
Abstract
Cerebral vasospasm is a major cause of morbidity and mortality after aneurysmal subarachnoid hemorrhage (SAH). It is a sustained constriction of the cerebral arteries that can be reduced by endothelin (ET) receptor antagonists. Voltage-gated Ca2+ channel antagonists such as nimodipine are relatively less effective. Endothelin-1 is not increased enough after SAH to directly cause the constriction, so we sought alternate mechanisms by which ET-1 might mediate vasospasm. Vasospasm was created in dogs, and the smooth muscle cells were studied molecularly, electro-physiologically, and by isometric tension. During vasospasm, ET-1, 10 nmol/L, induced a nonselective cation current carried by Ca2+ in 64% of cells compared with in only 7% of control cells. Nimodipine and 2-aminoethoxydiphenylborate (a specific antagonist of store-operated channels) had no effect, whereas SKF96365 (a nonspecific antagonist of nonselective cation channels) decreased this current in vasospastic smooth muscle cells. Transient receptor potential (TRP) proteins may mediate entry of Ca2+ through nonselective cationic pathways. We tested their role by incubating smooth muscle cells with anti-TRPC1 or TRPC4, both of which blocked ET-1-induced currents in SAH cells. Anti-TRPC5 had no effect. Anti-TRPC1 also inhibited ET-1 contraction of SAH arteries in vitro. Quantitative polymerase chain reaction and Western blotting of seven TRPC isoforms found increased expression of TRPC4 and a novel splice variant of TRPC1 and increased protein expression of TRPC4 and TRPC1. Taken together, the results support a novel mechanism whereby ET-1 significantly increases Ca2+ influx mediated by TRPC1 and TRPC4 or their heteromers in smooth muscle cells, which promotes development of vasospasm after SAH.
Keywords
Introduction
Subarachnoid hemorrhage (SAH) from a ruptured cerebral aneurysm affects 10 of every 100,000 people each year. Although it is less common than other forms of stroke, the younger mean age of those stricken results in disproportionately high cost in terms of mortality, morbidity, and cost to society. Of patients who survive to reach hospital, the causes of poor outcome include the initial effects of the SAH and delayed cerebral vasospasm. Because it does not occur immediately, vasospasm is potentially the most treatable component. Vasospasm is constriction of the large, subarachnoid conducting arteries of the circle of Willis that develops days after SAH, which can cause cerebral ischemia and infarction. Treatment includes the voltage-gated calcium (Ca2+) channel antagonist, nimodipine, and hemodynamic maneuvers to optimize cerebral blood flow. Many other therapeutic measures have been investigated but understanding of the pathophysiology of vasospasm is incomplete and has hampered development of effective treatment.
A key process involved in vasospasm is smooth muscle contraction, which is known to be dependent on increased intracellular Ca2+. Other processes may be involved in contraction, but sustained contraction of cerebrovascular smooth muscle is dependent largely on influx of Ca2+ through voltagegated Ca2+ channels or Ca2+-permeable nonselective cation channels (Karaki et al, 1997; Inoue et al, 2006). Voltage-gated Ca2+ channel antagonists have been and continue to be studied for prevention of vasospasm but results have been conflicting (Haley et al, 1993; Kasuya et al, 2005). Conversely, recent randomized, masked, placebo-controlled trials show for the first time marked efficacy of endothelin A (ETA) receptor antagonists against vasospasm in humans (Vajkoczy et al, 2005; Macdonald et al, 2006). Furthermore, these ETA receptor antagonists were effective at reducing vasospasm after SAH in dogs (Macdonald et al, 1998) and other species (Hino et al, 1995). ET-evoked contractions are believed to be mediated in large part by Ca2+ influx through nonselective cation channels (Iwamuro et al, 1999). Proteins mediating nonselective cation channel currents are not known but promising candidates include those of the transient receptor potential (TRP) family (Bergdahl et al, 2003; Miwa et al, 2005). Therefore, we hypothesized that vasospasm might be mediated by ET-induced increase in Ca2+ influx via TRP proteins. We tested this in a widely used dog model of SAH and vasospasm.
Materials and methods
Animal Model
Mongrel dogs weighing 15 to 25 kg were randomly assigned to serve as controls or to undergo SAH and be euthanized 4 or 7 days after SAH (Macdonald et al, 1998). Animals were anesthetized with intravenous sodium pentothal (15 mg/kg) and then intubated and ventilated on oxygen and 1% to 2% isoflurane. A cerebral angiogram was obtained. Fresh autologous arterial nonheparinized blood of 0.5 mL/kg was injected into the cisterna magna and animals tilted 30° head down during injection and for 15 mins thereafter. Blood injection was repeated 2 days later, and angiography and euthanasia was carried out 4 or 7 days after the first angiogram. Control dogs had 0.9% NaCl injected into the cisterna magna instead of blood and were euthanized on day 7. Experiments complied with procedures for the care and use of animals established by the National Institutes of Health and was approved by the Institutional Animal Care and Use Committee.
Smooth Muscle Cell Isolation
Animals were euthanized under general anesthesia by exsanguination and transcardiac perfusion with cold phosphate-buffered saline (pH 7.4) at physiological intraluminal pressure. The brain was excised and placed in 4°C phosphate-buffered saline. The basilar artery was isolated under a microscope, and the endothelial layer was removed by passing a wire of the same diameter as the artery through the arterial lumen and then flushing the artery with phosphate-buffered saline. The basilar artery was placed in a solution of (in mmol/L): 130 NaCl; 5 KCl; 1.3 MgCl2; 10 N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid; 7 glucose, penicillin (100 U/mL), and streptomycin (0.1 mg/mL) with pH 7.4 and osmolarity of 300 mosm/L. The artery was cut into ~1 mm2 pieces, which were incubated for 30 mins at 37°C in the same solution plus (mg/mL): 10 elastase; 1.1 collagenase type 4; 0.76 soybean trypsin inhibitor, and 0.1 deoxyribonuclease-1 dissolved in Ca2+-free dissection solution (enzymes from Worthington Biochemical Corp., Lakewood, NJ, USA). Supernatant fluid was discarded, the tissue washed in the original solution containing 0.2% bovine serum albumin and the smooth muscle cells released by trituration.
Electrophysiology
Voltage-induced currents were recorded in the whole-cell configuration of the patch-clamp technique under conditions where voltage-gated Ca2+ and K+ channel currents would be blocked (Jung et al, 2003). Extracellular solution was (in mmol/L): 140 NaCl, 5 CsCl, 1 MgCl2, 2 CaCl2, 10 N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid, and 10 glucose with pH 7.4 adjusted with NaOH. Osmolarity was 323 mosm/L. Patch pipettes were fabricated from borosilicate glass tubes on a micropipette puller (Sutter P-97, Sutter Instrument Co., Novato, CA, USA) and were fire polished. Pipette resistance was 2 to 3 MΩ when filled with the pipette solution of (in mmol/L): 110 Cs methanesulfonate, 25 CsCl, 2 MgCl2, 3.6 CaCl2, 30 N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid, 10 ethylene glycol-bis-(b-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (pH 7.2 with CsOH) with a calculated [Ca2+] of 100 nmol/L. Experiments were at 23°C.
Currents were evoked from a holding potential of 50 mV. Ramp protocols were from −100 to + 100 mV over 200 ms. A 30 ms step to −100 mV was used before the ramp was applied. Records were filtered at 0.5 kHz. Digitization was at 1 kHz during ramps and 25 Hz at 10 s intervals between ramps. Data were acquired with Clampex 6.0.4 on an Axonpatch-1D amplifier and a Digidata 1200 interface (Axon Instruments Inc., Sunnyvale, CA, USA).
Isometric Tension Recordings
The basilar artery was denuded of endothelium and cut into rings 3 mm long and rings were mounted on metal hooks immersed in tissue baths on a custom-built isometric tension recording device (Young et al, 2007). Baths contained Krebs—Henseleit buffer (in mmol/L: NaCl 123, KCl 5.9, CaCl2 2.5, MgSO4 1.2, NaHCO3 25, KH2PO4 1.2, and glucose 11), bubbled with 95% O2 and 5% CO2, pH 7.4, at 37°C. The diameter of the basilar artery in vivo was calculated by measurement of arterial diameters on angiograms, which were corrected for magnification and the arteries were stretched to the equivalent of their diameter in vivo. Rings were contracted at intervals with KCl, 60 mmol/L until stable contractions were obtained and then studied.
