
Abstract
The cytokine leukemia inhibitory factor (LIF) modulates neuronal function during development and promotes neuronal survival after peripheral nerve injury, but little is known about LIF expression after cerebral ischemia. In the present study, the localization of LIF protein was immunohistochemically examined in rats after 3.5, 12, 24, 48, and 96 hours of reperfusion following 1.5 hours of middle cerebral artery occlusion (MCAO) induced by the intraluminal suture method. Double-staining immunohistochemistry with microtubule-associated protein-2 (MAP2), glial fibrillary acidic protein (GFAP), lectin histochemistry, and interleukin (IL) 6 was also performed. The sham group and immunosorption test did not show any clear LIF immunoreactivity. Definite LIF immunoreactivity was first detected after 12 hours of reperfusion in each of the brain regions examined: ischemic core, periinfarct region, and contralateral cortex. However, expression of LIF was most prominent in the periinfarct region at each time point, peaked at 24 hours, and then gradually declined until 96 hours of reperfusion. Some LIF-positive neurons in the periinfarct region expressed IL-6. At 96 hours of reperfusion, GFAP-labeled astrocytes around the infarct core also expressed LIF protein. Induction of LIF mRNA and protein was also confirmed by reverse transcription polymerase chain reaction and western blot analysis, respectively. These findings suggest that LIF expression in ischemically threatened neurons may reflect a repair or defense mechanism against the ischemic insult.
Reperfusion after cerebral ischemia induces an inflammatory response, and several cytokines are considered to be related to its pathogenesis (DeGraba, 1998). As blocking of interleukin (IL) 1β and tumor necrosis factor-α has been found to reduce infarct size in focal cerebral ischemia in rats, these cytokines have been suggested to play a very important role in ischemic neuronal injury (Loddick and Rothwell, 1996; Barone et al., 1997). Interleukin-6, however, is also up-regulated in cortical and hippocampal neurons after cerebral ischemia (Suzuki et al., 1999a, 1999b) and considered to exert a neuroprotective effect (Mastuda et al., 1996; Loddick et al., 1998).
Leukemia inhibitory factor (LIF), which was first described as a glycoprotein that suppresses proliferation of M1 myeloid leukemic cells and induces their differentiation, is a member of the IL-6 cytokine family. Leukemia inhibitory factor exerts a broad range of effects on many types of cells and is known as a cholinergic neuronal differentiation factor that possesses a variety of potential functions in the developing and the mature nervous system (Hilton, 1992). This factor can promote the survival and outgrowth of sensory and motor neurons in vitro and after axotomy in vivo (Cheema et al., 1994a, 1994b; Gadient et al., 1998; Thompson et al., 1997). Leukemia inhibitory factor significantly improves regeneration of damaged peripheral nerves (Tham et al., 1997), and it maintains choline acetyltransferase expression in axotomized medial septal neurons (Cheema et al., 1998). It has also been reported that the numbers of spinal motor neurons in the LIF receptor knockout mouse are severely reduced (Li et al., 1995) and that administration of LIF reduces motor neuron loss in the wobbler mutant mouse (Ikeda et al., 1995). Regardless of these neurotrophic effects, little is known about the endogenous expression of LIF protein after central nervous system damage. Although LIF mRNA is induced within 6 hours after surgical injury of the cortex in the rat brain (Banner et al., 1997), induction of LIF after cerebral ischemia remains completely unknown.
The purpose of this study was to elucidate the chronological and topographic distribution of endogenous LIF protein by immunohistochemistry in a transient focal cerebral ischemia model in rats.
