-spectrum caspase inhibitor (BIOMOL). The dosages of the inhibitors were individually calculated for each dog to reach similar drug levels in the dogs’ cerebrospinal fluid (CSF), taking into account the relative size of the CSF space to obtain the final concentration in the CSF, assuming that canine CSF volume is 2 mL/kg (Zoghbi et al. 1985). Both caspase inhibitors were diluted in dimethyl sulfoxide (DMSO). One ml of CSF was withdrawn and mixed with the caspase inhibitors (about 200 μl). The inhibitor containing CSF was intracisternally injected within 30 seconds to obtain a final concentration of caspase inhibitors in the CSF of 100 μM. The first injection was conducted at 30 minutes after the first blood injection and was repeated daily for an additional three days.
Measurements of arterial diameter
Arterial diameters were measured in a double-blind fashion on magnified angiograms (Tibbs et al. 2000;Satoh et al. 2002;Kimura et al. 2002b). To eliminate magnification differences on the angiograms, a radio detectable scale was placed on the dog's chin during the angiography run. The same scale was used consistently and was put at the same point on each dog's chin. Relative to the size of this scale as a standard, all arterial diametric values were adjusted. Two researchers (Drs. Zhou and Yamaguchi) independently measured the arterial diameters on the magnified angiograms at 3 points: the distal (just before the bifurcating superior cerebral arteries), the proximal (just after basilar union), and the central (the midpoint between the previous two points) portions of the basilar artery. The mean of these 3 measurements was calculated to yield the arterial diameter. The mean of the values measured by the two researchers was taken as the final diameter of the basilar artery. The caliber of the basilar artery on Day 7 was calculated as a percentage of the mean basilar artery diameter on Day 0 before blood injection in each dog.
Clinical assessment
Clinical scores were recorded based on the independent observations of a veterinarian who was blinded to the study and one of the researchers (Dr. Yamaguchi). The scores were recorded daily. Three clinical observations were recorded and scores were given (Tibbs et al. 2000;Kusaka et al. 2003):
Appetite: normal appetite = 1; decreased appetite = 2.
Activity: dogs active (barking, wagging tails) = 1; dogs not active = 2
Neurological deficit: no neurological deficits = 1; apparent neurological deficit (paresis, ataxia) = 2.
Morphological assessment
After being euthanized with an overdose of pentobarbital (150 mg/kg), the dogs were perfused via both common carotid arteries with 200 mL of 0.1M PBS and then 500 mL of 4% paraformaldehyde in 0.1M PBS (pH7.4). The brain was removed and post-fixed with the same fixative solution (Satoh et al. 2002). The basilar artery was sectioned by a cryostat (Leica CM3050 S) into sections 4 μm thick. A series of sections were obtained from dogs in the normal control, SAH, SAH+DMSO, SAH+Ac-DEVD-CHO, and SAH+Z-VAD-FMK groups. Sections from each dog were divided into several groups for H&E staining, immunohistochemistry, TUNEL staining, and double fluorescence staining, respectively. They were observed under an OLYMPUS BX51 microscope.
H&E staining.
Sections were stained in Hematoxylin for 2 minutes and Eosin for 1 minute. They were then dehydrated and mounted with permon as described previously (Yin et al. 2003). The average wall thickness and the cross section area of the basilar artery were measured with Imaging-Pro-Plus software using an Olympus microscope as described previously (Gules et al. 2002;Perkins et al. 2002). Each basilar artery was measured at 4 points (3, 6, 9, and 12 clocks) to sum the average wall thickness (Table 1).
Histological quantification of the basilar artery
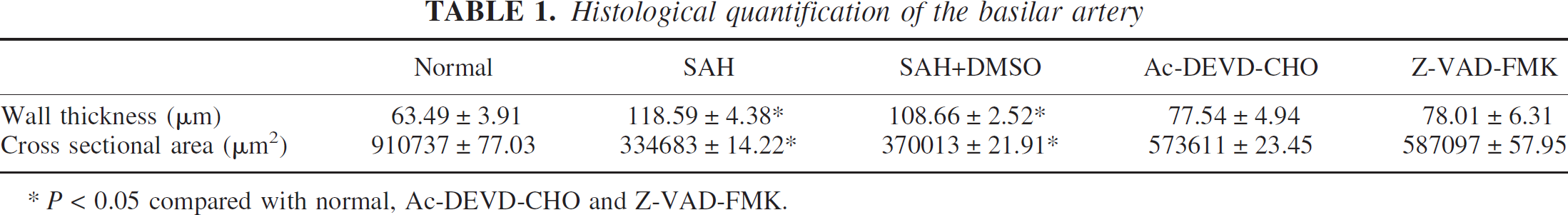
P < 0.05 compared with normal, Ac-DEVD-CHO and Z-VAD-FMK.
TUNEL staining.
Sections were stained by TUNEL, and the TUNEL positive cells were expressed by fluorescein-dUTP with dNTP and POD with 3–3′ diaminobenzidine (DAB) (manufacturer's protocol for the in situ Apoptosis Detection Kit [Roche Inc.]) as described previously (Yin et al. 2003).
Immunohistochemistry staining.
The methods for immunohistochemistry staining have been described previously (Yin et al. 2003). Four series of sections were incubated in 3% hydrogen peroxide (H2O2) and diluted in PBS (10 minutes) to prevent reaction with endogenous peroxidases. After 30 minutes in 3% normal serum in PBS, the sections were incubated with primary antibodies overnight at 4°C. The antibodies used were purchased from Santa Cruz Inc. and include: rabbit anti-PARP [1:200], rabbit anti-caspase-8 [1:200], rabbit anti-casepase-3 [1:200], and goat anti-TNFR1 [1:200], respectively. After rinsing them with PBS, the sections were treated with a rabbit (or goat) ABC kit (Santa Cruz Inc.). The sections were then incubated with goat anti-rabbit (or donkey anti-goat) IgG as a secondary antibody (1:200) for 30 minutes and were placed in avidin-peroxidase complex solution containing avidin-peroxidase conjugate for 30 minutes. Peroxidase activity was revealed by dipping the sections for 5 minutes in a mixture containing DAB and H2O2 (ABC kit, Santa Cruz Inc.) at room temperature. The sections were mounted, air-dried, dehydrated, and cover-slipped. Application of control serum, instead of the primary antibody, on another section of the same brain provided a negative control for each staining.
