
Abstract
The blood–brain barrier (BBB) is formed by the brain microvascular endothelium, and the unique transport properties of the BBB are derived from tissue-specific gene expression within this cell. The current studies developed a gene microarray approach specific for the BBB by purifying the initial mRNA from isolated rat brain capillaries to generate tester cDNA. A polymerase chain reaction–based subtraction cloning method, suppression subtractive hybridization (SSH), was used, and the BBB cDNA was subtracted with driver cDNA produced from mRNA isolated from rat liver and kidney. Screening 5% of the subtracted tester cDNA resulted in identification of 50 gene products and more than 80% of those were selectively expressed at the BBB; these included novel gene sequences not found in existing databases, ESTs, and known genes that were not known to be selectively expressed at the BBB. Genes in the latter category include tissue plasminogen activator, insulin-like growth factor-2, PC-3 gene product, myelin basic protein, regulator of G protein signaling 5, utrophin, IκB, connexin-45, the class I major histocompatibility complex, the rat homologue of the transcription factors hbrm or EZH1, and organic anion transporting polypeptide type 2. Knowledge of tissue-specific gene expression at the BBB could lead to new targets for brain drug delivery and could elucidate mechanisms of brain pathology at the microvascular level.
The transport of solutes and drugs into the brain is regulated by transport systems present at the blood–brain barrier (BBB). The BBB is formed at the luminal and abluminal membranes of the brain capillary endothelium (Brightman et al., 1970). The capillary endothelial cells of the vertebrate brain express epithelial-like high resistance tight junctions that fuse the plasma membranes of neighboring capillary endothelial cells in brain. The tight junctions of the BBB eliminate the porous pathways of solute diffusion across the endothelial cell in brain, which exist in capillary beds perfusing nonbrain organs. Transport systems expressed at the BBB may mediate either the influx of solutes or drugs from blood to brain or the active efflux of solutes or drugs from brain to blood (Pardridge, 1998).
Novel genes encoding BBB transporters can be identified with standard genomic approaches based on gene microarrays. However, the sensitivity of gene microarrays is approximately 10−4 (Schena et al., 1995). The volume of the brain capillary endothelium forming the BBB in vivo is <1 μL/g (Pardridge, 1998), or approximately 10−3 parts of whole brain. Therefore, it is statistically unlikely that genes that are selectively expressed in brain at the microvasculature will be detected in a gene microarray derived from whole brain. Blood–brain barrier–specific genes can be rapidly identified with gene microarrays derived from mRNA obtained from purified brain capillaries. The isolation of BBB-specific genes is aided with the use of subtraction cloning methods, which allow for the identification of genes that are differentially expressed at the BBB relative to peripheral tissues such as liver or kidney (Liang and Pardee, 1992; Diatchenko et al., 1996; Welford et al., 1998). One such approach is suppression subtractive hybridization (SSH) (Diatchenko et al., 1996), and the current investigation describes initial results using SSH to clone genes that are selectively expressed at the BBB.
MATERIALS AND METHODS
Isolation of poly (A)+ RNA
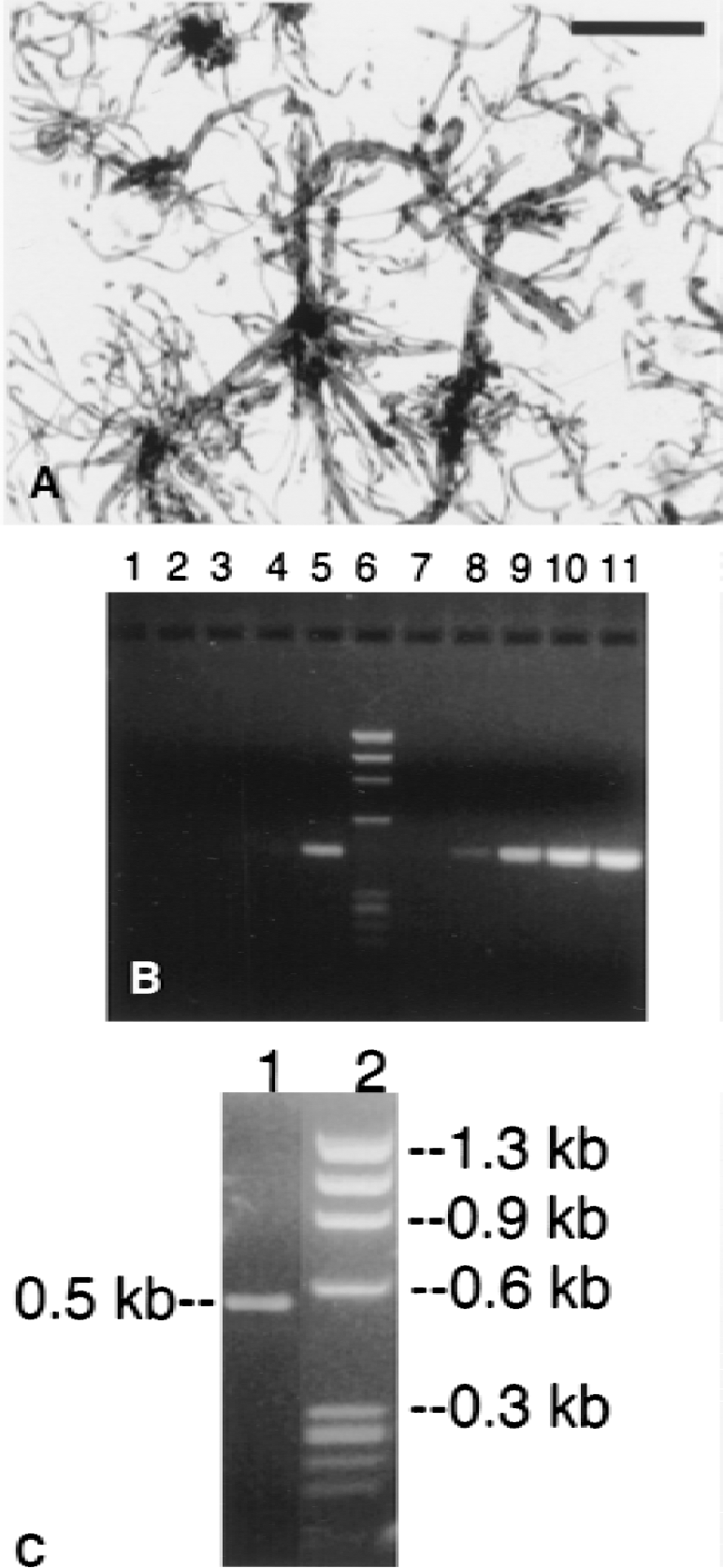
Poly A+ RNA was isolated from rat and bovine brain capillaries, and rat brain and peripheral tissues using a single step method previously described (Boado and Pardridge, 1991). Brain capillaries were free of adjoining brain tissue (Fig. 1A). The yield of poly A+ RNA from bovine and rat brain capillaries was 12 μg per single bovine brain cortical shell and 3.2 μg from the pooled cerebral hemispheres of 21 rat brains, respectively.