MATERIALS AND METHODS
Animal preparation
The protocol described here received prior approval at a meeting of the Committee on Animal Experimental Guidelines of Keio University School of Medicine. Adult male Sprague-Dawley rats (Japan Laboratory Animals, Tokyo, Japan) weighing 300 to 340 g were divided into two groups: a middle cerebral artery occlusion (MCAO) group (n = 50) and a sham group (n = 8). The animals in the MCAO group were anesthetized with a mixture of 1.0 to 1.5% halothane and 30% oxygen/70% nitrogen during the operation. A temperature probe (TD-300; Shibaura Electronics, Tokyo, Japan) was inserted into the rectum, and a heat lamp was used to maintain rectal temperature at 37.0°C to 37.5°C.
Middle cerebral artery occlusion was induced by the intraluminal filament technique of Koizumi et al. (1986), as modified by Belayev et al. (1996). In brief, a midline surgical incision was made to expose the right common, internal, and external carotid arteries. The distal portion of the external carotid artery was ligated, and the common and internal carotid arteries were temporarily occluded with clips (Sugita aneurysm clips; Mizuho Ikakogyo, Tokyo, Japan). A small incision was then made in the proximal portion of the external carotid artery, and a 3-0 nylon monofilament suture (Matsuda Ikakogyo, Tokyo, Japan) was inserted into the internal carotid artery. The distal segment of the suture had been coated with poly-L-lysine solution (Sigma, St. Louis, MO, U.S.A.) to ensure consistent ischemia in the brain. The filament was advanced until it obstructed the origin of the right MCA. After the intraluminal filament was fixed in position, the neck incision was closed with silk thread.
The animals were allowed to recover from the anesthesia and were evaluated for the extent of their neurological deficits 1 hour after MCAO. The deficits were graded as severe, moderate, mild, or absent according to the method of neurologic examination devised by Bederson et al. (1986). In brief, animals that consistently circled toward the left (paretic) side received a grade of 3. Rats that consistently showed reduced resistance to lateral push toward the left side with no circling movement received a grade of 2. Rats with any amount of consistent forelimb flexion and no other abnormalities were graded as 1. Rats with no neurological deficits were graded as 0. Only animals with grade 2 or 3 deficits were utilized for further study as the MCAO group (n = 24), as these animals were expected invariably to have a large infarct area in the territory of the MCA (Bederson et al., 1986). After 90 minutes of MCAO, the rats were reanesthetized with the same anesthetic combination as above, and the intraluminal suture was carefully removed. The common and internal carotid arteries were then inspected to ensure the return of good pulsation. The neck incision was closed with a silk suture, and the animals were allowed to survive with free access to food and water. In the sham group, the neck incision was made only to expose the right common, external, and internal carotid arteries, and a suture was not inserted into the artery. Other procedures were identical to those in the MCAO group.
Reverse transcription polymerase chain reaction
The animals were decapitated after 3.5 (n = 2) and 24 (n = 2) hours of reperfusion. Sham-operated rats served as controls (n = 2). Samples were taken from the periischemia region in the cerebral cortex, as shown in Fig. 1. Acidic phenol/chloroform extraction was used to extract RNA, and 250 ng of RNA was reverse-transcribed in a total volume of 20 μL according to the manufacturer's instructions (Life Technologies, Gaithersburg, MD, U.S.A.). Two microliters of the first-strand synthesis was used for PCR. The 50-μL PCR reaction additionally contained 0.5 μmol/L of each primer. The LIF primers were as follows: forward, 5′-ATGAAGGTCTTGGCCACAGG-3′; reverse, 5′-TGGACCACCGCACTAATGAC-3′; this resulted in a PCR product of 599 bp (Takahama et al., 1998). Amplification was performed at the following temperature profile: 60 seconds at 94°C, 30 seconds at 60°C, and 60 seconds at 72°C (27 cycles). Reaction products were then separated on a 2% polyacrylamide gel, stained with ethidium bromide, and photographed. Each set of PCR reactions included control samples run without RNA or samples in which the RT step was omitted.