Double fluorescence staining.
The methods for double fluorescence labeling have been described previously (Yin et al. 2003). Sections from SAH dogs were used for double labeling with TUNEL (green, fluorescein dUTP and dNTP Kit, Roche Inc) and rabbit Von Willibrand (a specific endothelium antibody, Chemicon Inc.); TUNEL and rabbit anti-caspase-3; and TUNEL and rabbit anti-TNFR1 (red, goat anti-rabbit IgG-TR [Texas Red]1:200, Santa Cruz Inc.). Some sections were double labeled with goat TNFR1 (red, donkey anti-goat IgG-TR 1:200) and rabbit caspase-3 (green, goat anti-rabbit IgG-FITC [fluorescein isothiacyanate] 1:200, Santa Cruz Inc.) fluorescence staining, respectively. Sections were cover-slipped with 30% glycerin and observed under an OLYMPUS BX51 microscope with fluoresce light. The wavelength of Texas Red is 630.22 nm. The Texas Red was excited by a Dye Laser at 595–605 nm and was emitted at 620 nm. The wavelength of FITC (fluorescein isothiacyanate) is 530.30 nm. The FITC was excited by an argon laser at 488 nm and was emitted at 530 nm.
Data analysis
Data are expressed as mean ± SEM. Statistical differences between the control and other groups were compared by one-way ANOVA and then the Tukey-Kramer multiple comparison procedure if a significant difference had been determined by ANOVA. The clinical behavior scores were compared by Kruskal-Wallis One Way ANOVA on Ranks, and then, if significant differences were found, the t-Test procedure was used. A probability value of P < 0.05 was considered statistically significant.
RESULTS
Angiography
Severe angiographic vasospasm was observed in the basilar arteries in the SAH and SAH+DMSO groups (Fig. 1A). Mild to moderate vasospasm was observed in the groups of dogs treated with Ac-DEVD-CHO and Z-VAD-FMK (Fig. 1A). The mean residual diameters of the basilar arteries on Day 7, as a percentage of that on Day 0 before blood injection, were: SAH − 54.83 ± 19.95%, SAH+DMSO − 49.25 ± 19.84%, Ac-DEVD-CHO − 78.73 ± 12.92%, and Z-VAD-FMK − 83.94 ± 10.34% (Fig. 1B). The diameters of the basilar arteries in the Ac-DEVD-CHO treated group were significantly greater than those of the SAH+DMSO group (P < 0.05, ANOVA). The diameters in the Z-VAD-FMK treated group were significantly greater than those of the SAH and SAH+DMSO groups (P < 0.05, ANOVA). No statistical difference was noted between the two treatment groups (P > 0.05, ANOVA, Fig. 1B).
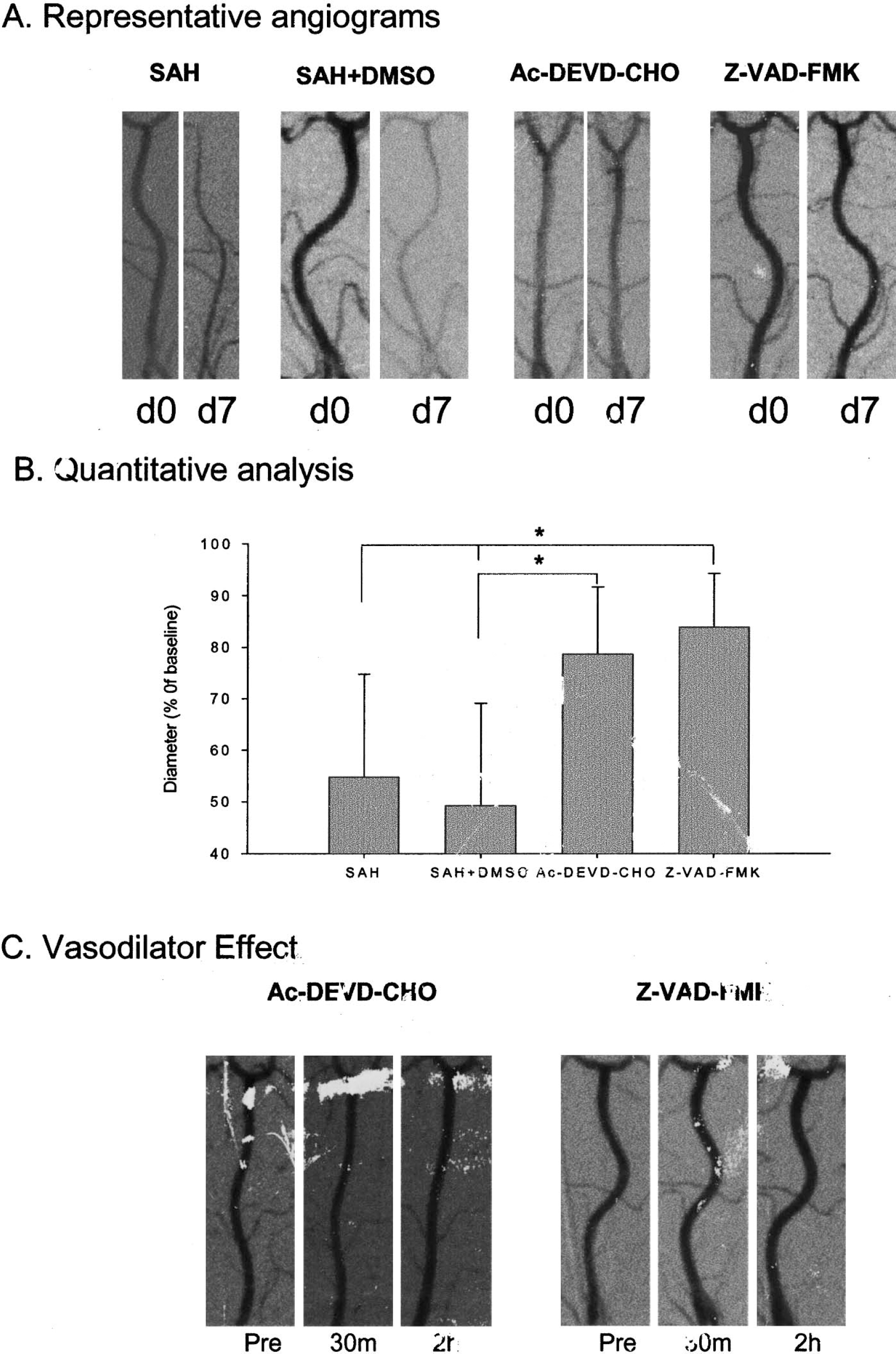
Representative angiograms and summary of diameters.
