
Abstract
Although previous Northern blot and in situ hybridization studies suggested that neurons express the monocarboxylate transporter MCT2, subsequent immunohistochemical analyzes either failed to confirm the presence of this transporter or revealed only a low density of immunolabeled neuronal processes in vivo. The authors report that appropriate section pretreatment (brief warming episode or proteinase K exposure) leads to extensive labeling of the neuropil, which appears as tiny puncta throughout the whole mouse brain. In addition, intense MCT2 immunoreactivity was found in cerebellar Purkinje cell bodies and their processes, on mossy fibers in the cerebellum, and on sensory fibers in the brainstem. Double immunofluorescent labeling with appropriate markers and observation with epifluorescence and confocal microscopy did not show extensive colocalization of MCT2 immunoreactivity with presynaptic or postsynaptic elements, but colocalization could be observed occasionally in the cortex with the postsynaptic density protein PSD95. Observations made at the electron microscopic level in the cortex corroborated these results and showed that MCT2 immunoreactivity was associated with wide membrane segments of neuronal processes. These data provide convincing evidence that MCT2 represents a major neuronal monocarboxylate transporter in the adult mouse brain, and further suggest that mature neurons could use monocarboxylates such as lactate as additional energy substrates.
Monocarboxylate transporters constitute a family of proton-linked transporters carrying monocarboxylates (i.e., pyruvate and lactate) and ketone bodies (i.e., acetoacetate and β-hydroxybutyrate) as substrates (Halestrap and Price, 1999). Nine putative monocarboxylate transporters (MCT1–MCT9) have been identified on the basis of sequence homologies (Halestrap and Price, 1999), but only four transporters (MCT1–MCT4) have been functionally characterized and proven to transport monocarboxylates (Garcia et al., 1994, 1995; Manning Fox et al., 2000; Yoon et al., 1997). For most MCTs, tissue distribution at least at the mRNA level has been established (Garcia et al., 1994, 1995; Kim et al., 1992; Philp et al., 1998; Price et al., 1998; Wilson et al., 1998). In the central nervous system, both MCT1 and MCT2 mRNAs are highly expressed in the neocortex, hippocampus, and cerebellum, with clear indications of expression in some specific neuronal populations (Koehler-Stec et al., 1998; Pellerin et al., 1998). Moreover, Northern blot analysis on primary cultures of mouse cortical astrocytes and neurons indicated a prominent expression of MCT2 mRNA in neuronal cultures, whereas MCT1 mRNA was particularly enriched in astrocytes (Bröer et al., 1997).
Surprisingly, immunohistochemical studies at the light and electron microscopic levels in rodents have reported a particularly low expression of MCT1 and MCT2 proteins in the brain parenchyma (Gerhart et al., 1997, 1998; Hanu et al., 2000). In the rat brain, MCT immunolabelings were associated with microvessels and glia limitans. MCT1 was expressed by endothelial cells and pericytes (Gerhart et al., 1997), whereas MCT2 expression was restricted to astrocytic end-feet processes (Gerhart et al., 1998; Hanu et al., 2000). Although much less prominent, expression of MCT1 and MCT2 was found in the neuropil (Gerhart et al., 1997, 1998; Hanu et al., 2000; Leino et al., 1999), but the exact nature of the labeled neuropil elements had not been determined. Similar findings were reported for the mouse brain, in which the most intense labeling for MCTs was observed in glia-limiting membranes, ependymocytes, and microvessellike structures, whereas other, less-intensely labeled elements also appeared throughout the brain parenchyma (Pierre et al., 2000). Double immunofluorescent labeling and confocal microscopy examination revealed that the MCT1-positive elements in the neuropil were associated with astrocytic processes, whereas MCT2 immunoreactivity was restricted to neuronal processes (Pierre et al., 2000). Moreover, in a recent study performed in the rat cerebellum, colocalization of MCT2 with glutamate receptors was observed in parallel fiber–Purkinje cell synapses (Bergersen et al., 2001). Nevertheless, the extent of expression of MCTs, and particularly of MCT2 by neurons, remained far below expectations on the basis of mRNA expression. Although such discrepancies had been attributed to methodologic aspects (e.g., alterations of the epitopes located at the C-terminal position) or to control of MCT expression exerted at the translational level, this question remains unresolved (Pierre et al., 2000).
We sought to improve the method for revealing MCT2 immunoreactivity on neurons in adult mouse brain sections. Our approach was based on a procedure that was applied previously to reveal immunoreactivity for neuronal membrane proteins (Watanabe et al., 1998). Using this improved method, we found that MCT2 is widely expressed in the neuropil throughout the whole mouse brain and immunoreactivity is indeed associated with numerous neuronal processes, as expected from mRNA distribution.
MATERIALS AND METHODS
Antibodies
The antibodies used in this study are listed in Table 1. All antibodies were diluted in phosphate-buffered saline containing 0.25% bovine serum.
Antibodies used in this study
MCT, monocarboxylate transporter; MAP2, microtubule-associated protein 2; NF, neurofilament; GFAP, glial fibrillary acidic protein; α-CamKII, α-calmodulin kinase II; PSD, postsynaptic density protein; GluR2, glutamate receptor. More complete manufacturing information is as follows: Sigma, Buchs, Switzerland; Roche Diagnostics, Rotkreuz, Switzerland; Affinity Bioreagents, Alexis Corporation, Lausen, Switzerland; BD Pharmingen, BD Biosciences, Heidelberg, Germany.