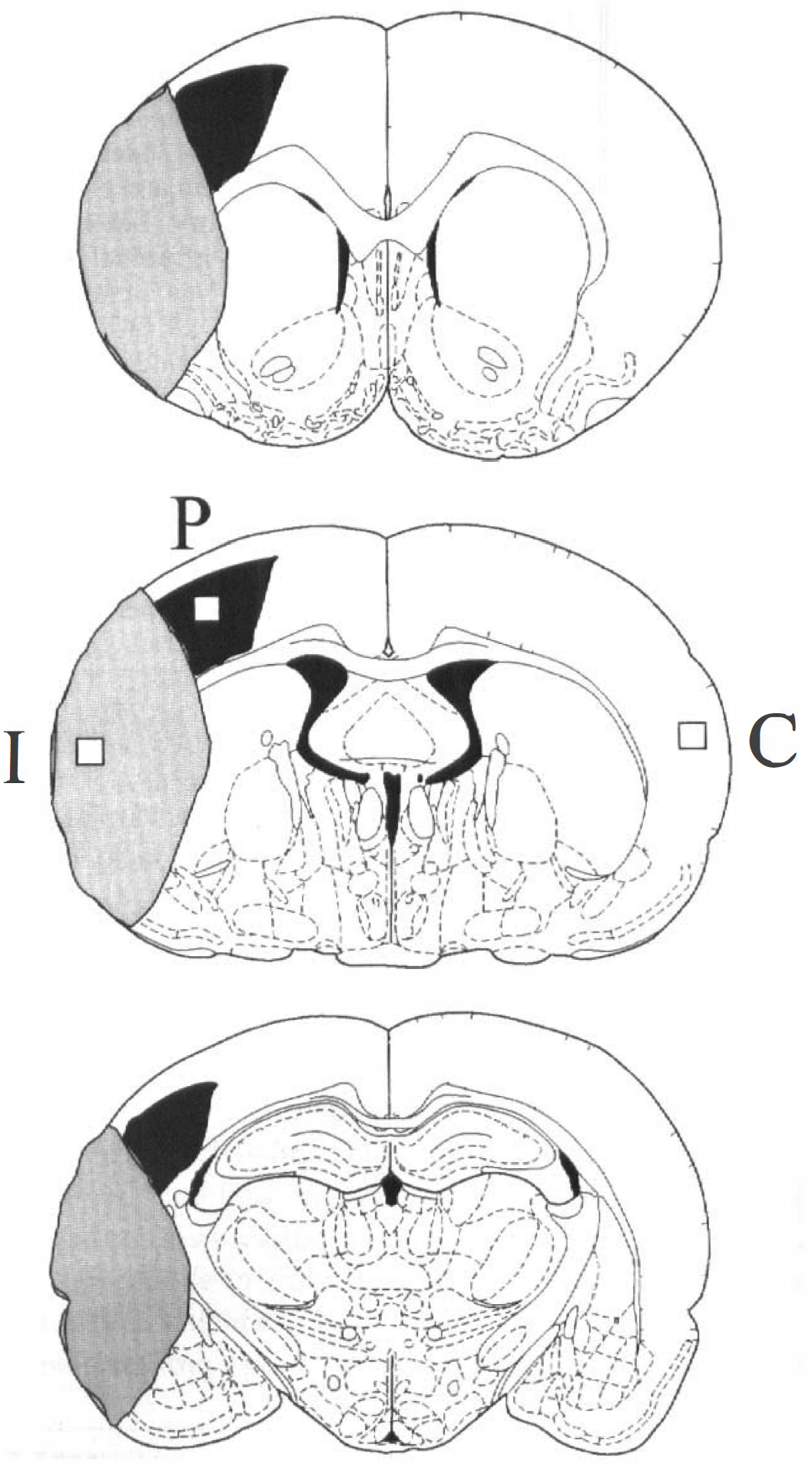
A schematic diagram of the rat brain. With guidance from a rat brain atlas (Paxinos and Watson, 1986), coronal sections were taken at the level of the caudate putamen, globus pallidus, and hippocampus. The infarct area is shown lightly shaded. The distribution of prominent leukemia inhibitory factor (LIF) immunoreactivity is shown darkly shaded. Three different areas of cerebral cortex in which the numbers of LIF-immunopositive cells were counted are also indicated (I, ischemic core; P, periischemia region; C, contralateral cortex).