Molecular Biology
Endothelial cells were removed from the basilar arteries. Total RNA, genomic DNA, and protein were extracted with TRIzol (Gibco BRL, Life Technologies, Rockville, MD, USA). RNA and protein were used to determine specific gene expression. Genomic DNA was quantified in a Gene Spec III (HITACHI Genetic Systems, Alameda, CA, USA) and was used as internal control to standardize RNA levels because standard housekeeping genes such as β-actin and glyceraldehyde-3-phosphate dehydrogenase are altered after SAH (RL Macdonald, unpublished observations) (Ohkuma et al, 2001).
RNA samples were treated with RNase-free DNase (Promega, Madison, WI, USA) to eliminate contamination with genomic DNA. RNA reverse transcription was performed with SUPER SCRIPT First-Strand Synthesis System (Invitrogen Life Technologies, Carlsbad, CA, USA). RNA (5 μg) was used for reverse transcription in a reaction mixture consisting of 50ng of random hexamers per 1 μg total RNA. RNase H was used to remove the remaining RNA from the reverse transcription product. Dog-specific sequences of mRNAs for canonical TRP channels were not found in the NCBI database (http://www.ncbi.nlm.nih.gov/Entrez) so partial nucleotide sequences for these genes were determined using degenerate primers based on other species. Poymerase chain reaction products were sequenced (Applied Biosystems 3700 96-capillary automated DNA sequencer; Foster City, CA, USA) and specific dog sequences were used to design specific primers and probes for quantitative real-time PCR (Table 1), which was performed with real-time TaqMan technology using a Sequence Detection System model 7700 (Applied Biosystems). RNA concentrations were normalized to genomic DNA. Splice variants of TRPC1 were identified using primer pairs (5-GAAGCTTTTCTTGCTGGCGT-3 sense and 5-GACAGGTGCAACATCCATAGTTG-3 antisense) common to all four TRPC1 splice variants. Poymerase chain reaction conditions were 40 cycles at 93°C for 30secs, 60°C for 30 secs and 72°C for 1min. Products were purified and sequenced to confirm their identity.
Oligonucleotide sequences of dog-specific primers and probes used for real-time quantitative PCR
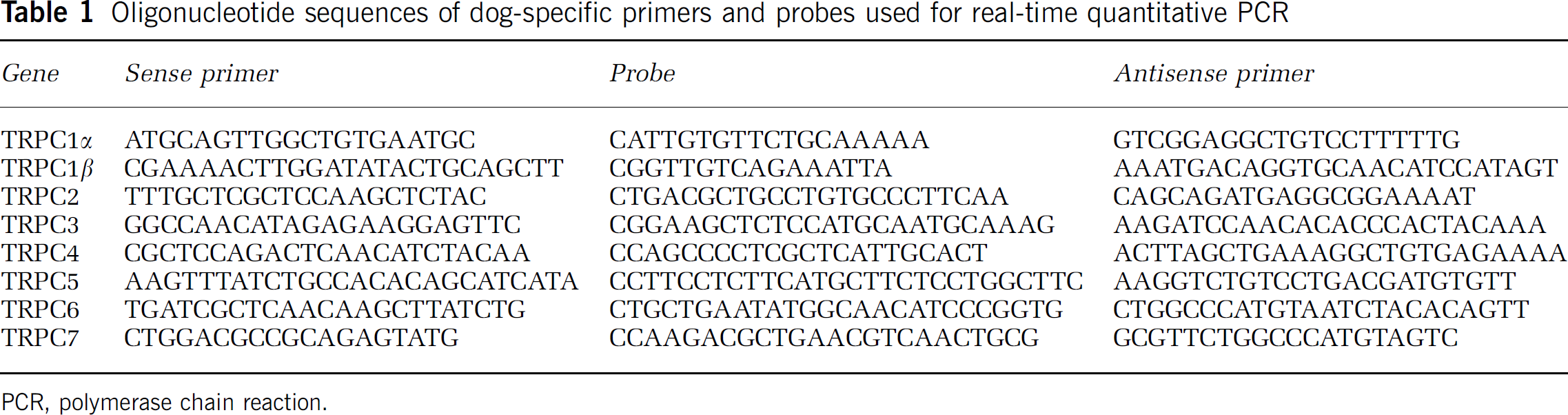
PCR, polymerase chain reaction.
Western Blotting
Protein (10 μg per basilar artery) was diluted in 1% sodium dodecyl sulfate (SDS) buffer (250 mmol/L Tris-HCl, pH 6.8, 10% SDS, and 10% glycerol) and separated on a 1.2% Tris-glycine gel (Invitrogen) in SDS/polyacrylamide gel electrophoresis running buffer for 1 h at 160 V. Proteins were then transferred onto methanol-soaked Hybond P (polyvinylidene difluoride) membranes in buffer containing 3-[cyclohexylamino]-1-propanesulfonic acid and 10% methanol for 2 h at 60 V. For blotting, filters were first incubated with 5 % skimmed milk in Tris-buffered saline with Tween 20 (20 mmol/L Tris-Cl, pH 7.6, 137 mmol/L sodium chloride, 0.1% Tween 20) at room temperature for 1 h to block nonspecific binding. Filters were incubated with antibodies to TRPC1, 3, 4, 5, or 6 (rabbit immunoglobulin G, 1:200, Alomone Labs, Jerusalem, Israel) or mouse monoclonal anti-TRP1 antibody (1:1,000 dilution, from L Tsiokas, University of Oklahoma Health Science Center, OK, USA). For negative controls, a blocking peptide was preincubated with the antibody before applying to the membrane. After washing, the membranes were incubated for 1 h with horseradish peroxidase-conjugated immunoglobulin G anti-rabbit antibody (1:2,500 dilution; Cell Signaling Technology, Beverly, MA, USA) for TRPC1, 3, 4, 5, 6, or horseradish peroxidase-conjugated anti-mouse antibody (1:2000 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for TRPC1 with primary mouse monoclonal anti-TRPC1 antibody. Proteins were visualized with the enhanced chemiluminescence Plus detection system (Amersham, Piscataway, NJ, USA). Total protein loaded in each lane was measured by staining the transferred polyvinylidene difluoride membrane with GelCode Blue Stain Reagent (Pierce, Rockford, IL, USA).
Translation In Vitro
Primers (5-ATGATGGCGGCCCTGTACCCG-3 sense and 5-TTAATTTCTTGGATAAAACAT-3 antisense) common to the end of the open reading frame of many species of TRPC1 were designed. Poymerase chain reaction conditions consisted of 40 cycles at 95°C for 30 secs, 50°C for 30secs, and 68°C for 3 mins with Advantage 2PCR Kit (BD Bioscience). Poymerase chain reaction fragments were purified, subcloned into pSTBlue-1 vector (Navagen, Madison, WI, USA) and sequenced. We selected the cDNAs encoding β variant of the dog TRP1 full sequences with 55 base pair insertion. In vitro translation was performing using a 2,4,6-trinitrotoluene coupled Reticulocyte Lysate System (Promega Madison, WI, USA). Plasmid DNA (2 μg) was translated (reaction volume, 50 μL) using SP6 RNA polymerase 30°C for 1 h. Denatured translation reaction (2 μL) was diluted to 15 μL in 10% SDS glycerol and loaded onto a 12% SDS-polyacrylamide gel. After electrophoresis and transfer to the membrane (same conditions as Western blotting), translation products were detected with the Transcend Non-Radioactive Translation Detection System (Promega Madison, WI, USA).
Data Analysis
Electrophysiology data were analyzed with Clampfit 8.2 (Axon Instruments), Origin 6.1, and Excel 2000. Results are shown as mean ± s.e. with n being the number of cells examined. All data were derived from three or more separate animals. The molecular biology was carried out on seven controls, and seven dogs with SAH and the electrophysiology and isometric tension results were obtained from 5 controls, 5 dogs 4 days and 7 dogs 7 days after SAH. Student's t-test, χ2, and analysis of variance tests were used to determine statistical significance. P < 0.05 was considered significant.