Tissues
Tissues were obtained from adult 3-month-old male and female mice. After 1.5 g/kg urethane anesthesia was administered intraperitoneally, the animals were perfused intracardially with a heparinized solution of saline (25 IU/mL in 0.9% sodium chloride) for 2 minutes, followed by a 15-minute perfusion with a freshly prepared solution of 4% paraformaldehyde in phosphate buffer (0.1 mol/L, pH 7.4) or 4% paraformaldehyde and 0.1% glutaraldehyde. Some brains were removed and postfixed in 4% paraformaldehyde for 2 hours, infiltrated with 20% saccharose for 48 hours, frozen in isopentane at −60°C, and cut with a cryostat to obtain serial 16-μm coronal sections collected on gelatin-coated slides. Other brains were postfixed in 4% paraformaldehyde overnight at 4°C and cut on a vibratome to obtain 75-μm coronal slices, which were collected in phosphate-buffered saline (PBS).
Section pretreatment
Before undergoing conventional immunohistochemical analysis, most sections were prewarmed for 10 minutes in a 37°C humidified chamber and incubated at 37°C for 5 to 20 minutes, either in 10-mmol/L Tris-hydrochloride prewarmed at 37°C and containing 10 μg/mL proteinase K (Roche Diagnostics, Rotkreuz, Switzerland). Sections were then briefly rinsed with cold (4°C) PBS to stop the warming reaction and processed for immunohistochemistry.
Immunohistochemistry
Effect of prewarming and proteinase K pretreatment on MCT2 immunohistochemistry in the piriform cortex
For double immunostaining, frozen sections were incubated for 10 minutes in 0.1% X-100 Triton (in PBS), for 1 hour in casein, and then overnight at 4°C in mixtures of primary antibodies. After rinsing in PBS, sections were incubated in a mixture of goat fluorescein isothiocyanate-conjugated antirabbit immunoglobulins (diluted 1/400) and goat antimouse immunoglobulins conjugated with Texas Red (1/400, Jackson Immunoresearch Laboratories) for 2 hours at room temperature. Sections were examined by epifluorescence or by confocal microscopy (DMIRBE; Leica Microsystems AG, Glattbrugg, Switzerland) with appropriate filters.
RESULTS
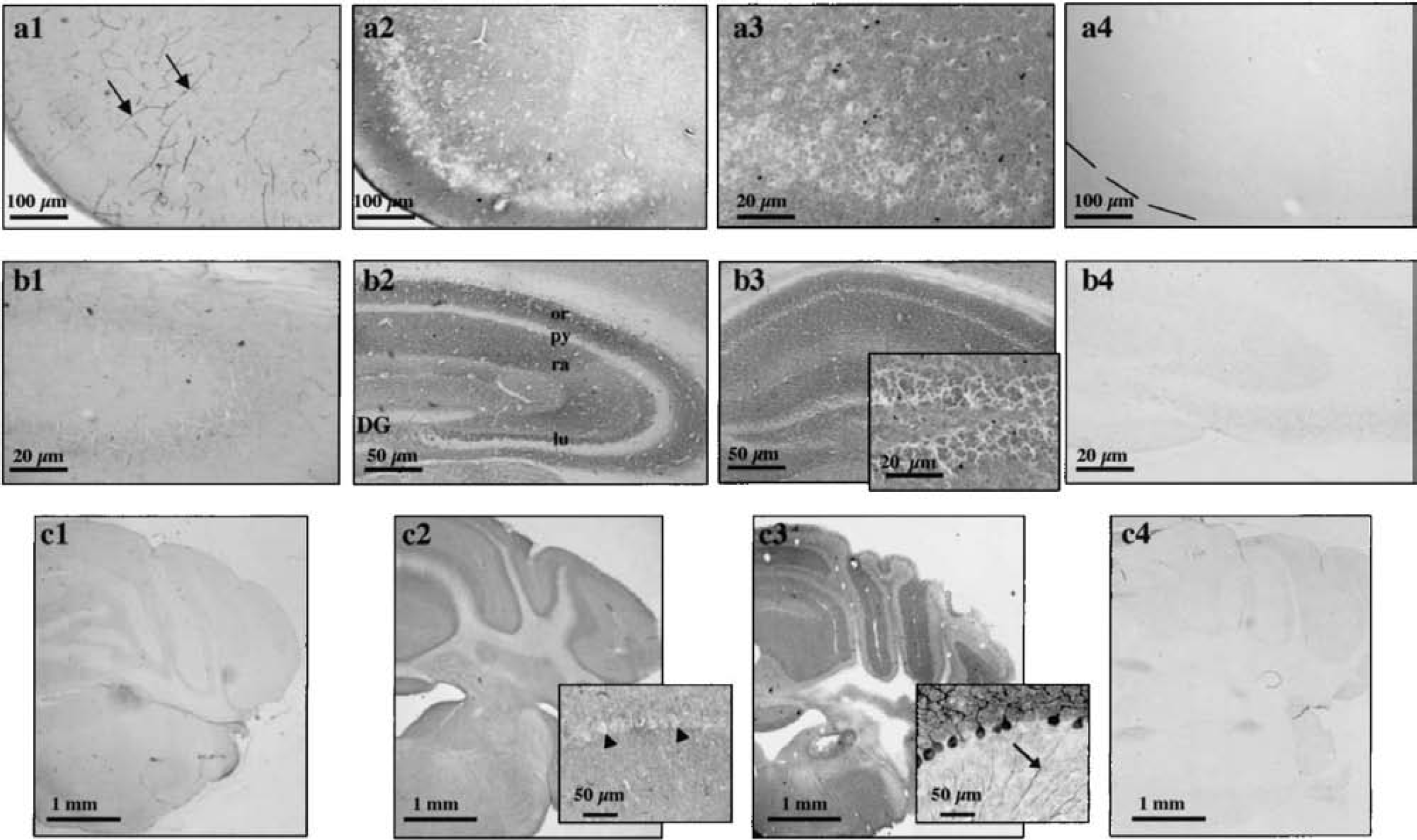
Untreated mouse brain sections displayed MCT2 staining patterns (Figs. 1A1–1C1) comparable with those observed previously (Pierre et al., 2000). Briefly, there was a strong staining at the brain surface corresponding to the location of the glial limiting membrane and in ependymocytes lining the ventricles. This was also visible in several elements resembling small and large blood vessels (Fig. 1A1, arrows). In addition, a small number of labeled cellular processes scattered throughout the brain parenchyma were detectable. To improve immunolabeling, the original immunohistochemical method was modified according to the approach of Fukaya and Watanabe (2000). The guiding principle behind this approach is to expose sections to a protease before performing immunohistochemical analyses to unmask epitopes. Surprisingly, prewarming the sections at 37°C in Tris-hydrochloride buffer without adding a protease was sufficient to dramatically improve immunostaining. Thus, a strong punctate MCT2 labeling appeared in the neuropil in prewarmed coronal sections, and surrounded immunonegative neuronal perikarya (Figs. 1A2, 2, 4A1, 4B1, 6, and 7). As in the untreated sections, labeling of microvessellike elements, choroid plexuses, and ependymocytes lining the ventricles and in glia limitans was still visible (Figs. 4A and 4B). Preabsorption of the anti-MCT2 antibody with its specific peptide antigen prevented the labeling, showing the labeling specificity (Figs. 1A4 and 1B4). In contrast, proteinase K section treatment led only to labeling that was unspecific, visible in all cell nuclei (Figs. 1A3 and 1B3), because it was not affected by preabsorption of the antibody with its specific immunogenic peptide (data not shown). Appearance of nonspecific labeling depended neither on the concentration of the proteinase K solution (10–30 μg/mL) nor on the incubation time with the protease (5–20 minutes).