Western blot analysis
The animals were decapitated after 3.5 (n = 2) and 24 (n = 2) hours of reperfusion. Sham-operated rats served as controls (n = 2). Samples were taken from the entire cerebral cortex on the occluded side. Tissue was homogenized in a Teflon-coated homogenizer containing buffer (0.32 mol/L sucrose, 1 mmol/L ethylenediaminetetraacetate, 5 mmol/L Tris-HCl, pH 7.4, 0.1 mmol/L phenylmethylsulfonyl fluoride, 10 μmol/L leupepsin, 10 μmol/L pepstatin A, and 1 mmol/L β-mercaptoethanol). Samples containing 50 μg of protein each or mouse recombinant LIF protein (Sigma) were subjected to 10% polyacrylamide gel electrophoresis in the presence of 0.1% sodium dodecyl sulfate. Protein bands were transferred from the gel to a nitrocellulose membrane (Hybond ECL; Amersham, Buckinghamshire, U.K.) by employing a transfer apparatus and a blotting buffer including 0.192 mol/L glycine and 5% methanol (pH 8.4). The membrane was then incubated with a 1:150 dilution of polyclonal goat anti-LIF antibody (Santa Cruz Biotechnology, Santa Cruz, CA, U.S.A.; cat. no. sc-1336). The membrane was next reacted with horseradish peroxidase-linked anti-goat antibody (Santa Cruz Biotechnology). Protein bands were detected by an enhanced chemiluminescence method (ECL Kit; Amersham). The protein concentration of each sample was determined with a Bio-Rad protein assay kit (BioRad Laboratories, Hercules, CA, U.S.A.), using bovine serum albumin as the standard.
Immunohistochemistry
Animals were anesthetized by intraperitoneal injection of pentobarbital 3.5 (n = 4), 12 (n = 5), 24 (n = 5), 48 (n = 5), or 96 (n = 5) hours after the start of reperfusion or 24 hours after the sham surgery (n = 4). They were then perfused with 200 mL of 50 mmol/L phosphate-buffered saline (PBS; pH 7.4), followed by 200 mL of 4% freshly depolymerized paraformaldehyde in 0.1 mol/L phosphate buffer (pH 7.4). The brains were removed and postfixed for 6 hours in 4% paraformaldehyde in phosphate buffer at 4°C before cryoprotection by sequential bathing in 10, 15, 20, and 30% sucrose. The brains were then frozen in powdered dry ice, and consecutive 20-μm-thick coronal sections or 16-μm-thick sections for double-staining were prepared on a cryostat (Cryocut 1800; Leica Instruments GmbH, Nussloch, Germany). The sections were thaw-mounted onto gelatin/chrome alum-coated slides and stored at −30°C. They were subsequently rehydrated and washed in PBS (pH 7.4) for 5 minutes, quenched with 0.3% H2O2 in PBS for 20 minutes to block endogenous peroxidase, and washed with PBS three times for 5 minutes each. Next, they were preincubated for 30 minutes in a solution containing skimmed milk to block nonspecific binding, and the sections were incubated overnight at 4°C with a 1:100 dilution of polyclonal goat anti-LIF antibody (same as above). The slides were reacted with an avidin-biotinylated enzyme complex system (Vectastain ABC Elite Kit; Vector Laboratories, Burlingame, CA, U.S.A.), and the immunoreactive product was visualized by the diaminobenzidine reaction. To examine the specificity of the immunoreactivity, the primary antibody was omitted to provide a nonspecific control. In addition, immunosorption testing was carried out by using a mixture of primary antibody and recombinant mouse LIF protein (same as above; concentration 5 ng/μL).