The possible direct vasoactive effects of the caspase inhibitors were evaluated by angiography. After a baseline angiogram was obtained, the caspase inhibitors were injected into the cisterna magna, and angiography was repeated at 30 minutes and 2 hours. As shown in Fig. 1C, the average diameter of the basilar artery in the Ac-DEVD-CHO dog was 1.02 mm (baseline), 1.00 mm (30 minutes after injection), and 1.08 mm (2 hours after injection). In the Z-VAD-FMK treated dog, the average diameter of the basilar artery was 1.24 mm (baseline), 1.30 mm (30 minutes after injection), and 1.28 mm (2 hours after injection). We did not observe any direct vasoconstriction or vasodilation effect of these caspase inhibitors.
Clinical behavior scales
The clinical behavior scales on appetite, activity, and neurological deficit of the 4 groups of dogs are shown in Fig. 2. The appetite of the dogs in both treatment groups was significantly better than the SAH or SAH+DMSO groups during the first three days (P < 0.05, Fig. 2A). After Day 4, the appetite of the dogs in the SAH and SAH-DMSO groups began to improve. However, the appetite of some dogs in the Ac-DEVD-CHO treatment group began to “fluctuate” (lose appetite).
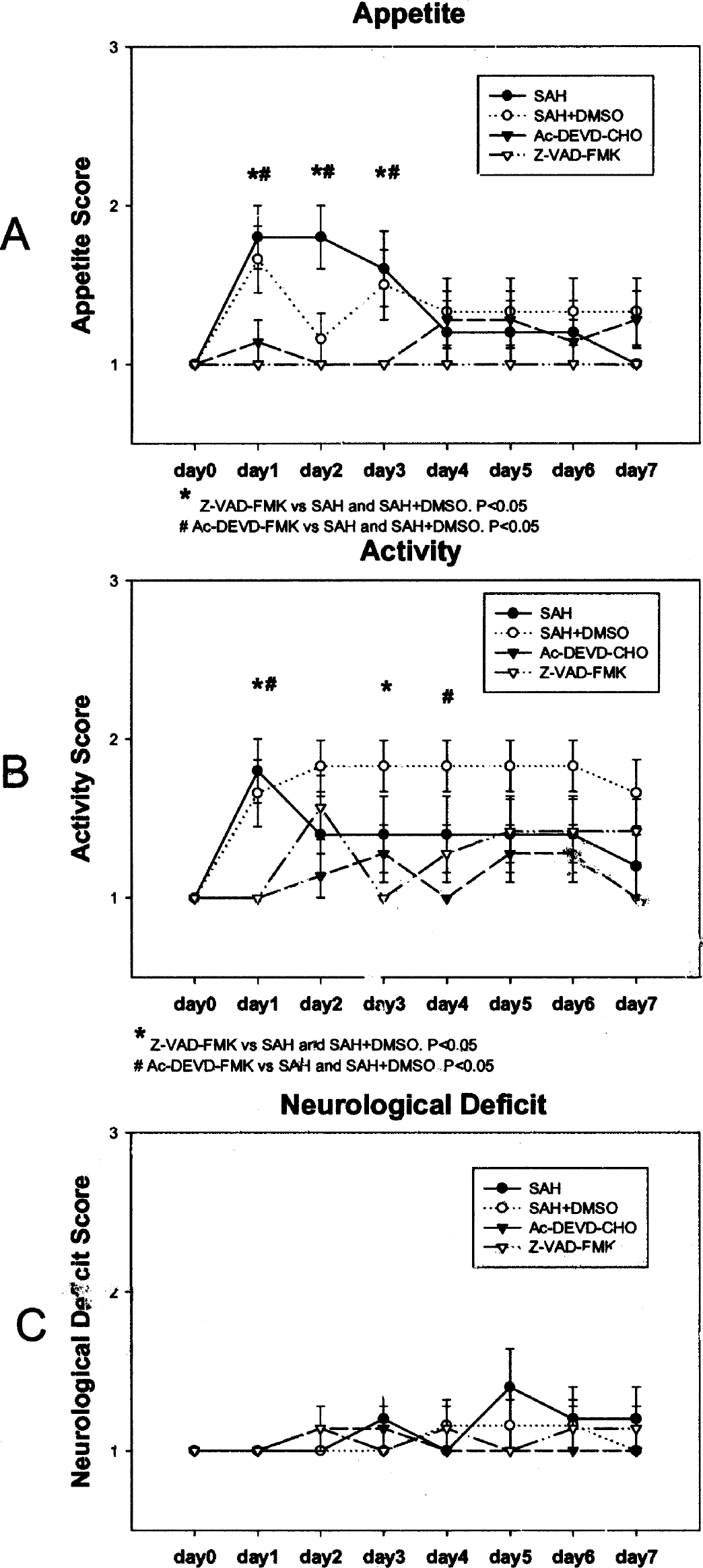
Clinical behavior scales on appetite, activity, and neurological deficit.