In the cerebellum, however, prewarming only the sections did not improve staining but enhanced background (Fig. 1C2), and proteinase K pretreatment gave rise to a strong immunoreactivity for MCT2 (Fig. 1C3). Preabsorption of the antibody with its specific antigen prevented the labeling obtained in proteinase K-treated sections (Fig. 1C4). The same approach was tested for MCT1, the other monocarboxylate transporter found in the CNS, which labels several glial processes scattered across the brain parenchyma (Pierre et al., 2000). However, MCT1 immunoreactivity was not improved with the proteinase K pretreatment, which only enhanced unspecific background, or after section warming alone. On these warmed sections, MCT1 immunoreactivity remained restricted to the cellular elements previously described (data not shown) (Pierre et al., 2000).
Distribution of MCT2 immunoreactivity in the mouse brain
To map MCT2 protein expression in the mouse brain, a series of rostrocaudal coronal sections were prewarmed to 37°C in 10-mmol/L Tris-hydrochloride for 10 minutes, whereas sections containing the cerebellum and the brainstem were exposed to proteinase K. Concentration-dependent (5–20 μg/mL) and time-dependent (5–20 minutes) proteinase K pretreatments revealed a bell-shaped effect for both parameters (data not shown). Immunostaining was detectable after 5 minutes of incubation, but the optimal concentration and period of enzyme incubation were 10 μg/mL and 10 minutes, respectively. The following data regarding cerebellar brain sections (Figs. 1C3, 3, 4C1, 5C, 5D, 5E, and 7C) were obtained by incubating the sections with 10-μg/mL proteinase K for 10 minutes at 37°C. The overall distribution of MCT2 in the mouse brain was examined with standard immunoperoxidase and single immunofluorescence. After examination of pretreated serial rostrocaudal coronal sections, it appeared that the MCT2 antibody heavily labeled the neuropil of most brain regions, with high densities in structures like the neocortex, hippocampus, and caudate-putamen (Fig. 2). Neuropil staining was uniform in each of these structures, and there was no delineation related to cortical layers. In the cerebellum, MCT2 immunoreactivity was not apparent in the neuropil, but the cytoplasm of Purkinje neurons perikarya was heavily labeled whereas their dendritic tree in the molecular cell layer was less intensely stained (Figs. 1C3, 3A–3D, 4C1, 5C, and 6C). Their axons were not immunolabeled for MCT2. A strong punctiform MCT2 immunostaining partly underlining the surface of most dendritic processes of the Purkinje neurons was also observed. Although MCT2 did not label other neuronal somata in the granular cell layer, it was expressed on the surface of many mossy fibers (Figs. 1C3, 3A and 3B, 5D, 5E, and 7C) as shown by a double immunofluorescent experiment using an antineurofilament antibody (Figs. 5D and 5E). In the brainstem, there was a strong immunolabeling for MCT2, in particular in sensory fibers such as the trigeminal and facial nerves (Figs. 3E and 3F).

Distribution of MCT2 immunoreactivity in the adult mouse brain. Antibodies raised against MCT2 applied on sections that underwent a warming pretreatment resulted in an intense staining of several structures distributed throughout the mouse brain (top left). Cortical structures [piriform (1), frontal (2) and parietal (3) cortex] were heavily labeled for MCT2. Labeling occurred in the neuropil around negative somata (1, 2, inset, 3). In the hippocampus, cell bodies within the pyramidal cell layer (py, 4) spanning from CA1 through CA3 as well as granule cells of the dentate gyrus (DG, 4) were immunonegative for MCT2. The stratum radiatum (ra, 4, 6) showed a dense MCT2 immunoreactivity whereas the stratum oriens (or, 4, 5, 6) and lucidum (lu, 4, 6) of the hippocampus were more heavily immunolabeled. Immunoperoxidase reaction was viewed with bright-field optics. CA1, CA3, CA1, and CA3, regions of the Ammon horn; mo, stratum moleculare; gr, stratum granulare.