Absolute immunoreactive cell count (per 0.15 mm2) with an ocular micrometer (Nikon, Tokyo, Japan) attached to a light microscope at ×160 magnification was made in a blinded manner by one of the co-authors. Based on a rat brain atlas (Paxinos and Watson, 1986), the ischemic core (parietal cortex area 2) and the periischemia region (hindlimb area of the cortex) of the cerebral cortex, the contralateral cerebral cortex (parietal cortex area 2, corresponding to the ischemic core on the surgery side), and the corresponding areas of the sham group at the level of the globus pallidus (bregma −0.8 mm) were selected for cell counting, as shown in Fig. 1. Immunoreactive cells in the pyramidal cell layers of bilateral hippocampal CA1 (per 0.5 mm) were also counted. Counting was performed on two consecutive slices, and the mean value for each area was calculated.
All data are expressed as means ± SD. The data were tested by analysis of variance followed by post hoc analysis with the Bonferroni correction for multiple simultaneous comparisons.
In additional histochemical studies, antibodies were applied to adjacent sections and detected with diaminobenzidine to identify the cellular source of LIF. Monoclonal anti-microtubule-associated protein-2 (anti-MAP2) antibody (dilution 1:500; Boehringer-Mannheim, Philadelphia, PA, U.S.A.) and anti-glial fibrillary acidic protein (anti-GFAP) antibody (dilution 1:300; Boehringer-Mannheim) were used to identify neurons and astrocytes, respectively. Monoclonal antiparvalbumin antibody (dilution 1:2,000; RBI, Natick, MA, U.S.A.) was used to identify GABAergic neurons. Microglia and monocytes/macrophages were also histochemically identified by using isolectin-B4 from Griffonia simplicifolia seeds (GSAI-B4; Sigma), as described (Streit, 1990; Streit and Krkeutzberg, 1987). In brief, the sections were incubated overnight at 4°C with isolectin (concentration 20 μg/mL) in PBS containing divalent cations. Adjacent sections were also stained with cresyl violet for conventional histological examination.
Double-staining immunohistochemistry
MAP2/LIF, GFAP/LIF, and GSAI-B4/LIF double-staining immunohistochemistry was performed by treatment first with anti-MAP2 antibody, anti-GFAP antibody, or GSAI-B4 staining and then staining with a fluorescein isothiocyanate-conjugated anti-mouse IgG antibody (Amersham). The sections were washed, and anti-LIF antibody was applied and stained with diaminobenzidine. To compare the distribution of LIF with that of IL-6 protein, we also performed LIF/IL-6 double-staining. As reported previously (Suzuki et al., 1999a, 1999b), we used polyclonal rabbit anti-mouse IL-6 antibody (dilution 1:100; Genzyme Corp., Cambridge, MA, U.S.A.). The LIF/IL-6 double-staining was performed overnight at 4°C by using a mixture of two corresponding primary antibodies. The Cy2-conjugated anti-goat IgG antibody (Jackson ImmunoResearch Laboratories, West Grove, PA, U.S.A.) was used to detect LIF, and IL-6 was detected with a Texas Red-conjugated anti-rabbit IgG antibody (Amersham).