Suppression subtractive hybridization
Suppression subtractive hybridization was performed using the PCR-Select cDNA subtraction system according to the manufacturer's instructions (Clontech, Palo Alto, CA, U.S.A.). The rat brain capillary poly A+ RNA was used to produce 2.5 μg of tester cDNA, and the subtraction procedure was completed using either rat liver or kidney mRNA to generate driver cDNA. The reaction was followed with [32P]-dCTP. The tester or driver cDNA was digested with RsaI to obtain shorter, bluntend molecules, and 2 tester populations were created as either adaptor 1 or adaptor 2R and were independently ligated to 350 ng of the tester cDNA. The 2 populations of adaptor-ligated tester cDNA were independently hybridized (at 98°C for 1.5 minutes and 68°C for 8 hours) to the driver cDNA to enrich for differentially expressed sequences, and hybridized a second time (at 68°C for 16 hours) to generate a polymerase chain reaction (PCR) template. A first-run PCR using PCR primer 1 amplifies differentially expressed sequences and was performed for 30 cycles (denaturation, 94°C for 30 seconds; annealing, 66°C for 30 seconds; extension, 72°C 1.5 minutes). A second-run PCR was performed for 15 cycles (denaturation, 94°C for 30 seconds; annealing, 68°C for 30 seconds; extension, 72°C 1.5 minutes), using nested PCR primers 1 and 2R. This second-run PCR further enriches for differentially expressed sequences and suppresses the background (Diatchenko et al., 1996).
Subtracted cDNA screening
The SSH-PCR products were cloned into the pCR2.1 vector and a cDNA library was prepared in E. coli INVαF' cells. Positive clones were identified by differential hybridization. The cDNA library was plated on Luria-Bertani medium/ampicillin (LB/amp) plates, and white colonies were picked and individually grown in 200 μL LB/amp medium in 96-well plates for 24 hours at 37°C. Colonies were individually blotted onto GeneScreen Plus membrane using a 96-well dot-blot system (Bio-Rad, Richmond, CA, U.S.A.). Membranes were hybridized with [32P]-labeled subtracted or unsubtracted rat brain capillary tester cDNA (Wang et al., 1998), and film autoradiography was performed with Biomax MS film (Kodak, Rochester, NY, U.S.A.) for 18 hours at 23°C. cDNA was purified with Qiaquick PCR purification kit (Qiagen, Santa Clarita, CA, U.S.A.) and labeled as previously described (Boado et al., 1999). Clones showing a strong hybridization signal with the subtracted probe compared with the unsubtracted one were selected for DNA sequencing and Northern blot analysis after release of the pCR2.1 insert with EcoRI. Northern blotting was performed as described previously (Boado et al., 1999).
DNA sequence analysis
DNA sequencing of isolated clones was performed in both directions at Biotech Core (Mountain View, CA, U.S.A.) using M13 forward and reverse primers (Boado et al., 1999). Similarities with other genes in GenBank were investigated using the BLAST program (NCBI, NIH). Plasmid DNA was amplified and purified from positive clones using the Qiafilter Maxi Kit (Qiagen, Valencia, CA, U.S.A.).
In situ hybridization
The cDNA fragment obtained from SSH was released from the pCR2.1 vector by double digestion with SalI and XbaI and cloned into the pSPT19 vector at the same restriction endonuclease sites. Antisense and sense RNA probes were generated using SalI and XbaI linearized pSPT19- plasmid and T7 and SP6 polymerases, respectively, and digoxigenin-11-uridine triphosphate (DIG-11-UTP), as part of the DIG RNA Labeling kit (Roche Molecular Biochemicals, Indianapolis, IN, U.S.A.). Freshly isolated rat brain capillaries were cytocentrifuged to glass slides and fixed with 10% formalin before storage at −70°C. The slides were hybridized with 12 ng digoxigenin (DIG)-labeled probes in 30 μL of hybridization solution overnight at 42°C, and control sections were hybridized with an equal amount of sense cRNA according to the manufacturer's instructions.
RESULTS
The cDNA products of the first and second cycle PCR ranged in size from 0.2 to 1.4 kb and the majority of the PCR products had a size ranging from 0.3 to 0.7 kb (Table 1). An initial small library was prepared with only kidney-derived driver cDNA, and three clones were isolated and designated K1-K3 (Table 1). The larger library was generated from rat liver and kidney-derived driver cDNA, and the first 50 clones isolated were designated LK1-LK50 (Table 1). The efficiency of the subtraction procedure was analyzed by PCR amplification of cDNA for G3PDH as shown in Fig. 1B. Using the subtracted tester cDNA, no G3PDH PCR product was identified until 33 cycles of PCR (lane 5, Fig. 1B). Conversely, the G3PDH cDNA was identified in PCR of the unsubtracted tester cDNA as early as 18 cycles (lane 8, Fig. 1B). Products of the secondary PCR were cloned into the pCR 2.1 vector for transformation of INVαF'. Comparison of the subtracted and unsubtracted Southern dot blots of the 96-well microarrays identified several gene products that were enriched at the BBB and the sizes of these inserts were determined by agarose gel electrophoresis after release with EcoRI (Fig. 1C).