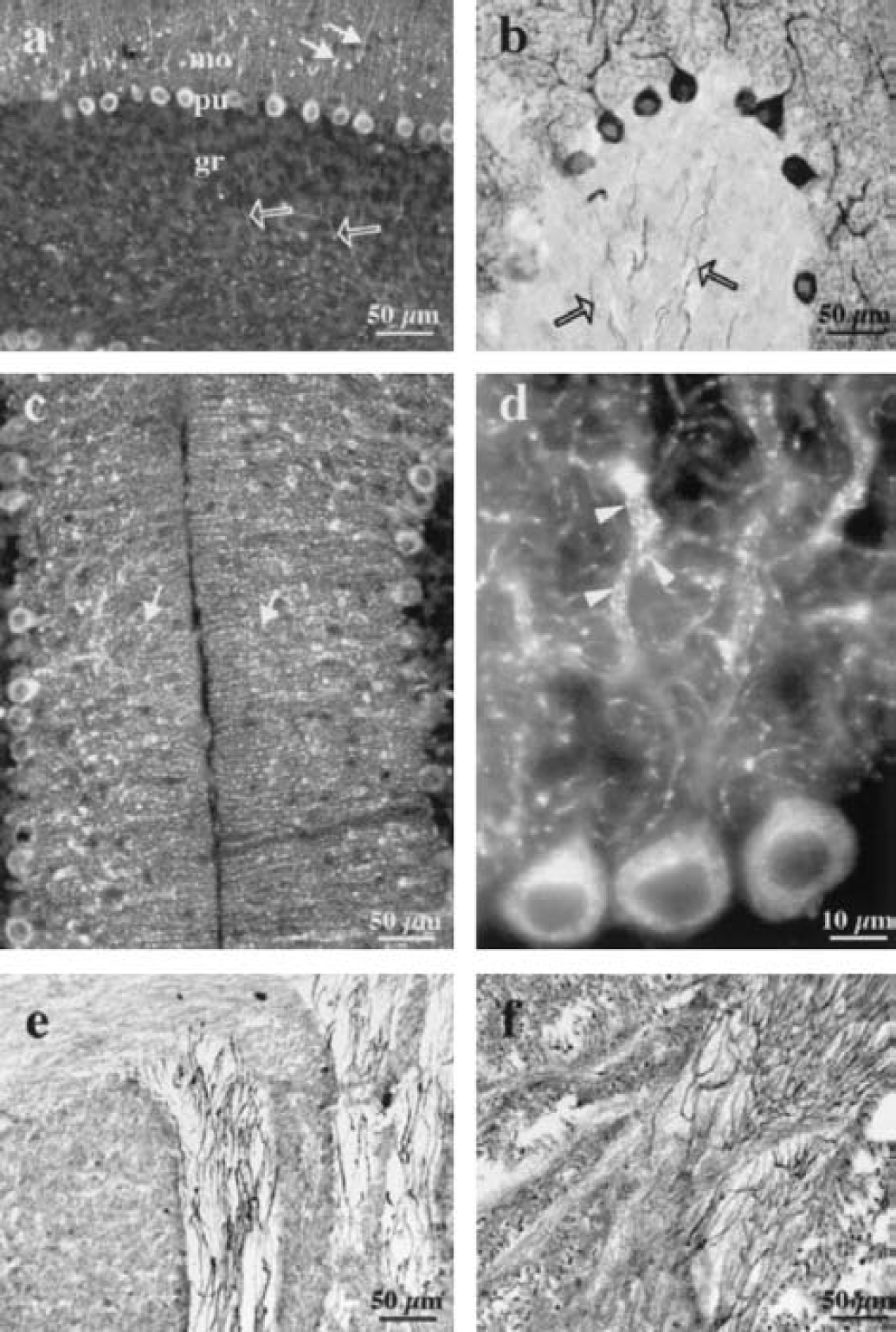
Expression of MCT2 immunoreactivity in the adult mouse cerebellum and brainstem. In the cerebellum, proteinase K pre-treatment of the sections led to an intense MCT2 immunoreactivity of Purkinje neurons observed with fluorescence
Cellular and subcellular localization of MCT2 immunoreactivity
MCT2 labeling occurred predominantly in the neuropil of most structures including the neocortex, caudate-putamen, and hippocampus. In addition, labeling was clearly visible in glia limitans, ependymal cells, and microvessellike structures (Figs. 4A1 and 4B1). To investigate whether MCT2-positive elements in the neuropil were associated with glial or neuronal cell processes, we performed double immunofluorescent labeling on mouse brain sections with either GFAP or neuronal markers (MAP2, mix of neurofilaments 68 and 200) using epifluorescence and confocal microscopy. MCT2 labeling did not colocalize with GFAP immunoreactivity in the neuropil whereas it did colocalize with glia limitans (Fig. 4). Although some of the staining appear to delineate microvessellike structures and could be associated with astrocytic end-feet, these elements only appose GFAP-positive processes and do not colocalize (Pierre et al., 2000). By contrast, a few MCT2-positive elements around MCT2 immunonegative perikarya colocalized with MAP2-positive processes (Figs. 5A1 and 5B3). This observation suggests that the punctiform labeling in the neuropil obtained with MCT2 may be associated with neuronal processes. At high magnification on confocal projections, many MCT2-positive puncta were localized on the surface of these neuronal processes (Fig. 5A3).