RESULTS
Reverse transcription polymerase chain reaction
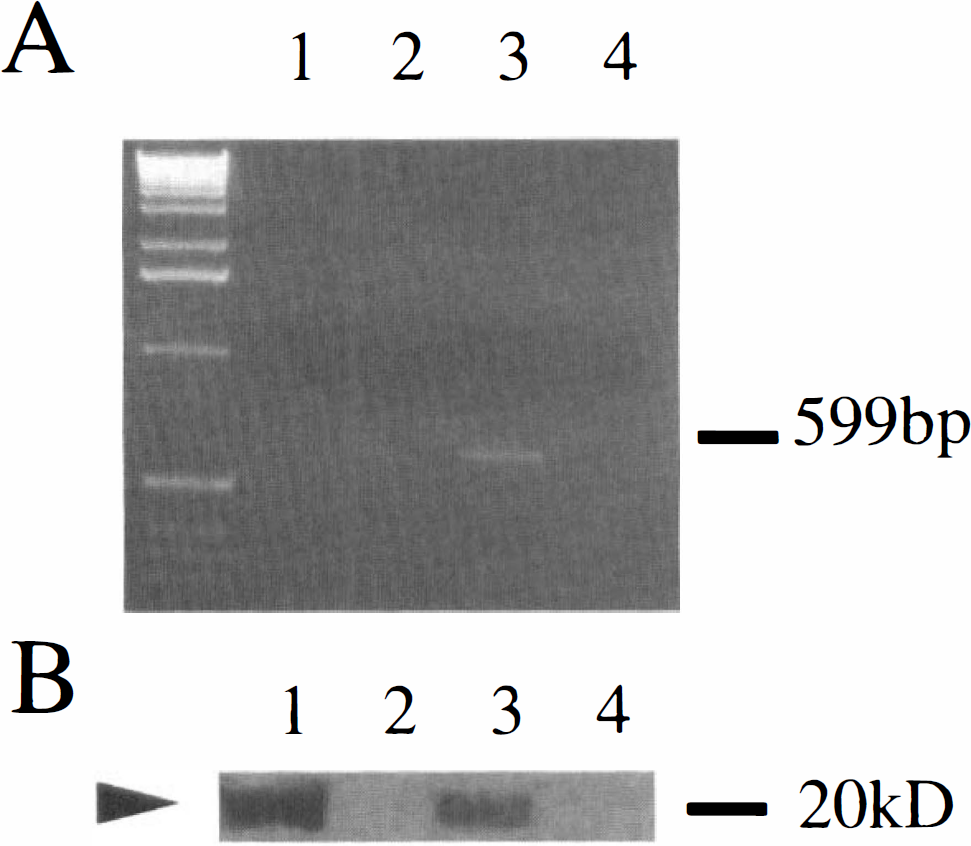
Leukemia inhibitory factor mRNA expression was detected in samples taken from animals subjected to 24 hours of reperfusion after ischemia but not in samples from 3.5-hour reperfusion or the sham group (Fig. 2A).
Western blot analysis
A single band with a molecular mass of 20 kDa was detected as recombinant LIF protein (Fig. 2B). A similar band having the same molecular mass was detected in samples taken from the animals subjected to 24 hours of reperfusion after ischemia but not in samples from the 3.5-hour reperfusion or the sham group.
Immunohistochemistry
Cresyl violet staining clearly showed the infarct area in the frontoparietal somatosensory cortex and the caudate putamen on the operated side from 24 hours of reperfusion after MCAO onward. Representative photomicrographs of sections stained with anti-LIF antibody are shown in Fig. 3. No clear LIF immunoreactivity was detected in any regions of the cerebral cortex in the sham group or at 3.5 hours of reperfusion after MCAO (Fig. 3A and B). The LIF immunoreactivity was first detected after 12 hours of reperfusion, and the periischemia region exhibited more marked staining than the ischemic core and the opposite side. In the periischemia region, many cortical neurons expressed LIF protein from 12 to 96 hours of reperfusion, as shown in Fig. 1. The LIF immunoreactivity peaked at 24 hours of reperfusion, after which there was a gradual decrease until 96 hours (Fig. 3C to Fig. 3F). Not only the perikarya but also the axons and dendrites of these cortical neurons stained with anti-LIF antibody.