Results
Angiographic Vasospasm
Creation of SAH in dogs caused significant reduction in basilar artery diameter 4 (45% ± 7% decrease, P < 0.001, paired t-test, n = 5) and 7 days (55% ± 3% decrease, P < 0.001, paired t-test, n = 14) after SAH. Basilar artery diameter did not change in control dogs (n = 12) that underwent the same procedures except that physiological saline and not blood was injected into the subarachnoid space. There were no significant differences in physiological parameters between groups at each time or within groups over time (data not shown).
Increased Sensitivity of Vasospastic Smooth Muscle to Endothelin-1
Our prior experiments showed that vasospasm in dogs was markedly reduced by treatment with an ETA receptor antagonist (Macdonald et al, 1998). However, ET-1 concentration in the cerebrospinal fluid and basilar arteries of these dogs was only mildly increased. Therefore, we examined whether vasospastic smooth muscle was more sensitive to ET-1 than normal smooth muscle. This was first studied on smooth muscle cells isolated from the dog basilar artery using whole-cell patch clamp electrophysiology under conditions designed to elicit nonselective cation currents. Endothelin-1, 10 nmol/L, had no effect on inward and outward currents in 26 of 28 cells (93%) from control dogs (Figure 1). The same concentration of ET-1, however, induced large inward and outward currents in 19 of 32 cells (59%) from dogs 4 days and 16 of 25 cells (64%) from dogs 7 days after SAH (Figure 1). Thus, there was an association between increasing vasospasm and increasing percentage of cells sensitive to ET-1. The effect of ET-1 was first noted at 1 nmol/L (data not shown). These results show that cells from dogs after SAH are more sensitive to ET-1 than normal cells and this is associated with vasospasm.