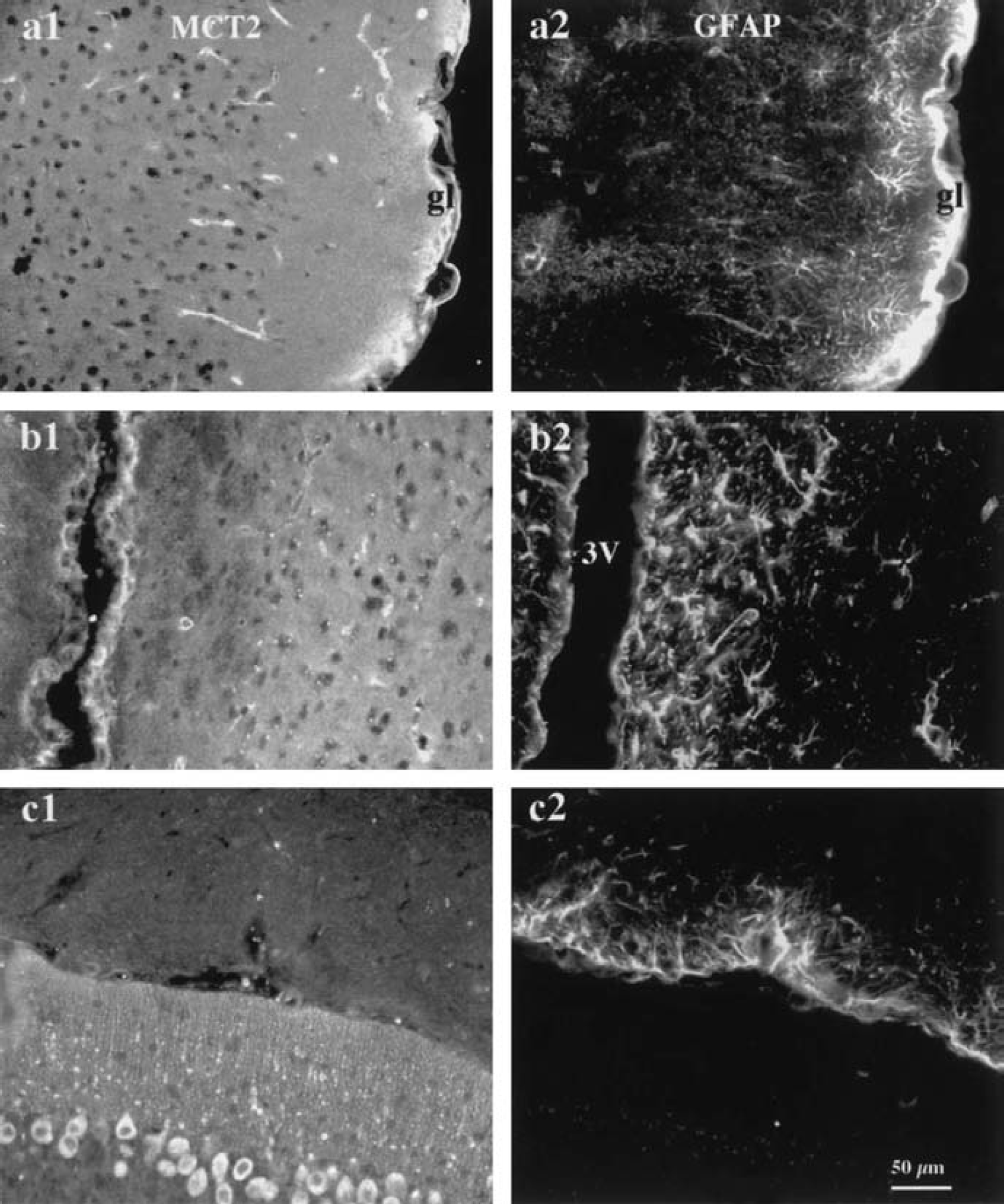
Double immunofluorescence labeling with antibodies against MCT2 and GFAP in the mouse brain. Double immunofluorescent labeling for MCT2
To determine whether MCT2 was present at the synaptic level, we performed double-staining procedures using appropriate markers. We did not observe any colocalization with MCT2 immunoreactivity using synaptophysin as a presynaptic marker (Figs. 6A1–6A3). Then, immunoreactivity for the ionotropic glutamate receptor GluR2, which is present exclusively on postsynaptic terminals, did not colocalize with MCT2 immunoreactivity in any of the regions studied (Figs. 6B1–6C3). Similar negative results were obtained when comparing MCT2 immunoreactivity with immunolabeling for α-CamKinase II (known to complex with proteins containing PDZ domains normally found in postsynaptic densities, data not shown). In contrast, yellow puncta resulting from the colocalization of the two antigen immunoreactivities were detected in the cortex when an antibody raised against PSD95, a postsynaptic protein containing a PDZ domain, was used in conjunction with the MCT2 antibody (Fig. 7A). It is clear, however, that a large majority of MCT2-positive elements were not localized in PSD95 containing synapses and vice versa. We did not observe any colocalization between MCT2 and PSD95 immunoreactivities in brain areas other than the cortex (e.g., in the striatum or in the cerebellum) (Figs. 7B and 7C). To ascertain the exact nature of the elements immunolabeled for MCT2 in the neuropil, we performed electron microscopy using immunoperoxidase-preembedded sections of mouse cortex. At the ultrastructural level, a strong reaction product of immunoperoxidase for MCT2 was clearly visible and covered a large portion of the surface of neuronal processes (Fig. 8). Intracytoplasmic immunoreactivity was not detected in neuronal somata or processes or in glial lamellae.