Summary of isolated clones
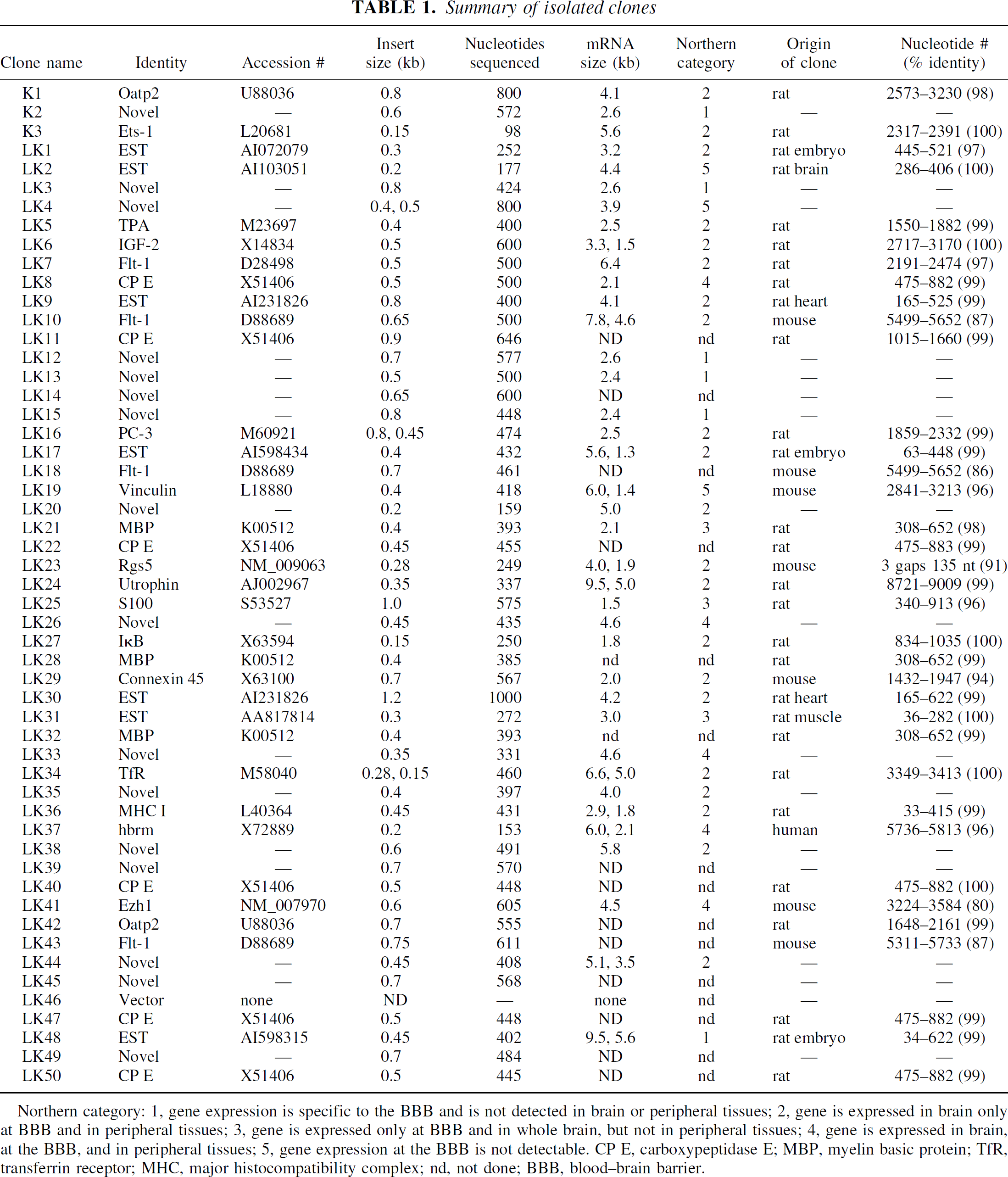
Northern category: 1, gene expression is specific to the BBB and is not detected in brain or peripheral tissues; 2, gene is expressed in brain only at BBB and in peripheral tissues; 3, gene is expressed only at BBB and in whole brain, but not in peripheral tissues; 4, gene is expressed in brain, at the BBB, and in peripheral tissues; 5, gene expression at the BBB is not detectable. CP E, carboxypeptidase E; MBP, myelin basic protein; TfR, transferrin receptor; MHC, major histocompatibility complex; nd, not done; BBB, blood–brain barrier.
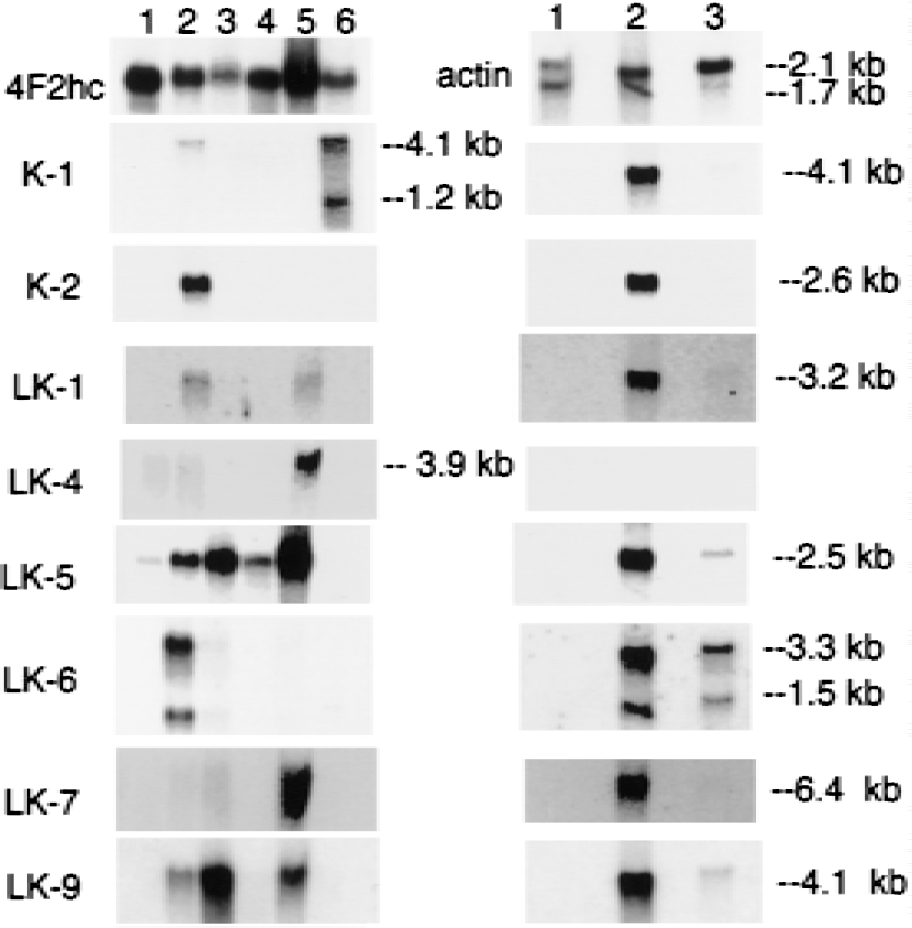
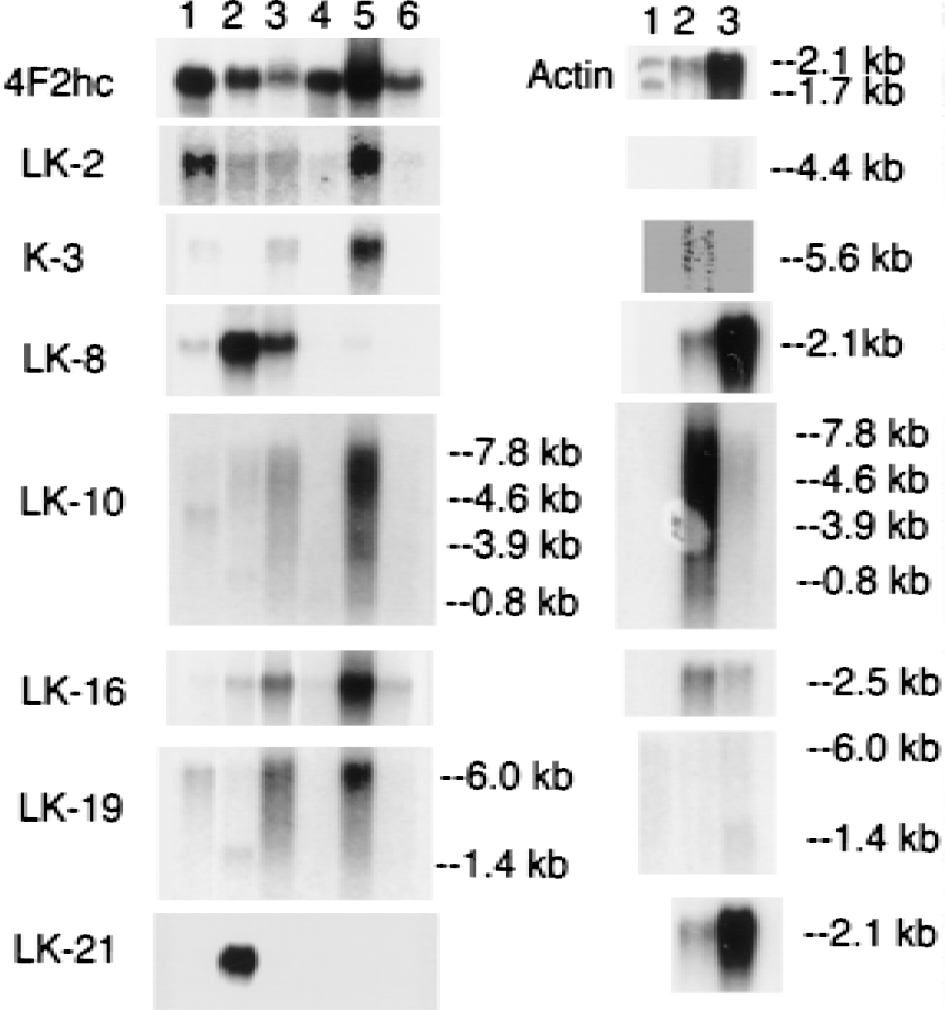
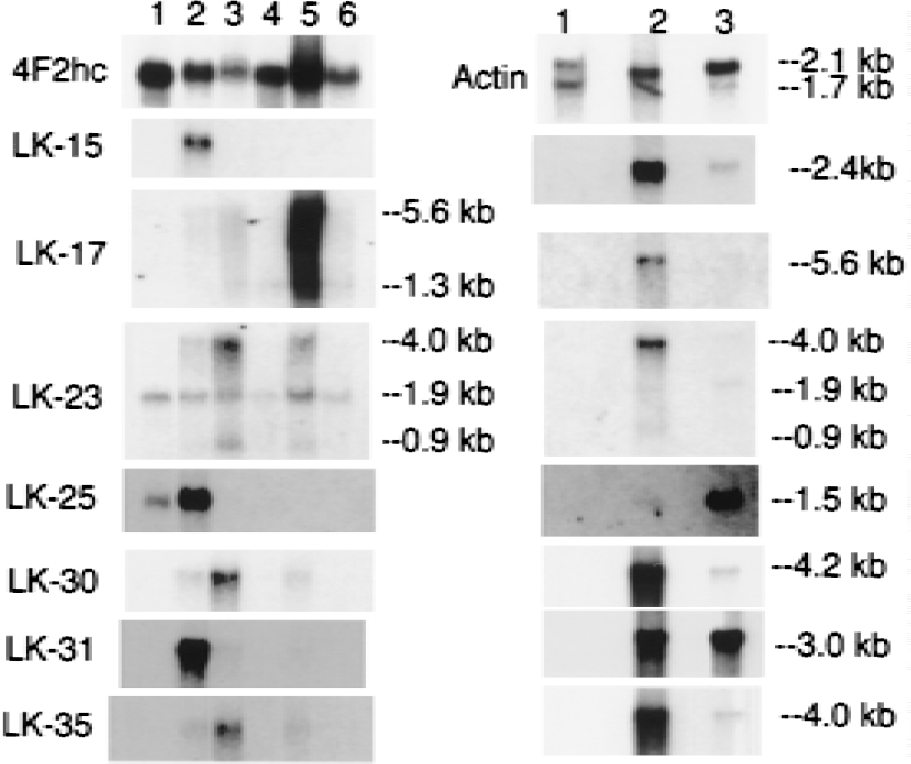
Northern blot analysis was performed on all clones, except where sequence analysis indicated there was overlap with other clones, and the results of the Northern blots are shown in Figs. 2 to 5. Each blot was also probed with cDNA for two commonly expressed genes, actin and 4F2hc, which encode the heavy chain of amino acid transporter heterodimers (Boado et al., 1999). In situ hybridization results for four selected clones are shown in Fig. 6 for antisense and sense RNA. Based on the Northern blotting results, the clones were classified into 1 of 5 categories of differential gene expression, as shown in Fig. 7. Northern blot analysis with clones LK2, a rat EST, LK4, a novel gene, and LK19, vinculin (Table 1) showed the corresponding transcripts were underexpressed at the BBB (Fig. 7).