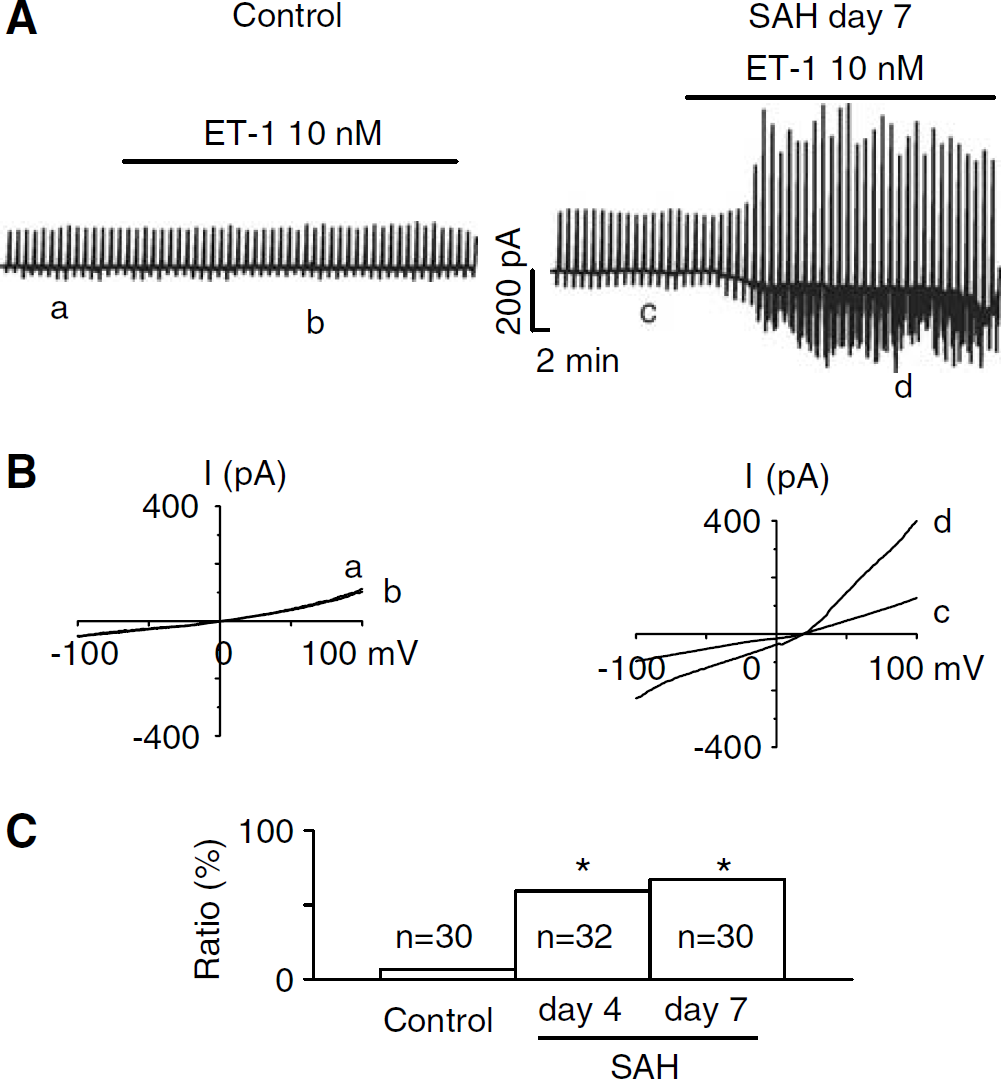
Endothelin-1, 10 nmol/L, significantly enhanced inward and outward currents in smooth muscle cells from dog basilar arteries 4 and 7 days after SAH. (
Nature of Currents Evoked by Endothelin-1
Because this current might mediate vasospasm after SAH, we characterized it further in cells from dogs 7 days after SAH. Replacing Na+ in the extracellular solution with N-methyl-
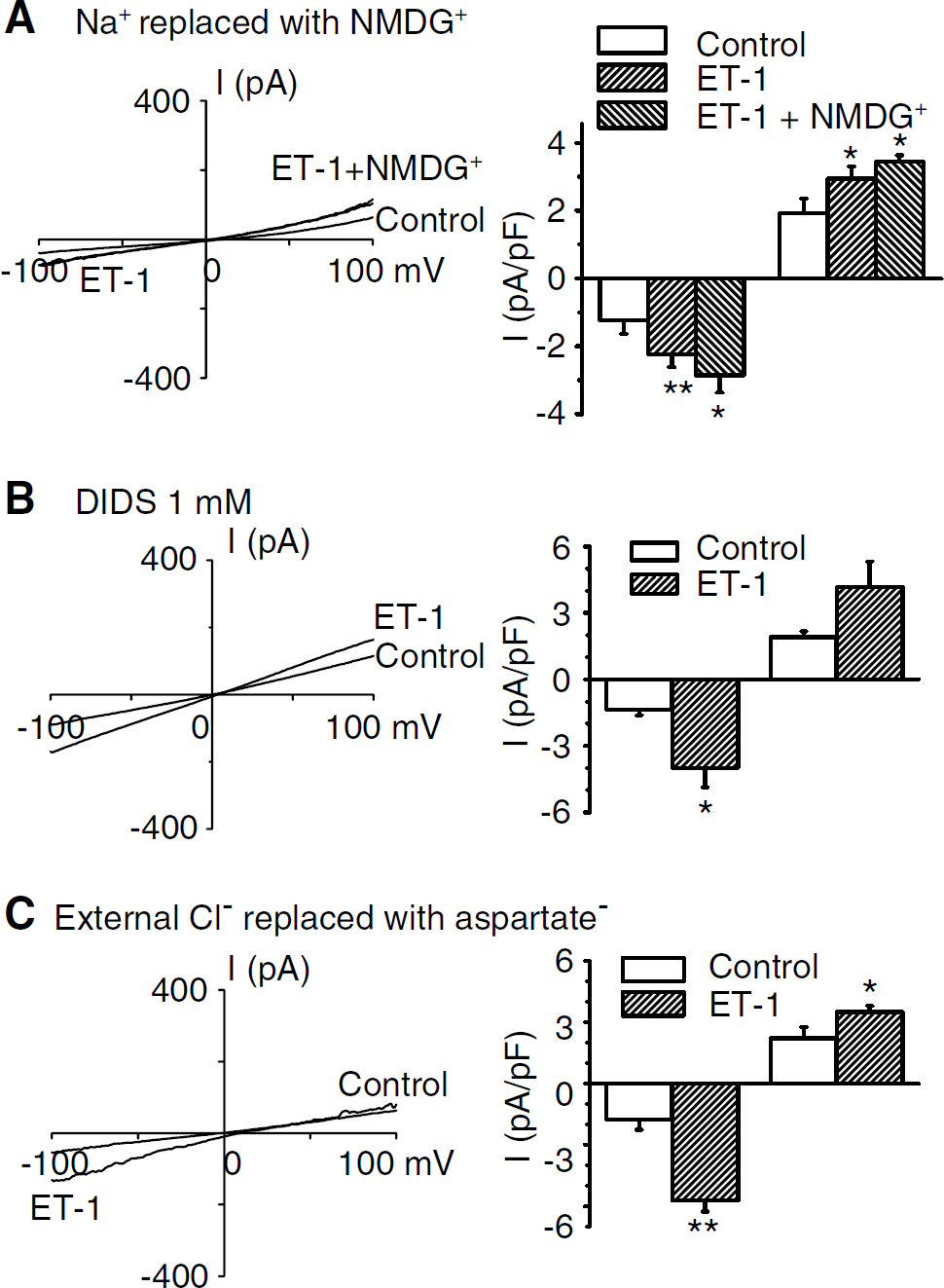
Ionic characteristics of ET-1-induced current in cells from dogs 7 days after SAH. (
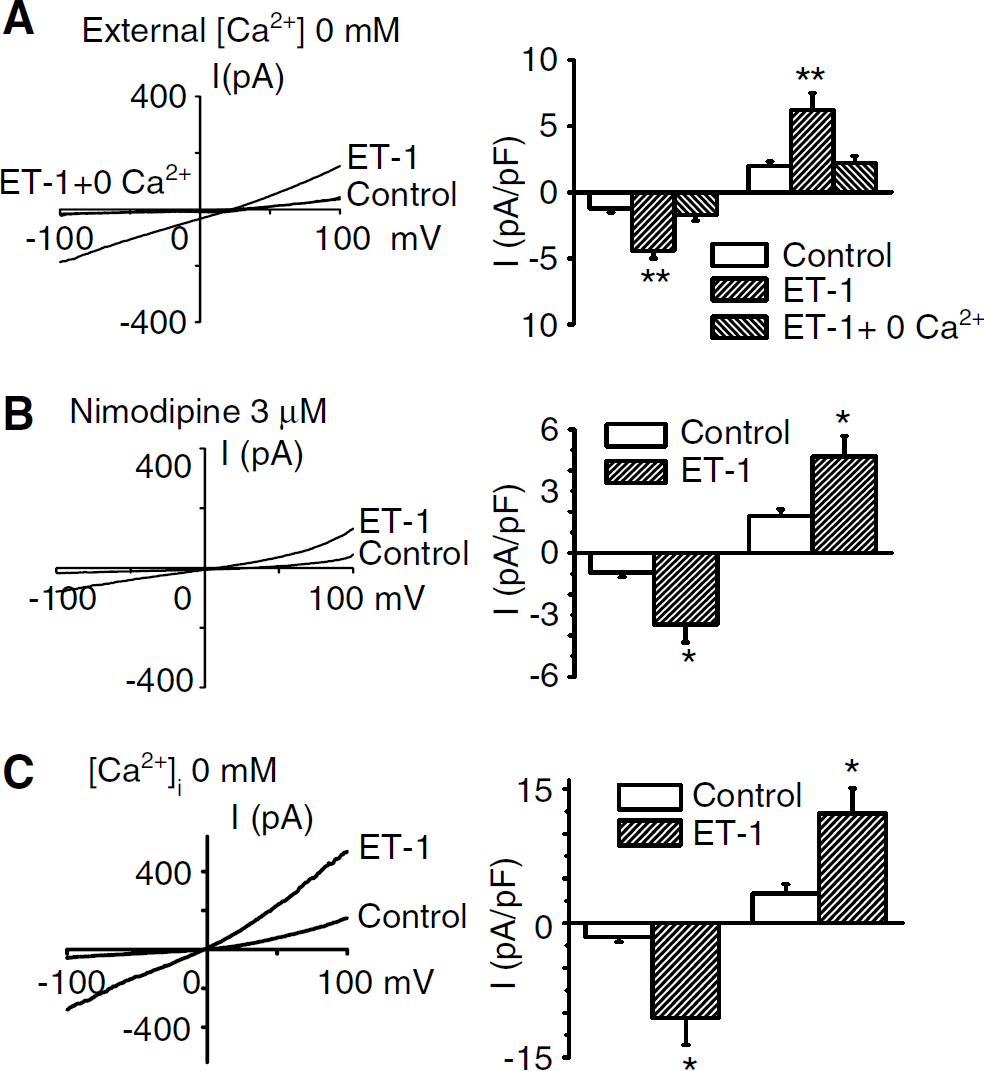
Role of Ca2+ in ET-1-evoked currents in cells from dogs 7 days after SAH. (
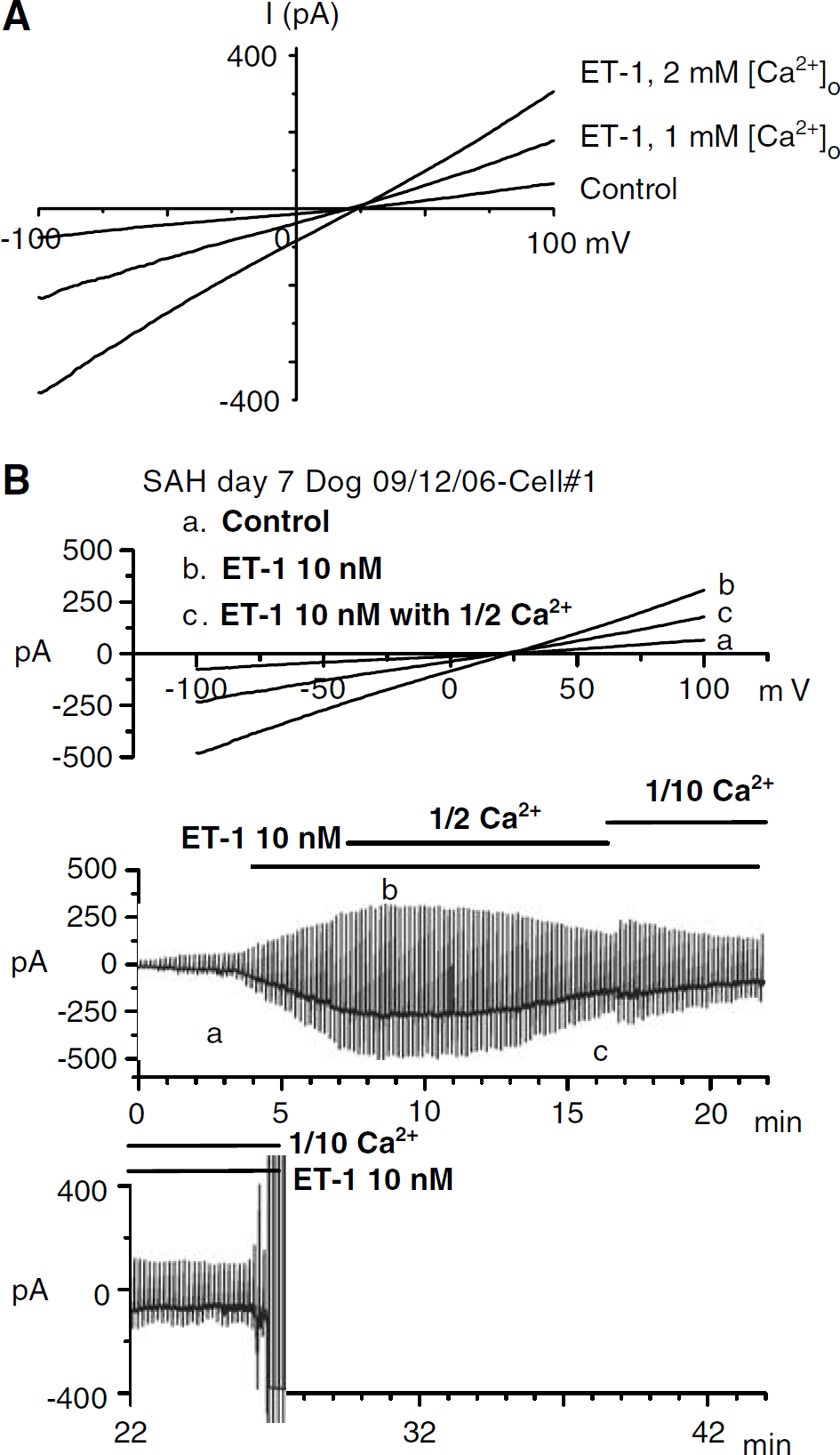
Effect of reduced extracellular Ca2+ on ET-1-induced current in cells from dogs 7 days after SAH. (
Next, we examined pharmacologic characteristics of ET-1-evoked current. 2-aminoethoxydiphenylborate 75 μmol/L) did not (Figure 5A), whereas SKF96365 (10 μmol/L) did significantly decrease current (Figure 5B). Both 2-aminoethoxydiphenylborate and SKF96365 antagonize nonselective cation currents that may be mediated by TRP proteins (Trebak et al, 2003; Sydorenko et al, 2003). 2-aminoethoxydiphenylborate may be a more specific antagonist of store-operated channels, whereas SKF96365 is less specific. Therefore, these results suggest that ET-1-induced Ca2+ influx is not through store-operated Ca2+channels but through nonselective cation channels, perhaps receptor-operated channels.