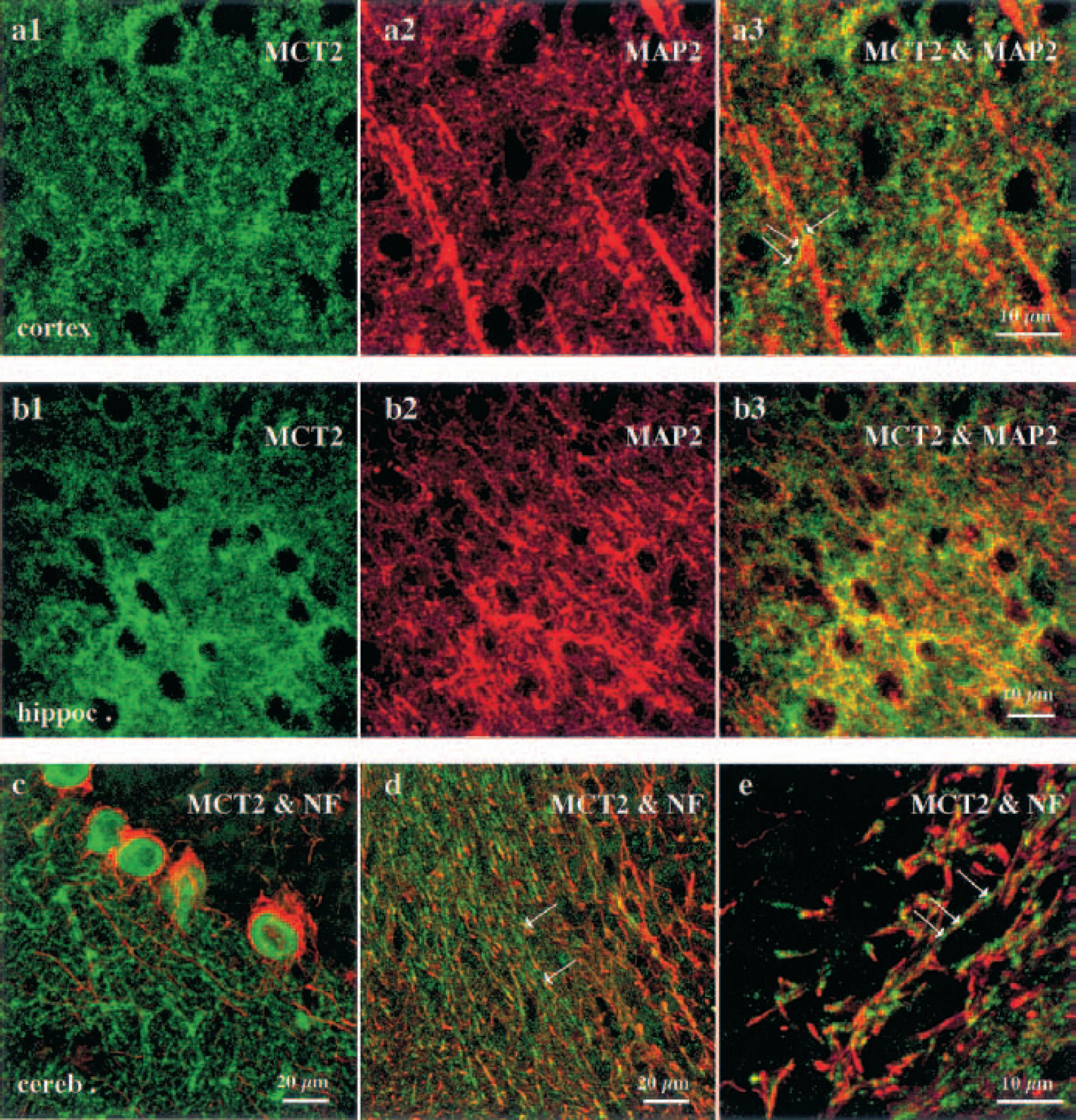
Double immunofluorescence labeling with antibodies against MCT2 and either MAP2 or neurofilament in the mouse brain. MCT2 was revealed with a fluorescein isothiocyanate-conjugated (green) antibody; MAP2 and the cocktail of neurofilaments 68 and 200 (NF), two neuronal markers, were visualized with Texas Red-conjugated immunoglobulins. Single optical projections in confocal microscopy shows MCT2 immunoreactivity in the neuropil around immunonegative perikarya in the cortex
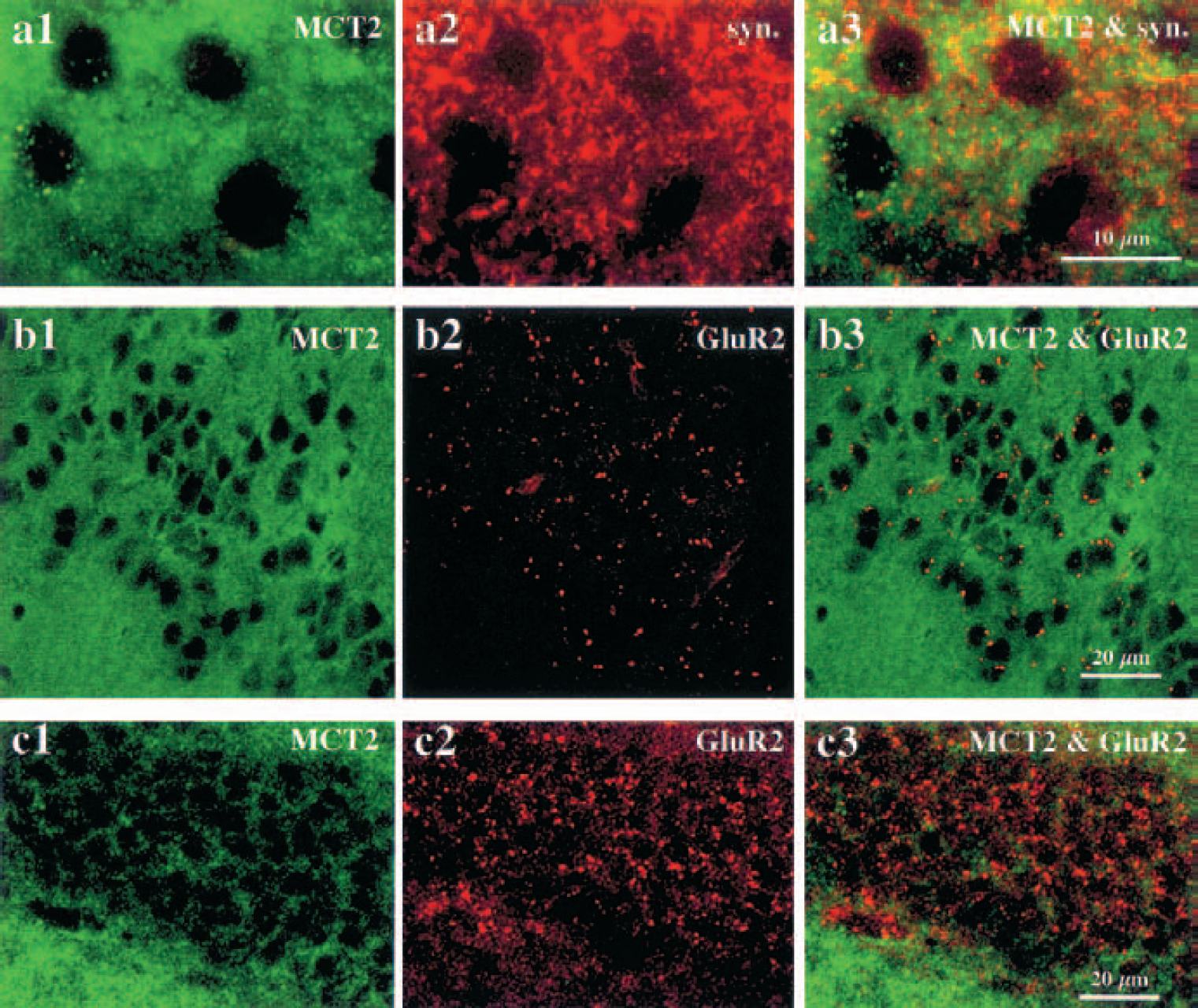
Double immunofluorescence labeling with MCT2 and either synaptophysin or glutamate receptor (GluR2) antibodies in the mouse brain. MCT2 immunolabeling (green) was observed with fluorescein isothiocyanate-conjugated antibody, and immunoreactivities for synaptophysin or GluR2 (red) were revealed with a