The activity of the dogs in both treatment groups was significantly better than those in the SAH and SAH+DMSO groups at Day 1. The activity of the Z-VAD-FMK treated dogs was better than the SAH+DMSO group at Day 3, and the activity of the Ac-DEVD-CHO treated dogs was better than the SAH+DMSO group at Day 4 (P < 0.05, Fig. 2B). Higher (less) activity scores were observed in the SAH+DMSO group than the SAH group two days after SAH (even though P > 0.05, ANOVA), probably due to the effect of multiple injections of DMSO into the cisterna magna. Similarly, the activity score began to “fluctuate” after Day 1 in both of the inhibitor treated groups.
On the contrary, most dogs did not have neurological deficits, and no significant difference was observed among the groups (P > 0.05, Fig. 2C). Mild to moderate neurological difficulties were observed in several dogs in all groups two to three days after SAH, which may be due to the effect of vasospasm in the SAH and SAH+DMSO groups, or due to the possible “toxic” effects in the caspase inhibitor treated groups.
Morphological assessment
Morphological assessment was conducted on samples from the basilar arteries on the pons with H&E, TUNEL, and immunohistochemistry staining.
H&E staining.
No blood clots were observed in the normal control dogs (Fig. 3A1). Subarachnoid blood clots (black “star” signs) were found on the surface of the pons and medulla in all four SAH groups (Fig. 3B1, C1, D1, E1). Severe arterial contraction was observed in the SAH and SAH+DMSO groups, characterized by a decreased vessel lumen, a thickened vessel wall (Fig. 3B2, C2), a corrugated internal elastic lamina (IEL) (Fig. 3B3, C3), contracted smooth muscle cells, and a shrunken endothelium, which desquamated from the basal lamina (Fig. 3B3, C3). The wall thickness and the cross section area of the basilar artery were measured and are summarized in Table 1. In the samples from dogs treated with Ac-DEVD-CHO (Fig. 3D2) and Z-VAD-FMK (Fig. 3E2), the lumen and the thickness of the vessel wall looked similar to those in the normal basilar artery (Fig. 3A2, Table 1), with smooth IEL, normal shaped endothelial cells, and elongated smooth muscle cells (Fig. 3A3, 3D3, 3E3).
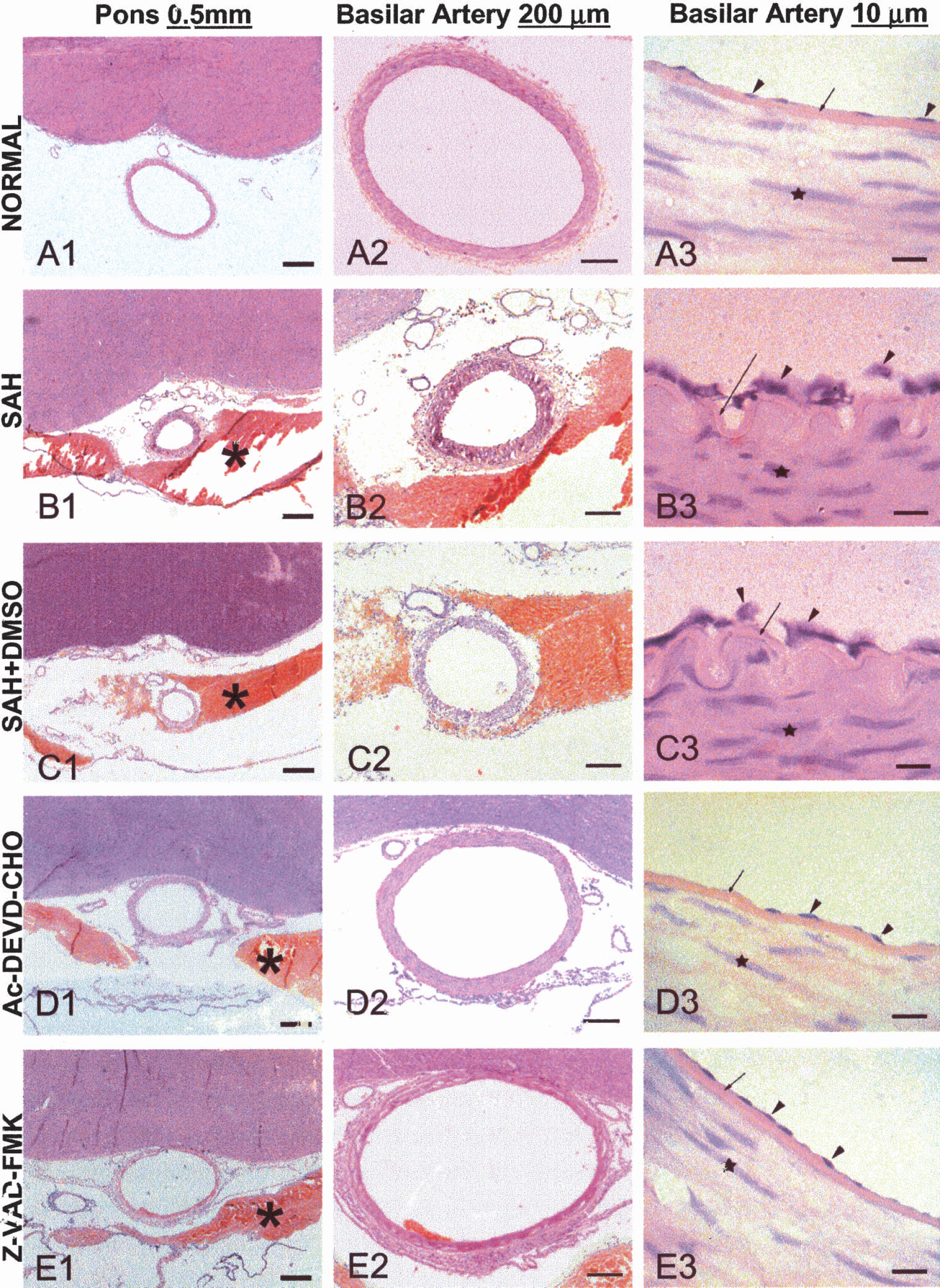
H&E staining of the basilar artery transect sections on the surface of pons.