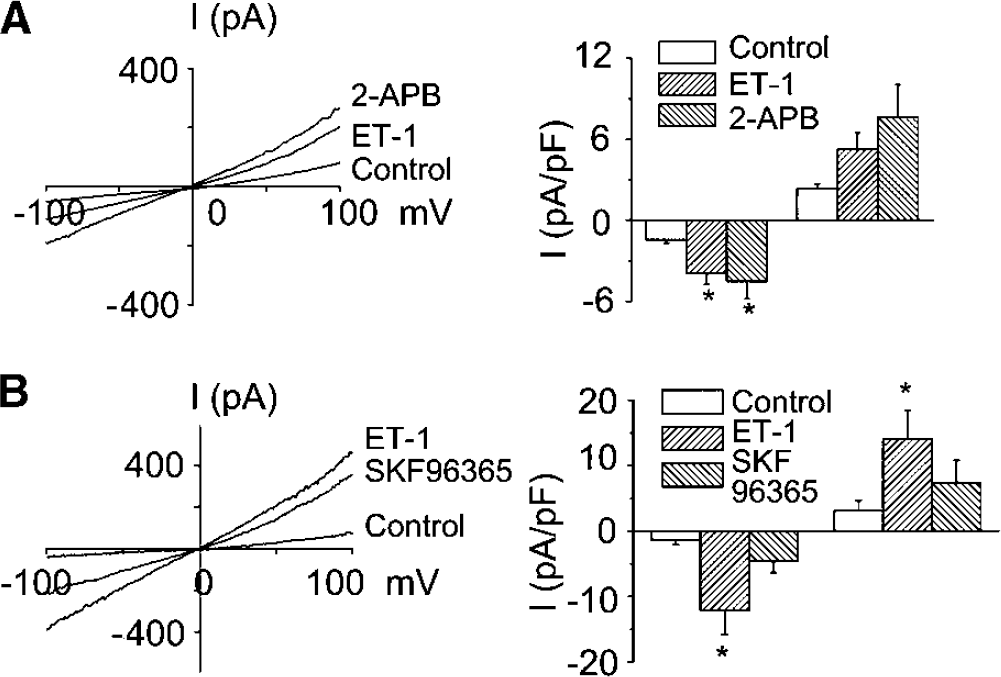
Pharmacologic characteristics of ET-1-induced current in cells from dogs 7 days after SAH. (
Transient Receptor Potential Proteins Involved in Endothelin-1 Currents
We next tested whether antibodies to various TRPC isoforms inhibited currents evoked by ET-1. Antibody to TRPC1, which was generated to a peptide corresponding to amino acids 557 to 571 of human TRPC1 between the fifth transmembrane segment and the pore loop was added to the extracellular solution (10 μg/mL) for 30 mins. Antibody to TRPC1, which was generated to a peptide corresponding to amino acids 943 to 953 of mouse TRPC4 in the intracellular C-terminus, was added to the pipette solution (10 μg/mL) for 30 mins. Both anti-TRPC1 and anti-TRPC4 inhibited inward and outward currents owing to ET-1 (10 nmol/L, Figures 6A and 6B, n = 7 and four SAH cells, respectively). An antibody to amino acids 959 to 973 of human TRPC5, in the intracellular C-terminus added to the extracellular and pipette solutions (10 μg/mL), however, had no effect on currents evoked by ET-1, 10 nmol/L (Figure 6C). This suggests that ET-1-induced Ca2+ influx involves TRPC1 and TRPC4 or their heteromultimers but not TRPC5.
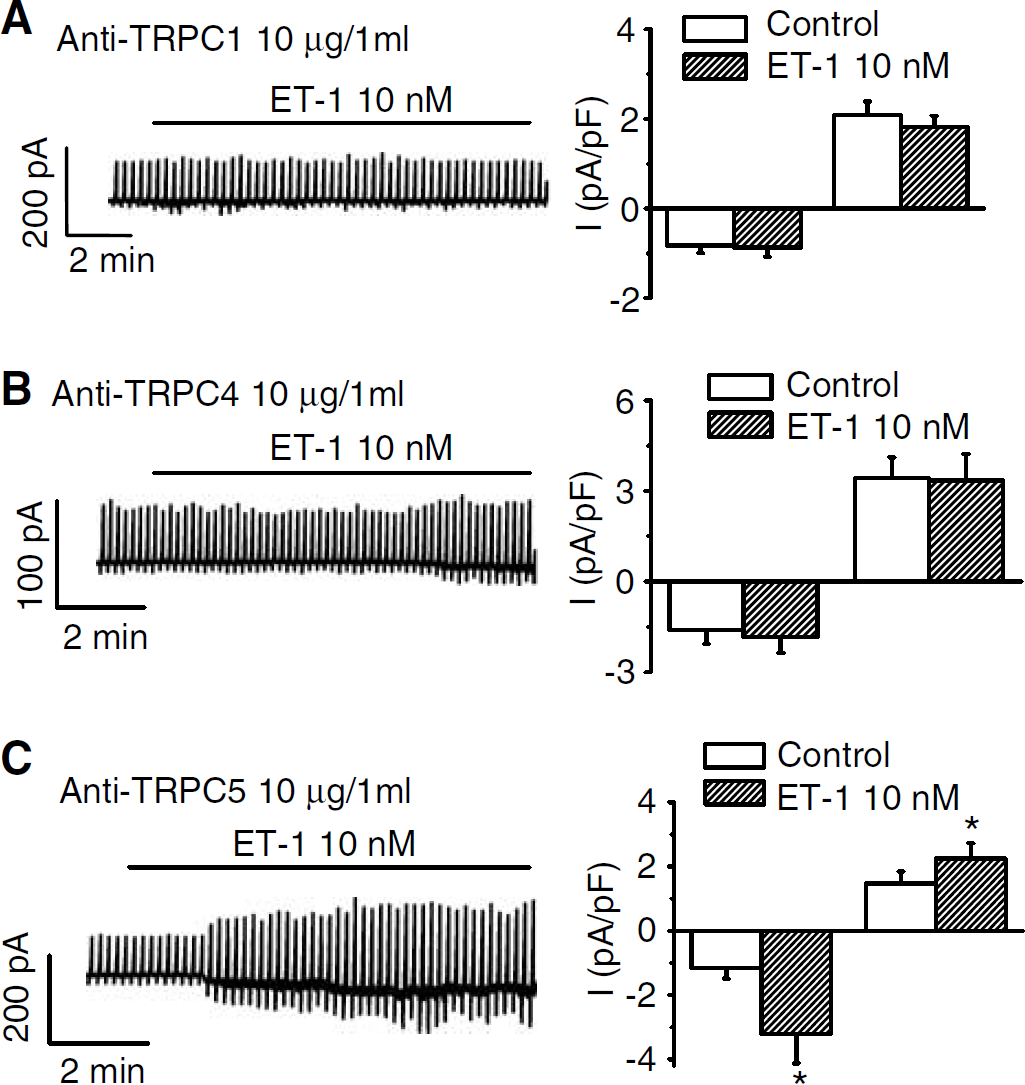
Effect of TRPC antibodies on ET-1-induced currents. Cells were exposed to (
Transient Receptor Potential C1 Mediates Endothelin-1 Contraction In Vitro
To further assess the functional importance of TRPC1 and TRPC4, control basilar arteries and arteries with vasospasm 7 days after SAH were exposed to increasing concentrations of ET-1 in the presence or absence of anti-TRPC1 or TRPC5 antibodies (10 μg/mL in extracellular bath solution for 30 mins) or SKF96365 (10 μmol/L). Vasospastic arteries were significantly more sensitive to ET-1 (Figure 7). Contractions to ET-1 also were inhibited and reduced to control level by both anti-TRPC1 or SKF96365.