Texas red-conjugated secondary antibody. The punctiform immunolabeling of MCT2 occurring around immunonegative cortical perikarya
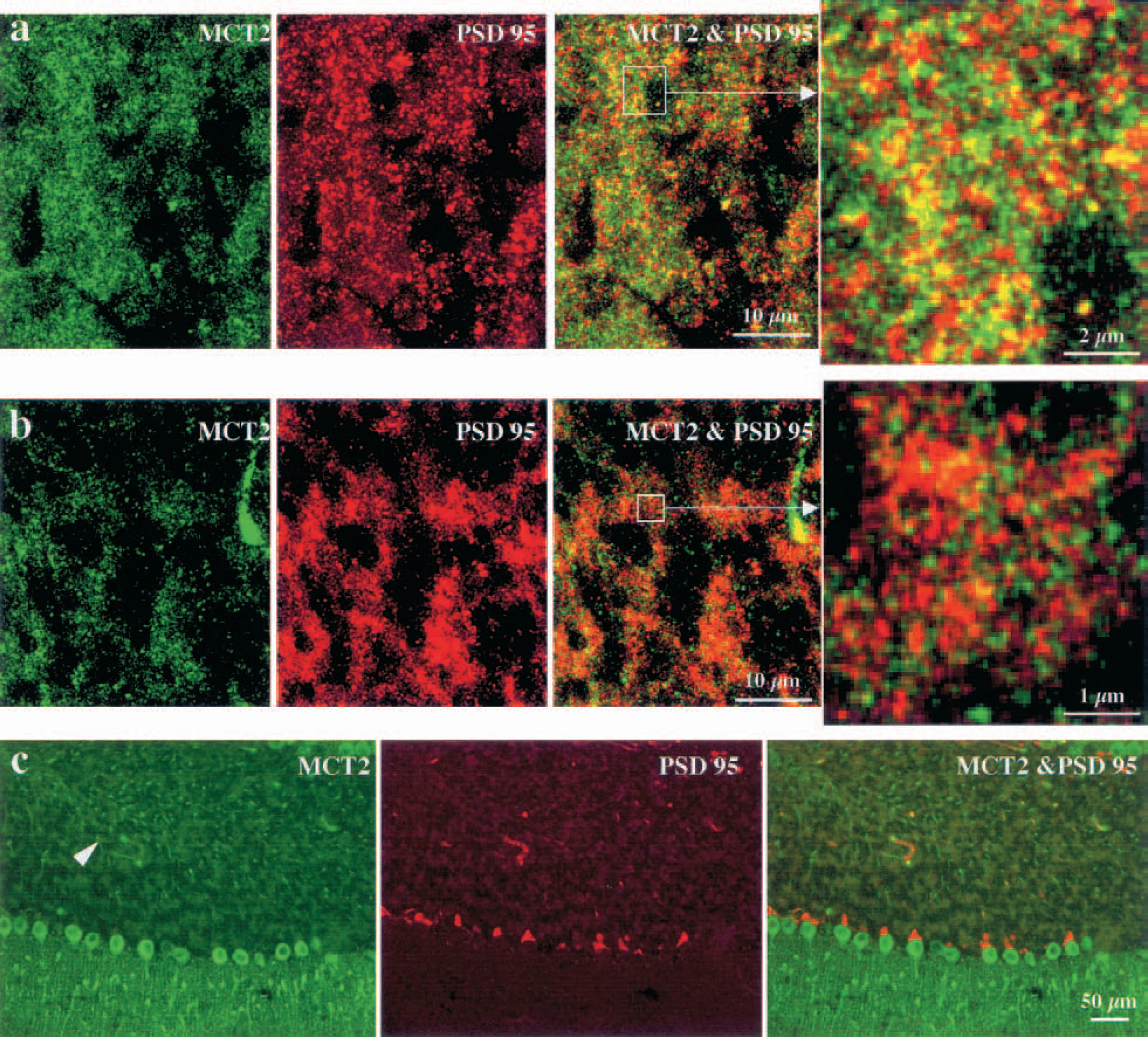
Double immunofluorescence labeling with antibodies against MCT2 and PSD95 in the mouse brain. A certain fraction of MCT2-positive puncta (green) observed with fluorescein isothiocyanate-conjugated immunoglobulins could be found in synaptic elements revealed with the postsynaptic marker PSD95 (red) observed with Texas Red conjugated antibodies in the cortex
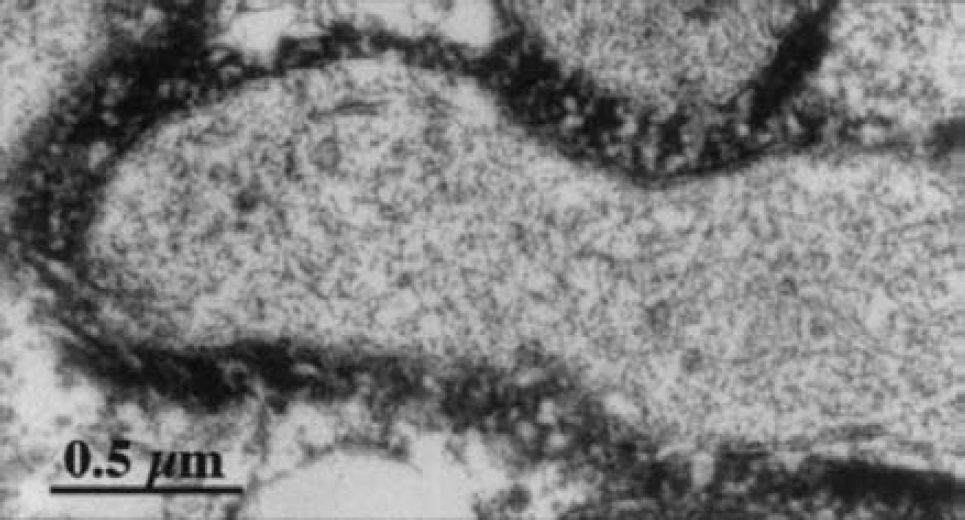
Ultrastructural localization of MCT2 immunoreactivity in the cortex. An intense MCT2 immunoreactivity is associated with the surface of dendrites. There is no immunolabeling in the cytoplasm of these processes. Preembedding immunoperoxidase reaction was observed with electron microscopy.