TUNEL staining.
The TUNEL positive material was localized in the nucleus of the endothelium in the SAH and SAH+DMSO groups, which were on the inner surface of the corrugated IEL (Fig. 4A1, B1). Some of the TUNEL positive endothelium was desquamated to reveal a denuded IEL (Fig. 4A2, B2). In samples from dogs treated with Ac-DEVD-CHO and Z-VAD-FMK, no corrugation of the IEL was noticed and no positive TUNEL staining was obtained in endothelial cells (Fig. 4C, D), resembling what was observed in samples from the normal control dogs (upright insert window in Fig. 4A1). Double fluorescence immunohistochemistry staining showed TUNEL positive nuclei (green, Fig. 4E1, F1) and the positive staining of cytoplasm by Von Willibrand specific antibody (red, Fig. 4E2) as well as by the caspase-3 antibody (red, Fig. 4F2). Merging two composites into Figs. 4E3 and F3 demonstrated that TUNEL positive nuclei in the endothelium and TUNEL nucleus staining co-localized with caspase-3 staining in the cytoplasm. It needs to be noted that the IEL was autofluorescent in both red and green fluorescence in these samples.
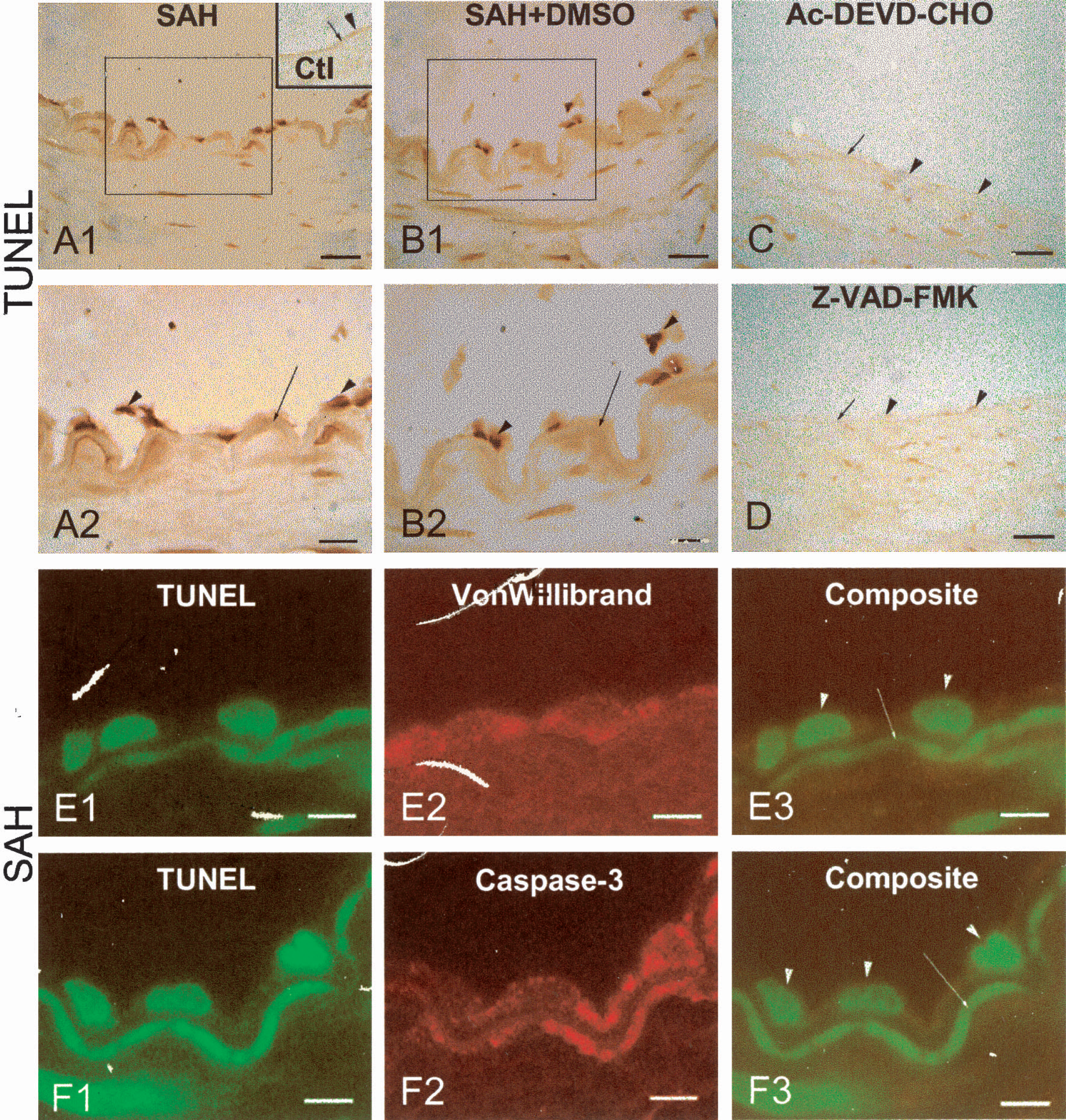
TUNEL and double fluorescence staining of dog basilar artery.
Immunohistochemistry staining.
In samples from the SAH and SAH + DMSO animals, positive staining of PARP, caspase-3, and caspase-8 was observed in endothelial cells, either on the inner surface of the IEL or desquamated from the basal lamina of the basilar arteries (Fig. 5A1, 5B1, 5C1, 5D, 5E, 5F). Higher magnification revealed that the immunoreactive materials of all three antibodies were localized in the cytoplasm of the endothelium (Fig. 5A2, 5B2, 5C2, upright insert windows in Fig. 5D, 5E, 5F). In the samples from dogs treated with the selective caspase-3 inhibitor Ac-DEVD-CHO, negative immunohistochemistry staining of caspase-3 (Fig. 5H) and its downstream PARP (Fig. 5G) was observed, but positive staining of caspase-8 was obtained (Fig. 5I and high magnification insert). Higher magnification in the insert of Fig. 5G demonstrated negative staining of PARP. On the contrary, in samples from dogs treated with the broad caspase inhibitor Z-VAD-FMK, negative immunohistochemistry staining of PARP, caspase-3, and caspase-8 was observed (Fig. 5J, 5K, 5L).