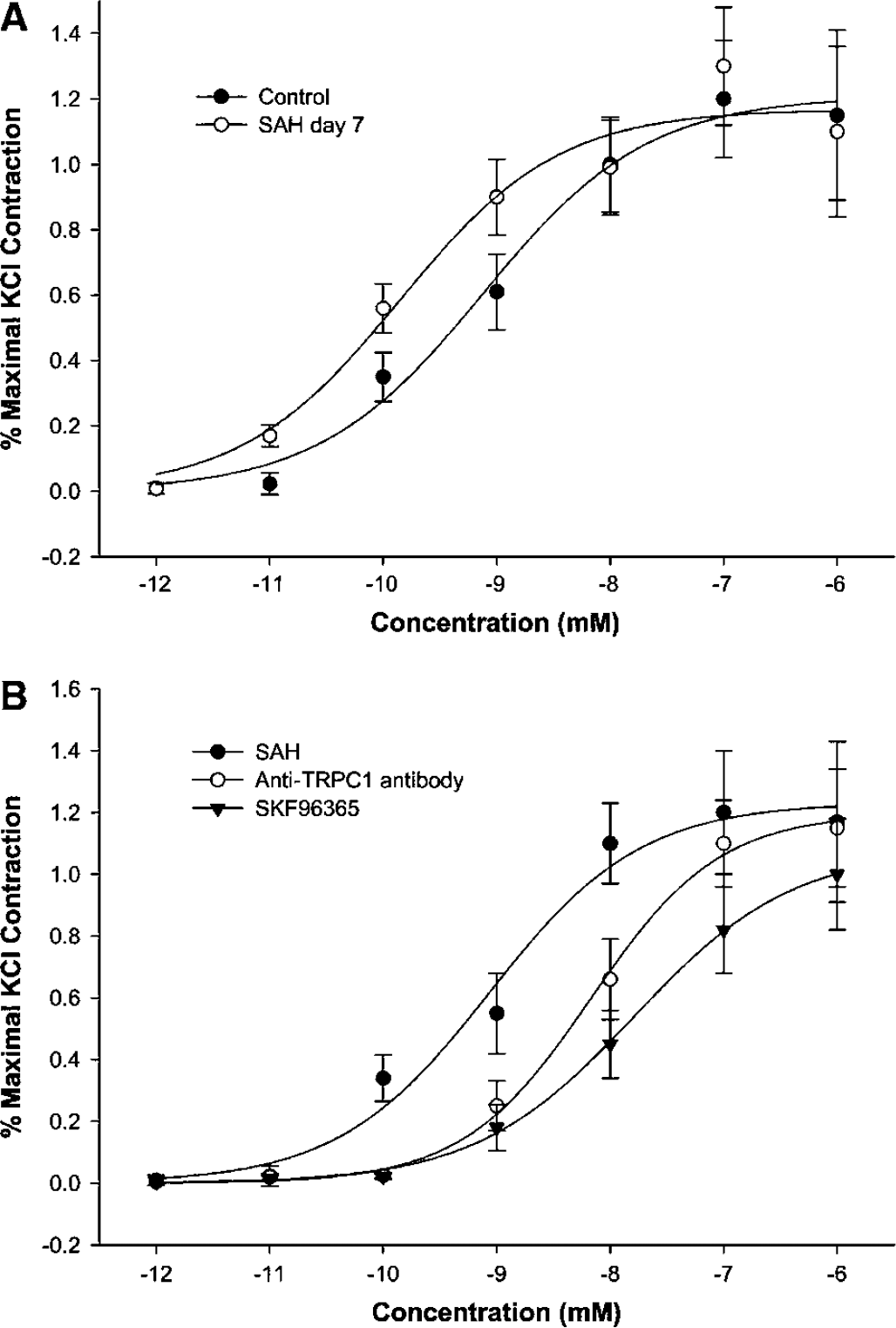
Contractions of control and day 7 SAH basilar artery to ET-1 in the presence or absence of anti-TRPC1 or TRPC5 antibodies (10 μg/mL) or SKF96365 (10 μmol/L). Contraction is shown on the y-axis as a percent of maximal contraction to KCl and final bath concentration of ET-1 is shown on the x-axis in mmol/L. (
Molecular Basis for Endothelin-1 Currents
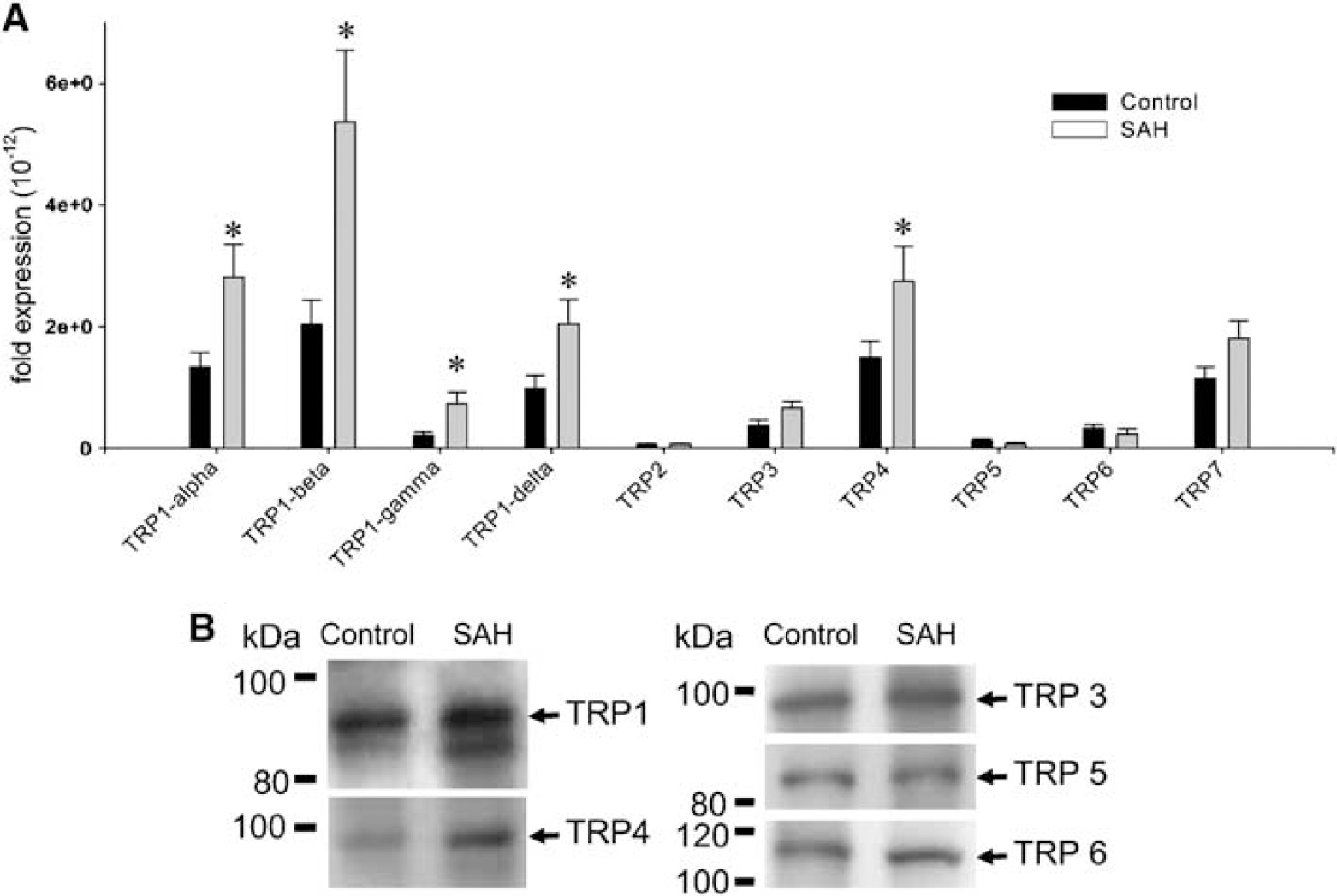
The electrophysiological and functional data suggested increased ET-1 currents were mediated by TRPC1 and TRPC4. We therefore determined whether there were changes in these proteins during vasospasm after SAH. Four splice variants of TRPC1 (α, β, γ, and δ) have been described in mice but little is known about TRP proteins in dogs (Walker et al, 2001). Therefore, we first designed primers to highly conserved positions to determine which of the four TRPC1 isoforms were present in dogs (Figure 8A). These primers distinguished four TRPC1 variants in dog brain and basilar artery. Using these primers and primers for TRPC2, 3, 4, 5, 6, and 7, real-time PCR was performed on normal control basilar artery and basilar artery with vasospasm 7 days after SAH. Levels of TRPC1 and 4 were significantly increased after SAH (Figure 9A). Protein levels of TRPC3 (98 kDa), 5 (85 kDa), and 6 (110 kDa) were not altered after SAH compared with control basilar artery and no signal was present when using a blocking peptide (Figure 9B). Transient receptor potential C1 (98 kDa) and 4 protein (97 kDa) were increased after SAH.