DISCUSSION
Improved detection of MCT2 immunoreactivity by warming or protease pretreatment of brain sections
MCT2 immunoreactivity was dramatically modified in most brain regions in sections that had been warmed up to 37°C before conventional immunohistochemistry. In view of the present results, the observed expression pattern of the MCT2 protein is now more consistent with its previously described mRNA distribution, which showed specifically high expression in the neocortex and the hippocampus (Koehler-Stec et al., 1998; Pellerin et al., 1998). In the cerebellum (and in the brainstem), however, a proteinase K pretreatment of the sections was necessary to observe an enhanced MCT2 immunostaining. After such treatment, results are in agreement with those obtained by Bergersen et al. (2001), who recently reported intense MCT2 labeling in Purkinje cell bodies and a punctate labeling in the molecular layer in a rat model.
Such a remarkable enhancement in immunohistochemical detection has been reported previously for synaptic proteins, such as the different N-methyl-
Cellular and subcellular distribution of MCT2
In the present study, immunoreactivity for MCT2 was detected in numerous cellular elements of the neuropil distributed throughout the mouse brain parenchyma. Examination with double immunofluorescence labeling and confocal microscopy was performed to identify the cell types bearing MCT2-positive processes. Using epifluorescence or confocal microscopy, we found no colocalization between MCT2 and the classical astrocytic marker GFAP in elements of the neuropil, whereas such colocalization was observed in the glia limitans. Although MCT2 is strongly expressed by astrocytes and by astrocytic end-feet in the rat brain (Gerhart et al., 1998; Hanu et al., 2000), MCT2 was not associated with glial elements in the neuropil and in those processes closely apposed to microvessellike structures in the mouse brain (Pierre et al., 2000). Despite an increased expression of MCT2 after warming or appropriate proteinase K pre-treatment of sections, we did not observe colocalization between MCT2 and GFAP immunoreactivities in the neuropil. This result was corroborated by electron microscopy, which failed to show any MCT2 peroxidase product in any glial soma or process in the neocortex. These data suggest that astrocytes (and astrocytic end-feet), with the exception of glial cells of the glia limitans, do not significantly express MCT2 in this species. However, in accordance with our previous results (Pierre et al., 2000), MAP2-positive elements and processes containing neurofilaments 68 and 200 immunoreactivities showed MCT2 immunolabeling. Thus, MCT2 is clearly a neuronal isoform in the mouse brain.
Examination by confocal microscopy with double fluorescence labeling revealed that MCT2 immunoreactivity was localized on the surface of identified neuronal processes, in contrast to neuronal markers such as MAP2 or neurofilaments, which are cytoskeletal elements. Electron microscopy in the cortex confirmed that MCT2 immunoreactivity did not occur intracytoplasmically in neuronal processes, but was rather strongly expressed on their surface. A striking exception, however, remains in the cerebellum where Purkinje cell bodies were heavily stained. Nevertheless, the strong MCT2 immunoreactivity in the neuropil of most brain regions is thus associated with the plasma membrane of numerous neuronal processes. Attempts to find a colocalization between either presynaptic or postsynaptic markers were unsuccessful. Thus, data strongly suggest that the majority of the MCT2 immunoreactivity is extrasynaptic but remains associated with neuronal membranes, an observation substantiated by electron microscopic findings. In view of such an extensive expression and distribution on neuronal surface, MCT2 can be considered as a major neuronal monocarboxylate transporter throughout the mouse brain.
Functional considerations
The role of monocarboxylates as energy substrates for the developing brain has been well documented (Nehlig and Pereira de Vasconcelos, 1993). In contrast, the role of monocarboxylates, and particularly lactate, as energy substrates in the normal adult brain has remained more controversial (Ames, 2000). Recent evidence obtained by nuclear magnetic resonance both in vitro and in vivo indicates that extracellular lactate is used effectively by neurons (Bouzier et al., 2000; Qu et al., 2000; Waagepetersen et al., 1998), which is possible only if an appropriate transporter is expressed on their surface. As evidenced by our results, the monocarboxylate transporter MCT2 could fulfill this role in the mouse brain.
Footnotes
Acknowledgments:
The authors thank Mr. C. Verdan for technical help with electron microscopy and Dr. M. Di Luca for her initial gift of PSD95 antibody.