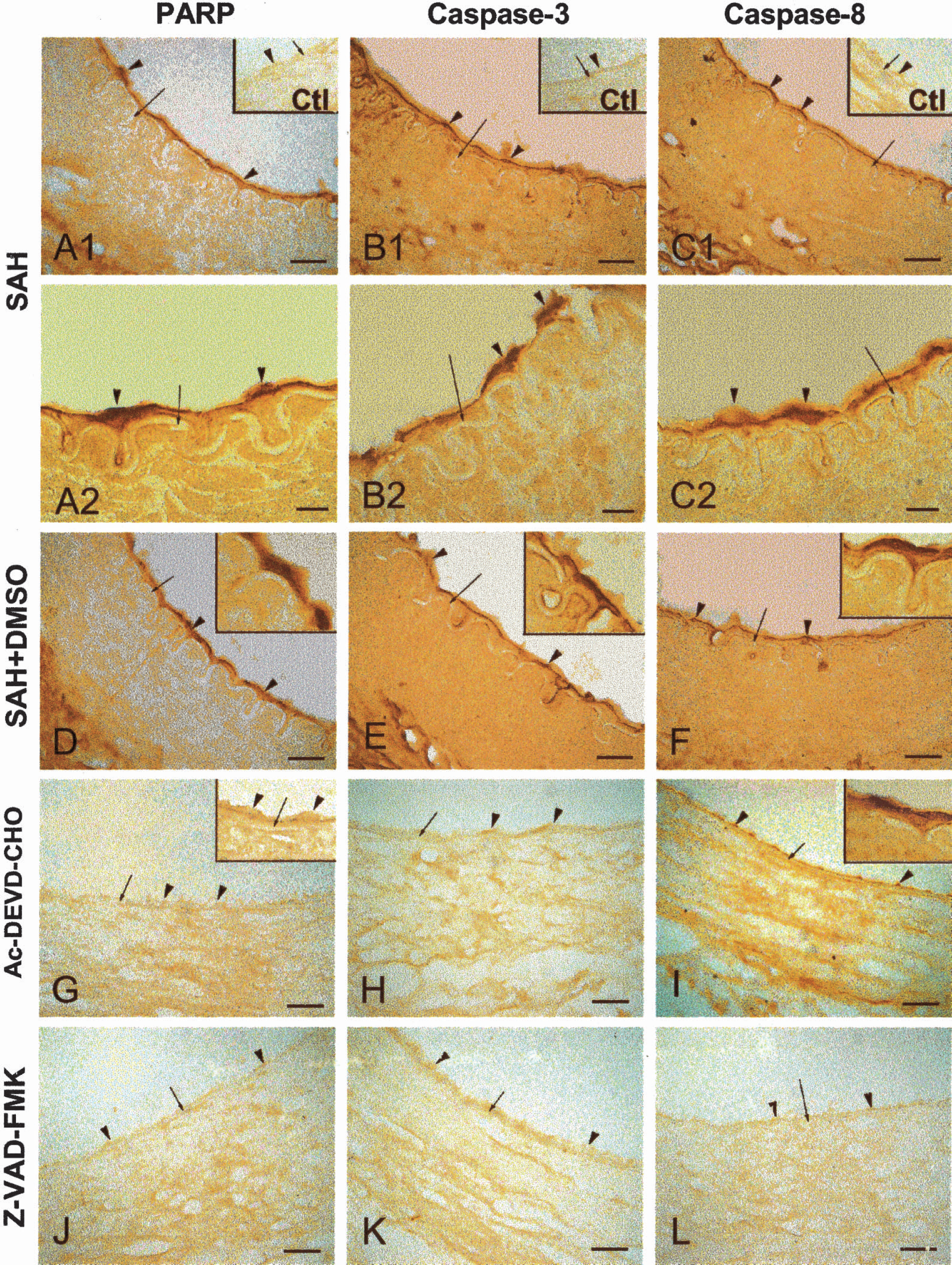
PARP, Caspase-3, and Caspase-8 immunohistochemistry in dog basilar artery.
TNFR1 immunohistochemistry staining was observed in samples from the SAH dogs (Fig. 6B) but not from the normal control dogs (Fig. 6A). Higher magnification demonstrated expression of TNFR1 in the membrane and cytoplasm of the endothelium on the inner surface of the basal lamina of the basilar artery from a SAH dog (Fig. 6C). Double fluorescence staining showed positive staining of TNFR1 in the cytoplasm of endothelial cells (red, Fig. 6D1) and positive staining of TUNEL in the nuclei (green, Fig. 6D2). Merging two composites into Fig. 6D3 demonstrates that co-localization of TNFR1 in the cytoplasm is circulated by the TUNEL positive nuclei. Similarly, double fluorescence immunohistochemistry staining of TNFR1 (red, Fig. 6E1) and caspase-3 (green, Fig. 6E2) produces a yellow color, which demonstrates co-localization in the cytoplasm of endothelial cells (Fig. 6E3).