Northern blotting.
Northern blotting.
Northern blotting.
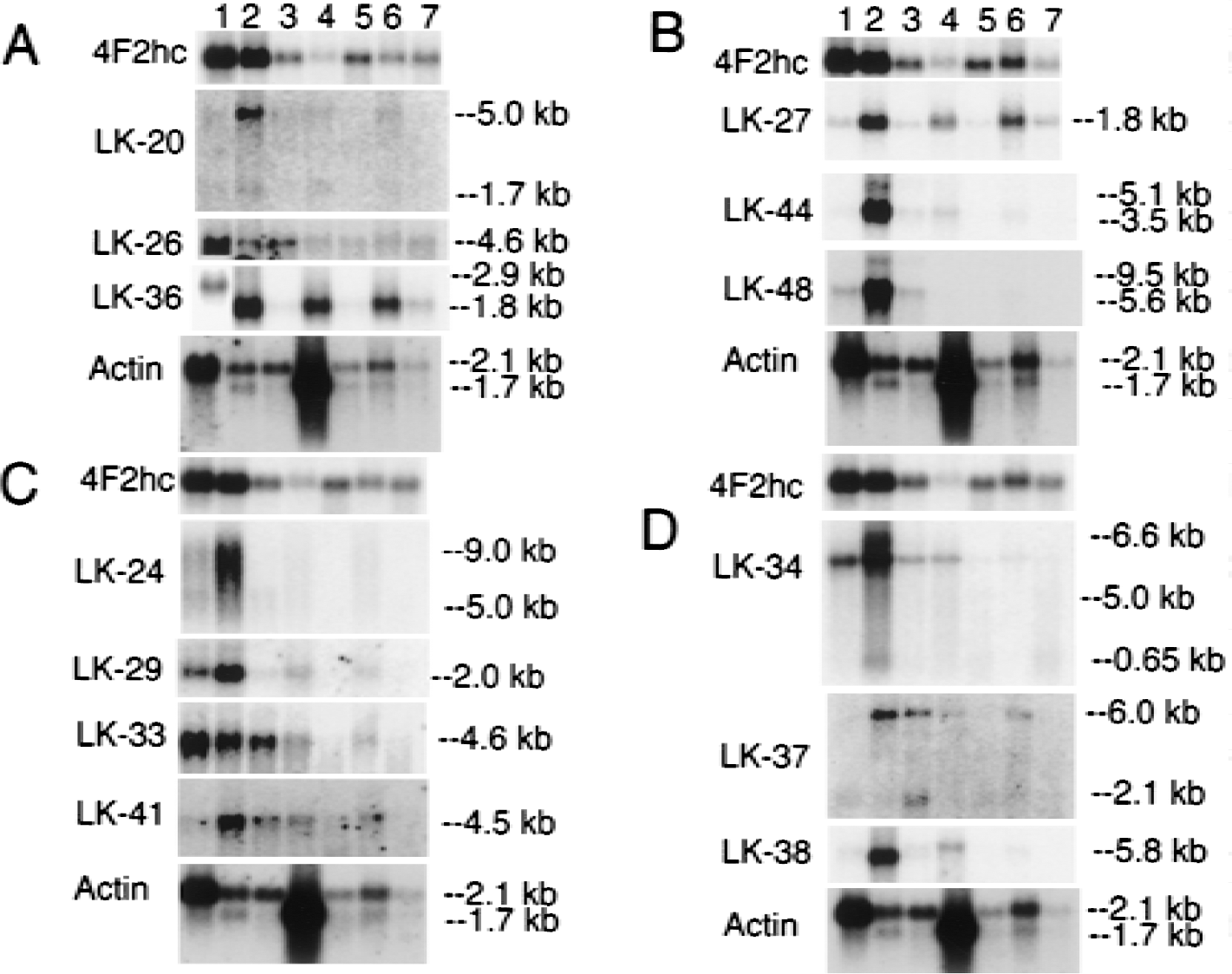
Northern blotting. Lanes 1 to 7: C6 rat glioma cells, rat brain capillaries, total rat brain, rat heart, rat kidney, rat lung, and rat liver, respectively. Film exposure time: 1 day for actin, 4F2hc, LK24, LK27, LK36, LK38, LK44, and LK48; 5 days for LK20, LK26, LK29, LK33, LK34, and LK41; 6 days for LK37. Two micrograms poly A+ RNA was applied to all lanes. Panels
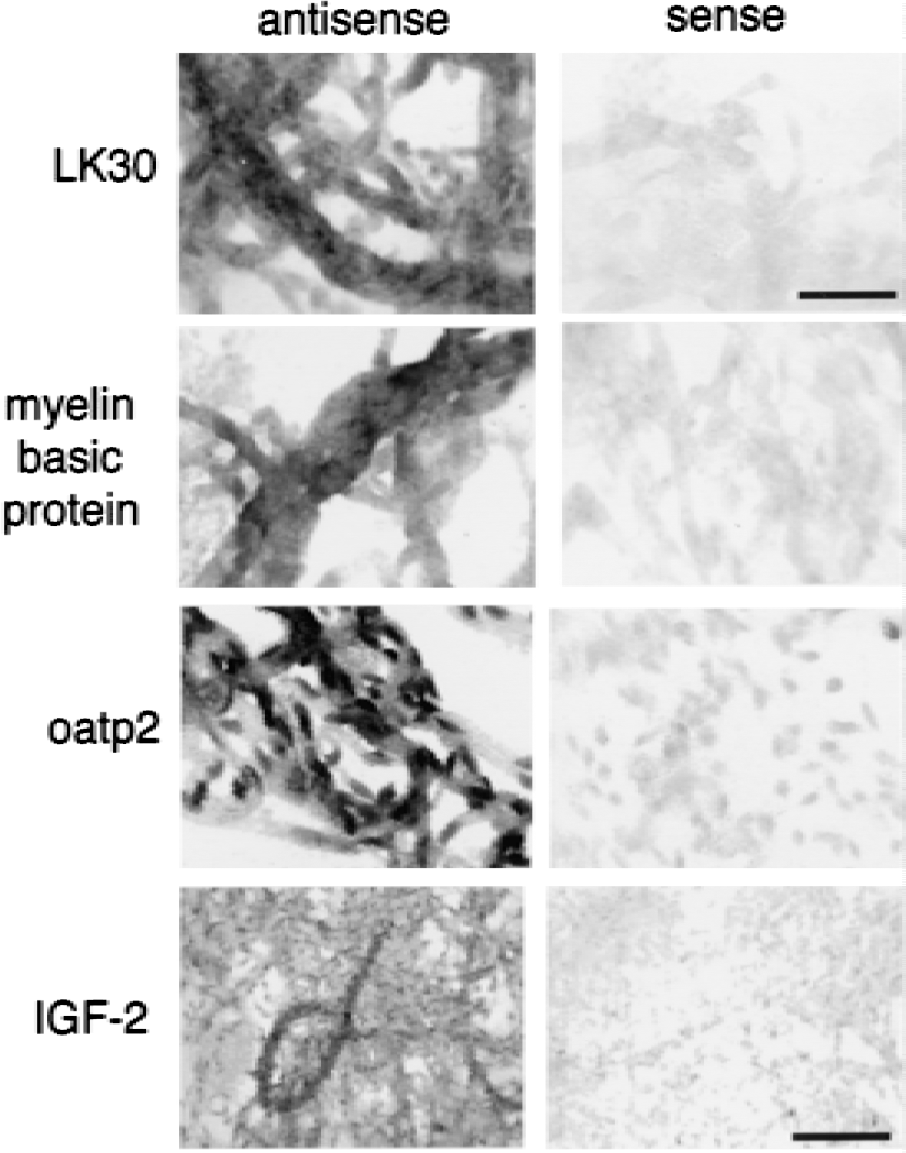
In situ hybridization. Magnification for LK30, myelin basic protein, and oatp2 is the same. Magnification bars = 34 and 85 μm for LK30 and IGF-2, respectively.
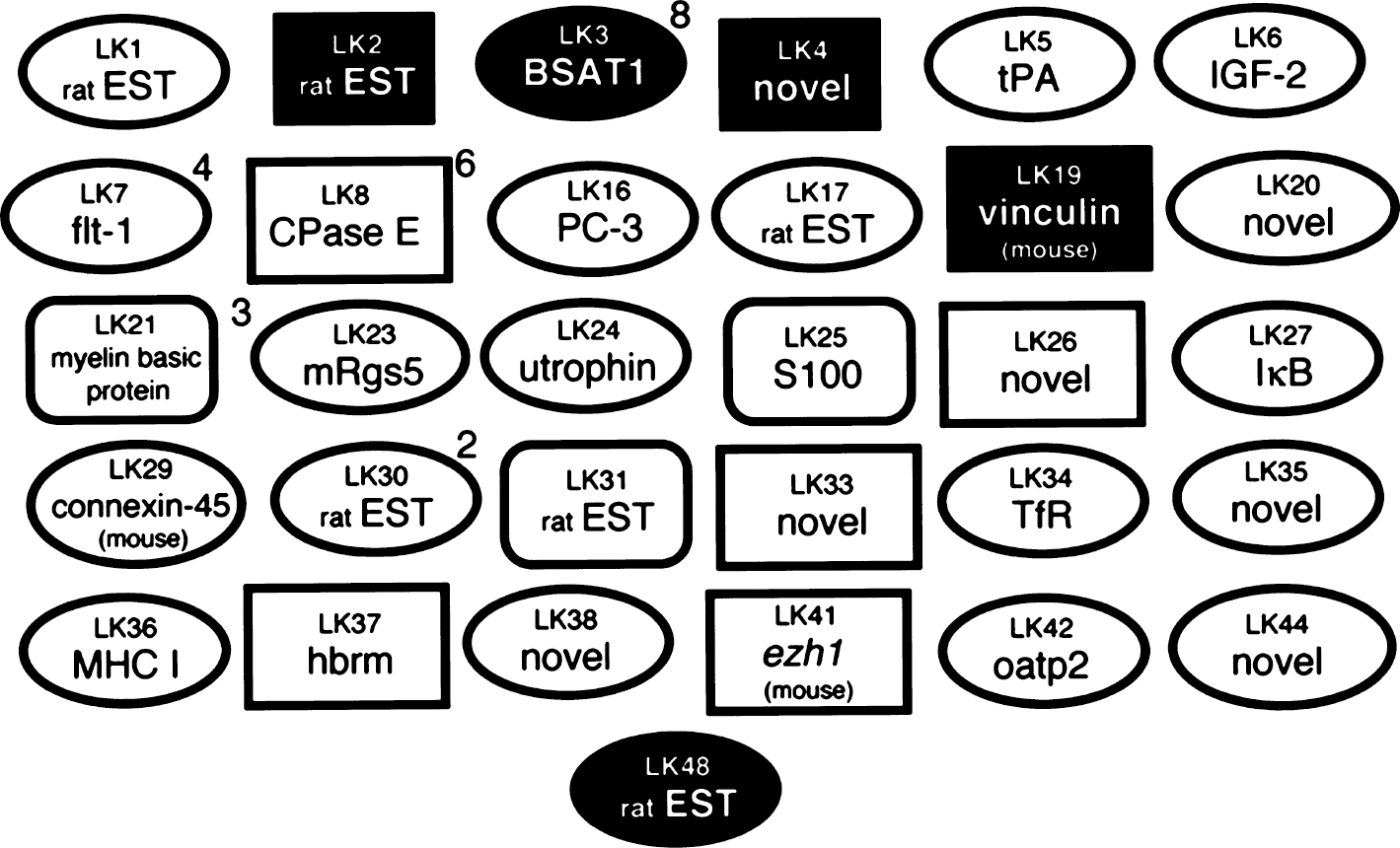
Summary of clones. Clones shown in black ovals correspond to genes that are expressed only at the BBB, and gene expression is not detectable in either total rat brain or in rat peripheral tissues. Clones shown in white ovals represent genes that are expressed only in brain at the BBB, but are also expressed in some rat peripheral tissues. Clones shown in curved white rectangles represent genes that are expressed only in brain and at the BBB, but gene expression in peripheral tissues is not detectable. Clones shown in white squares represent genes expressed widely in brain, at the BBB, and in peripheral tissues. Clones shown in black squares represent genes that are not detectable at the BBB. Numeric superscripts indicate the number of clones out of the 50 clones screened for the liver and kidney subtracted library that were detected for the same gene product.