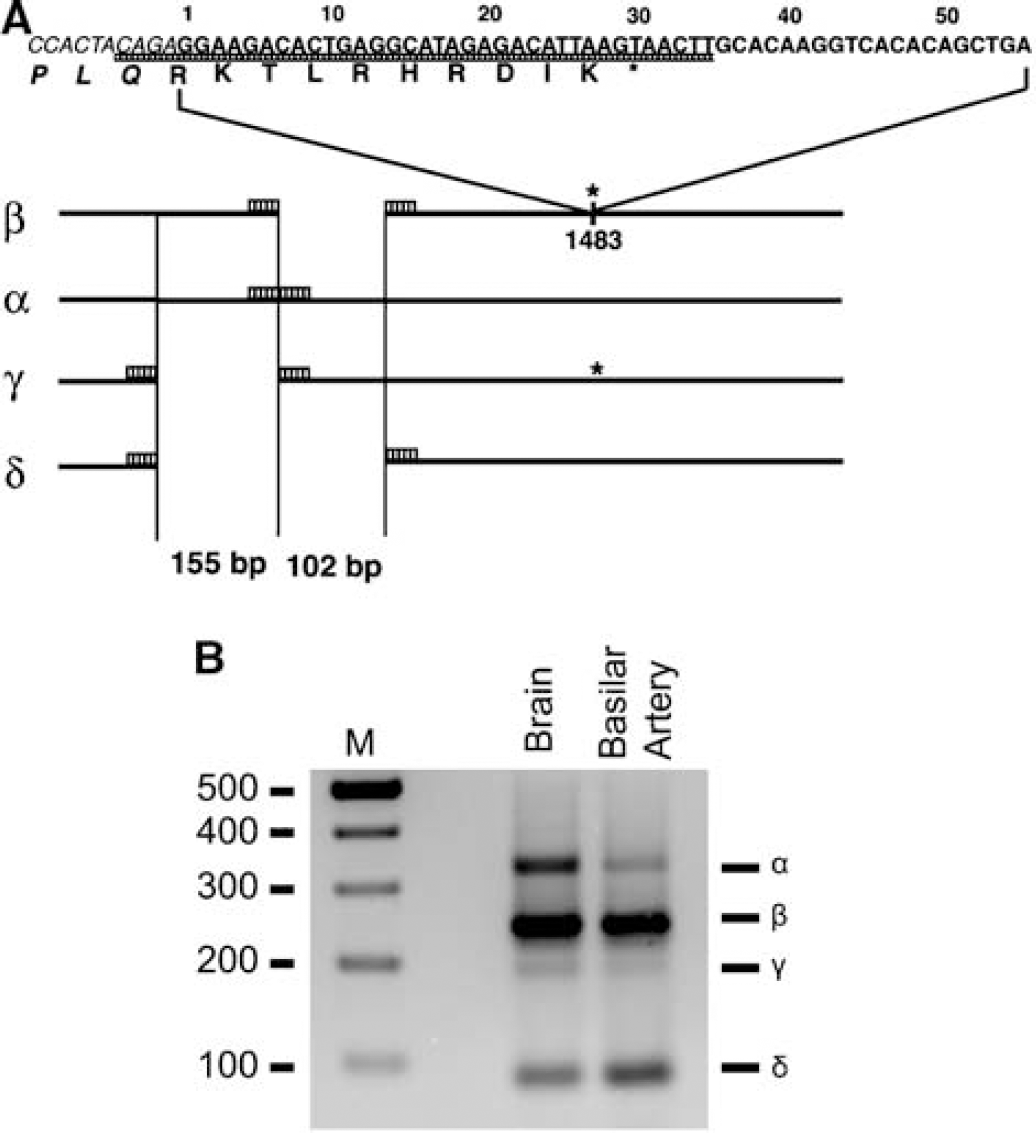
(
(
To explore further the expression of TRPC1 isoforms, we cloned full-length dog TRPC1β. In addition to full-length TRPC1β that had high homology to other mammalian species, we also detected a variant of TRPC1β (TRPC1β55) that had a 55 base pair insertion at amino acid 518. This 55 base insertion produced a termination codon at amino acid 521, producing a smaller protein compared with TRPC1β. Analysis of the TRPC1β hydrophobicity plot, suggests that TRPC1β55 lacks the last three transmembrane domains and the pore region located between fifth and sixth transmembrane domains. The primers used for real-time PCR would not distinguish these two isoforms and the antibody for TRPC1 was directed downstream at a portion of protein, which would be missing in TRPC1β55. We therefore designed new primers to detect TRPC1β55. Only TRPC1β55 was increased (threefold) after SAH without a change in the amount of its longer product (Figure 10A). The total amount of mRNA increase also was appropriate for the overall increase detected before (Figure 9A, TRPC1α and TRPC1β). To determine if this mRNA was translated, we obtained an antibody, which recognized the N-terminal region of TRPC1 (gift of L Tsiokas, University of Oklahoma Health Science Center, OK, USA) and performed Western blot analysis (Figure 10B). Molecular weight of TRPC1β55 is expected to be 58 kDa. Bands of this size were seen in basilar artery 7 days after SAH and were markedly increased compared with control artery. Finally, existence of TRPC1β55 also was suggested by translated product detected in vitro in a rabbit reticulocyte lysate assay (Figure 10C).