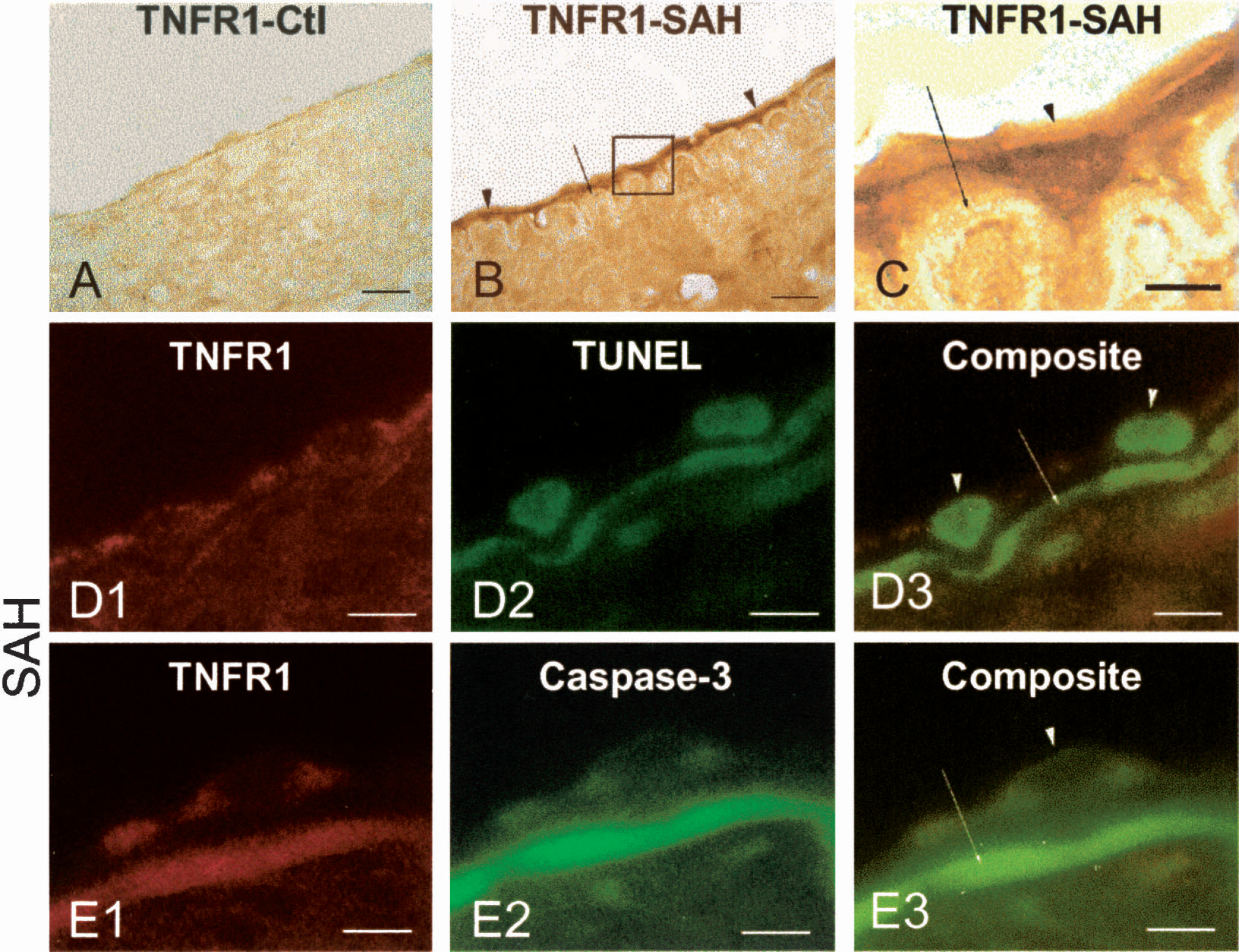
TNFα receptor-1 (TNFR1) expression in SAH dog basilar artery.
DISCUSSION
There are three major observations in the present study. First, both the selective caspase-3 inhibitor and the broad caspase inhibitor prevented apoptosis in the cerebral endothelium, attenuated angiographic vasospasm, and reduced histological vasospasm. Second, the signaling pathways for apoptosis after SAH in endothelial cells are mediated, at least partially, by TNFα-receptor-1, which recruits caspase-8, which in turn activates caspase-3, as demonstrated in other tissues (Victor and Gottlieb 2002). Activation of caspase-3 cleaves PARP and results in DNA fragmentation (Porter and Janicke 1999;Graham and Chen 2001), which was shown in the present study by positive TUNEL staining in the nucleus of the endothelial cells. This hypothesis is supported further by double fluorescence labeling which revealed co-localization of TNFR1 with caspase-3 and TUNEL nuclei, as well as co-localization of TUNEL with caspase-3. Third, caspase-3 is downstream of caspase-8, and selective inhibition of caspase-3 will block the signals from caspase-8, even though caspase-8 remains activated (Fig. 5I). This study demonstrated the central role of caspase-3 in apoptotic endothelial death and the development of cerebral vasospasm.
Endothelial apoptosis and cerebral vasospasm
The ultra-structural changes in the endothelium of cerebral arteries following SAH have been reported, including cellular distortion, formation of intracellular vacuoles, disruption of tight junctions, and widening of interendothelial spaces, which leads to detachment of endothelial cells (Findlay et al. 1989;Mayberg et al. 1990). Damaging endothelial cells after SAH may increase blood-brain barrier permeability, diminish the generation of endothelial-dependent relaxing factors, and alter the production of prostaglandin metabolites, as well as endothelin, and lead to the reduction of cerebral blood flow (Kassell et al. 1985;Clower et al. 1994;Sobey and Faraci 1998;Zhang et al. 1998). Denudation of endothelial cells exposes the smooth muscle layer to neurotransmitters, toxins, and other vasoactive agents circulating in the blood stream; enhances vasoactivity; and aggravates vasospasm since different vasogenic agents such as serotonin, uridine triphosphate, and noradrenalin interact and enhance contractility of cerebral arteries (Debdi et al. 1992;Miranda et al. 1996;Zubkov et al. 1999;Ishiguro et al. 2002;Cambj-Sapunar et al. 2003). In theory, keeping the endothelium intact will prevent those “intraluminal spasmogens” from aggravating and maintaining vasospasm. In the present study, we have demonstrated that histological vasospasm is accompanied by endothelial damage with features of apoptosis. Double labeling imaging showed TUNEL positive nuclei superimposed with caspase-3 staining in the cytoplasm (Fig. 4). The caspase inhibitors abolished endothelial apoptosis and reduced both angiographic and histological vasospasm. All of these results indicate that endothelial apoptosis may be one of the initiators and maintainers for cerebral vasospasm, and protection of the endothelium may be a potential direction in the prevention of vasospasm after SAH.