DISCUSSION
Novel genes selectively expressed at the blood–brain barrier
Northern blotting results with clones K2, which encoded a novel sequence, and LK48, a rat EST, showed that the transcripts corresponding to these clones were only expressed at the BBB in brain and were not expressed in peripheral tissues (Figs. 2 and 5B, respectively). Clone LK48 hybridized to a 5.6 kb major transcript and a 9.5 kb minor transcript in rat brain capillaries. The clone LK48 mRNA was not detected in rat peripheral tissues and was only detected at trace amounts in whole rat brain, which may be derived from the capillary fraction in the brain homogenate (Fig. 5B).
The sequence of K2 was also novel and not found in any databases (Table 1). This clone was used to screen a rat brain capillary cDNA library in the pSPORT vector, which had been described previously (Boado et al., 1999). A 2.6 kb full length cDNA was identified and sequenced and named BBB-specific anion transporter type 1 (BSAT1), because of distant sequence similarity with a liver-specific anion transporter (Abe et al., 1999). The full sequence for BSAT1 encompassed the sequences of 8 other clones found in the initial BBB library (LK3, LK12, LK13, LK14, LK15, LK39, LK45, and LK49). Therefore, the BSAT1 clones represented 16% of the initial 50 clones identified from the liver and kidney subtracted library. This suggests that the mRNA for BSAT1 is highly enriched at the BBB; this was confirmed by Northern blot analysis as shown in Fig. 2 for clone K2. These Northern blot analysis studies were performed with a BSAT1 partial cDNA encoding for the 3'-UTR and there was no crosshybridization between this clone and mRNA generated from bovine brain capillaries as shown in Fig. 2. The failure to detect BSAT1 in the bovine brain capillary preparation suggests that the 3'-UTR of the BSAT1 mRNA is not conserved across species.
Several clones hybridized to transcripts that were expressed in brain only at the BBB, and this mRNA was also differentially expressed in some peripheral tissues (Fig. 7). Three clones (LK1, LK17, and LK30) were found in the rat EST database (Table 1). Clone LK1 hybridized to a 3.2 kb mRNA that was selectively expressed in brain at the BBB and was also expressed in rat lung (Fig. 2). Clone LK17 hybridized to a 5.6 kb mRNA that was selectively expressed in brain at the BBB and was also expressed in rat lung (Fig. 4). Clone LK30 hybridized to a 4.2 kb mRNA that was selectively expressed in brain at the BBB and was also expressed in trace amounts in rat heart (Fig. 4). In situ hybridization with sense or antisense RNA derived from clone LK30 is shown in Fig. 6 and reveals continuous immunostaining of the rat brain microvasculature, which is indicative of an endothelial origin of the mRNA. The trace signal in whole rat brain could represent LK30 mRNA derived from the rat brain microvasculature. Four additional clones (LK20, LK35, LK38, and LK44) represented novel sequences not found in the EST or Genbank databases (Table 1), and the mRNA corresponding to this cDNA was selectively expressed at the BBB in brain, in parallel with selective expression in peripheral tissues (Figs. 4 to 5). Clone LK20 hybridized to a 5.0 kb mRNA that was selectively expressed at the BBB, compared with whole rat brain or rat peripheral tissues (Fig. 5A). Clone LK35 hybridized to a 4.0 kb mRNA that was selectively expressed in brain at the BBB and was also expressed in rat heart (Fig. 4). Clone LK38 hybridized to a 5.8 kb mRNA that was selectively expressed in brain at the BBB and was also expressed at lower levels in rat heart (Fig. 5D). Part of the sequence of clone LK44 is 84% identical with nucleotides 129–320 of an EST identified in human aortic endothelium exposed to tumor necrosis factor-α (TNF-α) (Adams et al., 1995). Northern blot analysis with LK44 showed a selective expression of 5.1 and 3.5 kb transcripts at the rat BBB that were present at levels many fold greater than in total rat brain or rat peripheral tissues (Fig. 5B). The finding of a TNF-α inducible gene product at the BBB parallels previous studies showing that the receptors for TNF-a, designated TNFR1 and TNFR2, are both expressed at the BBB (Nadeau and Rivest, 1999).
Known genes selectively expressed at the blood–brain barrier
Figure 7 lists 11 known genes that are selectively expressed in brain at the BBB, with parallel expression in some peripheral tissues. These genes include tissue plasminogen activator (clone LK5); insulin-like growth factor (IGF)-2 (clone LK6); the vascular endothelial growth factor receptor, flt-1 (clones LK 7, LK10, LK18, and LK43); the PC-3 gene product (clone LK16); the regulator of G protein signaling (Rgs)-5 (clone LK23); utrophin (clone LK24); IκB (clone LK27); connexin-45 (clone LK29); the transferrin receptor (clone LK34); the class I major histocompatibility complex (clone LK36); and organic anion transporting polypeptide type 2 or oatp2 (clones K1 and LK42). The tissue-specific expression of these genes at the BBB is shown in the Northern blots in Figs. 2 to 5, and the sequence information on each clone is shown in Table 1.