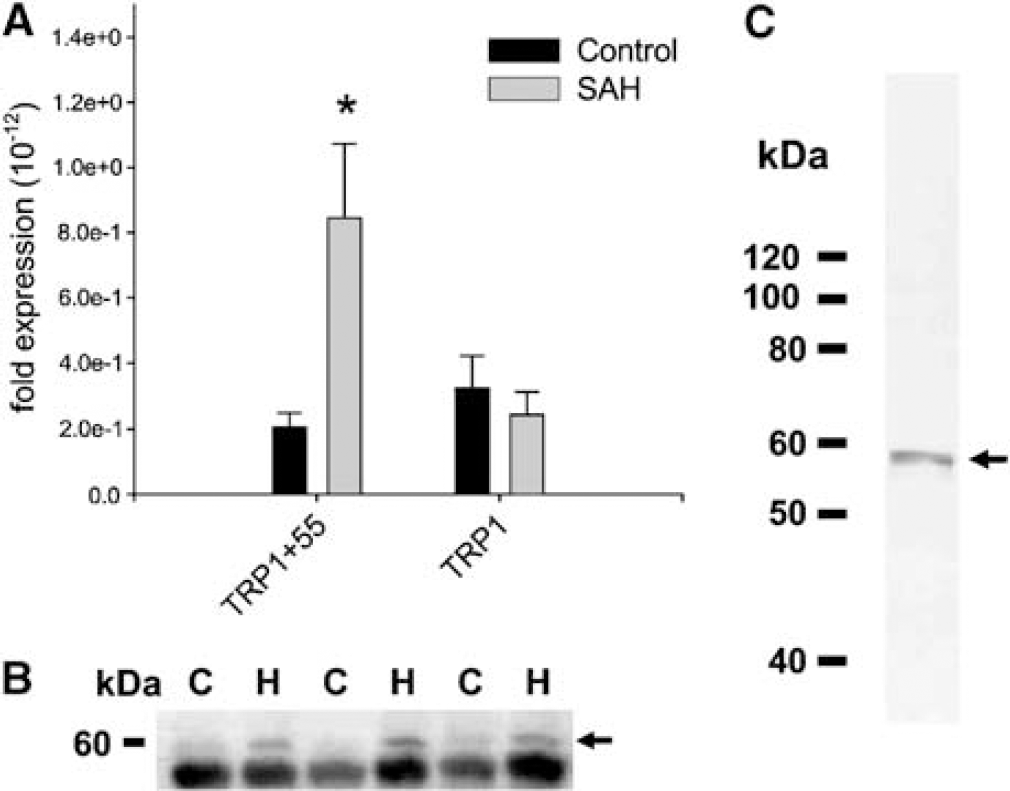
Western blot analysis of TRPC1β55 expression in the dog basilar artery showing summary data (
Discussion
Endothelin-1 is a 21 amino-acid peptide, which is one of the most potent endogenous vasoconstricting agents. Shortly after its discovery, it was implicated in the pathogenesis of cerebral vasospasm (Asano et al, 1989). A review of numerous studies, however, noted that although there was some evidence that ET was increased in cerebrospinal fluid and vasospastic arteries after SAH, concentrations tended to be in the picomolar to low nanomolar range, which would be insufficient to cause the severe constriction often seen during vasospasm (Juvela, 2000). This was contrasted with reports from our and other laboratories that ET receptor antagonists were almost without exception efficacious against vasospasm in animal models (Hino et al, 1995). Furthermore, effectiveness of the ETA receptor antagonist, clazosentan, has been demonstrated in humans after SAH (Vajkoczy et al, 2005; Macdonald et al, 2006). To explain this inconsistency and to determine the molecular mechanism of ET-1-mediated vasospasm after SAH, we studied effects of ET on normal and vasospastic dog basilar artery smooth muscle.
The present results support a mechanism of vasospasm after SAH that involves an increase in smooth muscle TRPC1 and 4 proteins through unknown mechanisms, likely involving phenotypic modulation of smooth muscle (Kumar et al, 2006). These mediate increased sensitivity to ET-1 that contributes to vasospasm. In support of this, vasospastic arteries were more sensitive to ET-1 than normal arteries. The increased sensitivity was antagonized by an antibody to TRPC1 but not to TRPC5. SKF96365, which is used to antagonize store-operated Ca2+ entry, also reduced contractions to ET-1 (Iwamuro et al, 1999). SKF96365, however, also antagonizes voltage-gated Ca2+ entry and cannot be used to differentiate antagonism of store or receptor-operated cation influx or specific currents produced by different heterologously expressed TRP proteins (Iwamuro et al, 1999; Shalabi et al, 2004; Lin et al, 2004). We excluded involvement of voltage-gated Ca2+ channels because dihydropyridine antagonists such as nimodipine did not reduce ET-1-evoked currents. In isolated vasospastic smooth muscle cells, there was increased nonselective cation current, owing to Ca2+ influx, that was also antagonized by anti-TRPC1 and TRPC4 antibodies as well as SKF96365, further supporting a role for TRPC1 and 4. Finally, we found upregulation of a splice variant of TRPC1 and of TRPC4 in vasospastic arteries, which is consistent with the electrophysiologic and isometric tension results.
Further support for this mechanism would be altering vasospasm by antagonism of TRPC isoforms in vivo in the basilar arteries of dogs. The pharmacologic agents that alter nonselective cation channels and their putative mediators, TRP proteins, including SKF96365, LOE908, pyrazole derivatives, and peptide toxins are not very specific (Iwamuro et al, 1999; Shalabi et al, 2004; Lin et al, 2004). Ni2+ has been used but affects numerous ion channels (Beedle et al, 2002). Therefore, it is likely that molecular methods will be needed. Another consideration is that electrophysiology experiments were conducted on isolated cells without pressurization or stretch. This may affect resting membrane potential. We previously measured membrane potential in isolated cells and pressurized arteries from control dogs and those with SAH and SAH cells are depolarized (Weyer et al, 2006). Currents recorded here were not voltage gated but were dependent on membrane potential. As membrane potential in vivo is difficult to know, although the increased sensitivity to ET-1 that was seen is not in question, the magnitude of the effect is uncertain.
Evidence for involvement of TRPC proteins rested in part on use of antibodies to these proteins. The antibody approach is widely used (Morel et al, 2003; Bergdahl et al, 2003; Shalabi et al, 2004; Xu et al, 2006; Kumar et al, 2006). How ET-1 activates the TRPC1/4-mediated Ca2+ influx is unclear. Interactions of TRPC isoforms are complex and activation mechanisms controversial. Transient receptor potential C1 is thought to form a component of the store-operated channel in arterial smooth muscle but it may have other activation mechanisms not dependent on store depletion (Beech, 2005). Blocking TRPC1 inhibited ET-1 contraction of rat caudal artery but not basilar artery unless the basilar artery was organ cultured, a condition associated with increased store-operated Ca2+ entry (Bergdahl et al, 2003; Bergdahl et al, 2005). Transient receptor potential C1 also is upregulated when arterial smooth muscle is stimulated to proliferate (Lin et al, 2004). This study found that the increase in TRPC1 after SAH was not due to an increase in full-length TRPC1 but to a truncated isoform TRPC1β55 that lacks the last three putative transmembrane domains and the pore region. This splice variant resembles that described by Yang et al (2002), in human myometrium and was postulated to disrupt capacitative Ca2+ channel function if translated. We show that this splice variant is potentially translated. It is possible that it disrupts TRPC1/4 hetermultimers, leading to the enhanced ET-1 effects observed here.
Vasospasm and the alteration in TRPC1 was associated with upregulation of TRPC4. Transient receptor potential C4 alone or in heteromultimers with TRPC1 and TRPC5, has been shown to form receptor-operated Ca2+-permeable cation channels (Plant and Schaefer, 2003). Other studies suggest that it forms store-operated Ca2+ channels. This latter role is particularly evident in endothelial cells where it is involved in endothelium-dependent vasodilation, but these cells were not studied here and the endothelium was removed in the molecular biology studies (Freichel et al, 2001). Transient receptor potential C1 might be involved in forming receptor-operated channel complexes with TRPC4 and both may be needed for membrane localization (Hofmann et al, 2002; Ma et al, 2003). The interactions of TRPC1 and 4 might account for their simultaneous increase after SAH in this study and the blocking effects of antibodies to each.
In conclusion, ET-1 significantly increases Ca2+ influx into smooth muscle cells during vasospasm after SAH and this may be mediated by TRPC1 and 4 or their heteromultimers. Thus, manipulation of these TRPC isoforms may be a novel method for inhibiting vasospasm after SAH.