TNFα pathway in endothelial apoptosis
We have previously shown that oxyhemoglobin produced apoptosis by activating caspases-8, −9, −2, and −3 in cultured cerebral endothelial cells (Meguro et al. 2001a;Meguro et al. 2001b). In the bloody CSF from animal models of SAH, numerous agents besides oxyhemoglobin were detected and are believed to produce vasogenic or cytotoxic actions (Weir et al. 1999). Regardless of the apoptotic stimulants, three major pathways for caspase-mediated apoptosis have been proposed, including caspases downstream of TNFα/TNFR1 such as caspase-8, caspases released from mitochondria such as caspase-9, and caspases released from the endoplasmic reticulum such as caspase-12 (Nicotera 2002;Moskowitz and Lo 2003). As mentioned previously, the level of TNFα was elevated in the bloody CSF of patients suffering from SAH (Gruber et al. 2000;Fassbender et al. 2001), which, in combination with other cytokines, contributes greatly to the development of cerebral vasospasm (Dumont et al. 2003). A transient elevation of TNFα was observed in a rabbit SAH model which results in cerebral vasospasm (Tran Dinh et al. 2001). One of the functions of TNFα is to target endothelial cells (Madge and Pober 2001) and produce apoptosis in endothelial cells, including cerebral endothelial cells (Robaye et al. 1991;Hsu et al. 1997;Marino and Cardier 2003). In this study, which is consistent with other studies, we have observed that TNFα-related caspase pathways play an important role in endothelial cell death in the spastic basilar arteries. First, TNFR1 immunostaining was enhanced in endothelial cells in all SAH groups (but not in normal dogs) and co-localized with caspase-3 and TUNEL positive cells (Fig. 6). Second, we observed caspase-8 immunostaining in endothelial cells after SAH, and the broad caspase inhibitor Z-VAD-FMK abolished caspase-8 staining (Fig. 5). Interestingly, the selective caspase-3 inhibitor Ac-DEVD-CHO, which abolished the immunostaining of caspase-3, did not affect the staining of caspase-8 in cerebral endothelial cells (Fig. 5I, insert). This observation indicates that caspase-3 is downstream from caspase-8 and mediates the action of caspase-8 (Daniel 2000;Ferrer and Planas 2003). Third, activation of caspase-3 leads to cleavage of PARP and DNA fragmentation (Porter and Janicke 1999). We have shown immunostaining of caspase-3 and PARP in endothelial cells after SAH, which was abolished by both caspase inhibitors (Fig. 5). In addition, we have observed co-localization of TUNEL and caspase-3 (Fig. 4). Our data indicate that the broad caspase inhibitor Z-VAD-FMK may perform its effect at multiple locations, including caspase-3 and caspase-8. However, it is caspase-3 that plays a central role in apoptosis in endothelial cells after SAH. The selective caspase-3 inhibitor Ac-DEVD-CHO abolished endothelial apoptosis even in the presence of activated caspase-8 (Fig. 5I, insert). Even though we did not measure the level of TNFα in the bloody CSF because the ELISA kit commercially available is specific to humans and other animals but not dogs, an elevated TNFα in the bloody CSF was reported by others in patients after SAH (Gruber et al. 2000;Fassbender et al. 2001) and in a rabbit model of cerebral vasospasm (Tran Dinh et al. 2001). In addition it needs to be noted that apoptotic cell death, either in the vascular system or in the neuronal tissues, might trigger further inflammatory responses to or aggravate cerebral vasospasm. Caspase inhibitors that prevent or reduce apoptosis might have the potential to prevent this “secondary inflammatory response.”
Caspase inhibitors & cerebral vasospasm
Since caspases play an important role in apoptosis, caspase inhibitors have emerged as new therapeutic strategies for biological disorders, including neurological diseases (Robertson et al. 2000;Reed 2001;Onteniente et al. 2003). Caspase-3, as a downstream central player for several apoptotic pathways, has attracted attention, especially for stroke and neurodegenerative diseases (Schulz et al. 1999;Porter and Janicke 1999). We have previously conducted preliminary studies using different caspase inhibitors in a similar canine double-hemorrhage model, and we examined both major cerebral arteries and penetrating arterioles but achieved mixed results (Zubkov et al. 2002a;Aoki et al. 2002). One of the caspase inhibitors achieved “near-significant” reduction of cerebral vasospasm by 13% (Aoki et al. 2002). From this experience, we recommended more potent inhibitors, earlier treatment, and a higher concentration for future studies (Aoki et al. 2002). Thus, the present study was designed to correct the weaknesses in previous studies by using two new caspase inhibitors, one selective for caspase-3 and one with broader actions. We administered the caspase inhibitors on Day 0 and continued daily to Day 3 instead of using a delayed treatment from Day 2-Day 6. We used a higher concentration of caspase inhibitors (100 μM), which is ten times more than the previous study (Aoki et al. 2002). This new strategy improved the outcome remarkably by preventing apoptosis in endothelial cells and by reducing angiographic vasospasm as well as histological vasospasm. The effect of caspase inhibitors was likely mediated by anti-apoptosis as shown in the immunohistochemistry studies, but not by a direct vasodilator effect (Fig. 1C). Both caspase inhibitors improved clinical behavior, which was scored by appetite, activity, and neurological deficit. An interesting observation was that clinical scores for the SAH and SAH+DMSO dogs were reduced (dogs were improving) with time, especially after Day 4, while dogs treated with caspase inhibitors showed an opposite tendency—an initially good score in the first 2–3 days and becoming more fluctuated (deteriorated) at Day 3–4 (Fig. 2). This phenomenon may indicate a possible “toxic” effect of these caspase inhibitors, even though the level of dysfunction is extremely limited.
Footnotes
Acknowledgment
We would like to thank Caraline Domb for editing this article.