Each of these gene products may play an important role in brain function. Tissue plasminogen activator (TPA) mediates neurite outgrowth and learning in brain (Seeds et al., 1999). Therefore, BBB-derived TPA may play a role in neuronal migration and synaptic connections. The mRNA for IGF-2 is highest in adult rat brain compared with any other tissue (Murphy et al., 1987). However, ISH showed IGF-2 transcript only at the choroid plexus in brain (Hynes et al., 1988), and this led to the hypothesis that choroid plexus is the site of origin of IGF-2 production in brain. However, the current results suggest that an additional source of IGF-2 in the brain is local production at the brain microvasculature. In situ hybridization studies demonstrated continuous immunostaining of isolated rat brain capillaries with the antisense probe (Fig. 6), which is indicative of an endothelial origin of the microvascular IGF-2 mRNA.
There are at least three receptors for vascular endothelial growth factor and these are designated as flt-1, flk-2/kdr, and flt-4 (Stacker et al., 1999). In whole body autoradiography, the organ with the highest binding of radiolabeled vascular endothelial growth factor is the brain and this binding in brain was restricted to both the choroid plexus and to the microvasculature (Jakeman et al., 1992). The current studies suggest that the flt-1 isoform may be the predominant vascular endothelial growth factor receptor at the brain microvasculature under normal conditions. Rgs5 acts as a GTPase activating protein for subunits of heterotrimeric G proteins (Chen et al., 1999), and these proteins play a role in the regulation of caveolin and endothelial cell transcytosis (Schnitzer et al., 1995). Utrophin is also called dystrophin related protein and is a 395 amino acid protein that is 73% identical to the cytoskeletal protein, dystrophin (Galvagni and Oliviero, 2000).
The transferrin receptor mRNA (clone LK34) is selectively localized at the BBB, and Northern blot analysis demonstrated expression of 5.0 and 6.6 kb transcripts in isolated rat brain capillaries as shown in Fig. 5D. The expression of the 6.6 kb transferrin receptor transcript was specific for rat brain capillaries and the 5.0 kb transcript was also found in total rat brain, although at reduced levels compared with isolated rat brain capillaries. The 5.0 kb transferrin receptor transcript was also detected in C6 glial cells and in rat heart, and there was no detectable transcript in rat liver. Recent studies have identified a second form of the transferrin receptor, which is encoded by mRNA of 2.9 and 2.5 kb and is specific for liver (Kawabata et al., 1999).
Clones K1 and LK42 corresponded to oatp2 and Northern blot analysis suggested the expression of this gene in the brain is confined to the BBB (Fig. 2). The K1 clone was also used in ISH and showed continuous immunostaining of isolated rat brain capillaries, indicating an endothelial origin of the oatp2 transcript (Fig. 6). The functional role of oatp2 at the BBB is not clear, but this protein may participate as an active efflux system at the BBB. A principal substrate of oatp2 is estrone sulfate (Noe et al., 1997). However, estrone sulfate does not cross the BBB in vivo (Steingold et al., 1986), which suggests that oatp2 may function as an active efflux system at the BBB.
Genes were selectively expressed in brain and at the BBB, but not in peripheral tissues. Clone LK21, myelin basic protein, clone LK25, the S100 calcium binding protein, and clone LK31, a rat EST, all hybridized to mRNA found only in brain and at the BBB, but not in rat peripheral tissues (Figs. 3 and 4). The level of the transcript for myelin basic protein in whole rat brain was comparable to the level of myelin basic protein mRNA in isolated rat brain capillaries as shown in Fig. 3. (The amount of mRNA from rat brain applied to lane 3 (2.0 μg) of the right panel in Fig. 3 was 4-fold greater than the amount of rat brain capillary mRNA applied to lane 1 or 2 (0.5 μg) of the right panel of Fig. 3.) The finding of gene expression for myelin basic protein at the BBB was unexpected. The expression of the gene for myelin basic protein at the BBB is of interest, because both myelin and the BBB evolved in parallel in all vertebrates. To further identify the site of origin of the myelin basic protein transcript at the rat brain microvasculature, ISH studies were performed and are shown in Fig. 6. The ISH shows continuous immunostaining of the brain microvessels, which is indicative of an endothelial origin of the transcript encoding for myelin basic protein. Prominent hybridization was also found in precapillary arterioles. The function of microvascular myelin basic protein is currently unknown. The earliest neuropathologic lesion in the brain of multiple sclerosis is a perivascular cuffing of lymphocytes around brain microvessels (Adams, 1977), and myelin basic protein is an autoantigen in multiple sclerosis (Bornstein et al., 1987). The current work also shows selective expression of the class I major histocompatibility complex at the BBB (clone LK 36), which suggests that antigen presentation takes place at the brain microvasculature. This extends previous work showing abundant class II major histocompatibility complex at the human brain microvasculature in multiple sclerosis (Pardridge et al., 1989).
Genes widely expressed in peripheral tissues, in brain, and at the blood–brain barrier
Carboxypeptidase E (clones LK8, LK11, LK22, LK40, LK47, and LK50), the transcription factors, hbrm (clone LK37) and EZH1 (clone LK41), and two clones of novel sequence, LK26 and LK33, were widely expressed at the BBB, in brain, and in peripheral tissues, based on Northern blot analysis (Figs. 2 to 5). The hbrm gene product is the human homologue of yeast SW12/NSF2 protein and is an activator of transcription factors (Trouche et al., 1997). The finding that several transcription factors such as PC-3, IκB, hbrm, or EZH1, are selectively expressed at the BBB suggests these proteins may regulate cell division at the brain microvasculature in states of angiogenesis.
In summary, these studies describe the initial results of a BBB genomics program and the numerous gene products that are selectively expressed at the BBB compared with whole brain. The initial library was prepared from tester cDNA derived from rat brain capillary poly A+ mRNA after subtraction with driver cDNA derived from rat liver and rat kidney poly A+ mRNA. Screening just 5% of the subtracted tester cDNA resulted in identification of 50 gene products and over 80% of these were selectively expressed at the BBB. Numerous ESTs or genes with novel sequences of unknown function were selectively expressed at the BBB and the availability of these partial cDNA will enable cloning of the full-length gene products for subsequent elucidation of the function of these genes.