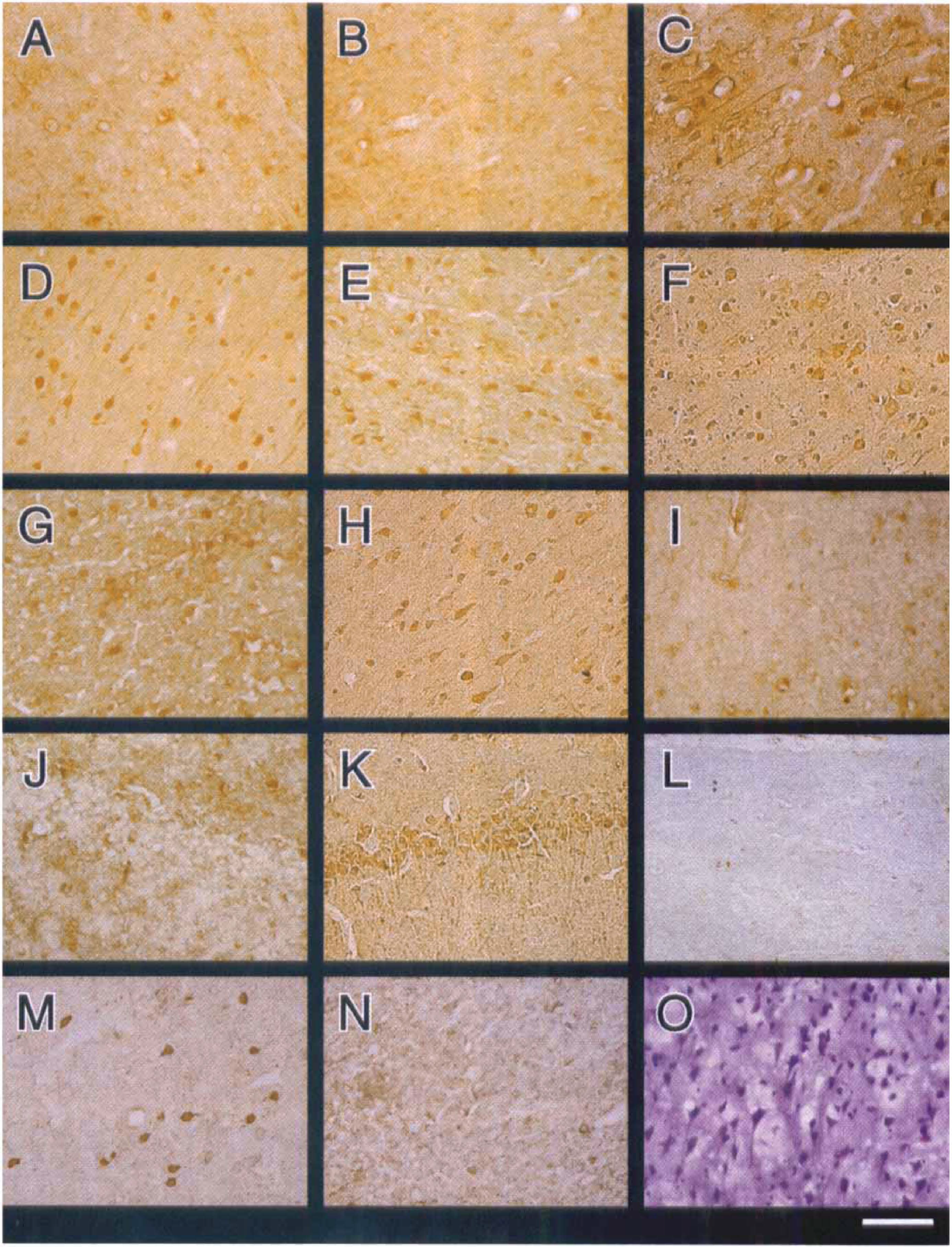
Representative photomicrographs showing anti-leukemia inhibitory factor (anti-LIF) antibody immunoreactivity in the periischemia region
In the ischemic core, LIF protein was partially expressed in ischemically damaged shrunken neurons having triangular cytoplasm (Fig. 3G). Cresyl violet staining clearly demonstrated ischemic damage after 12 hours of reperfusion. After 48 hours of reperfusion, complete necrotic change was observed. On the opposite side, LIF protein was expressed in some cortical neurons from 12 to 96 hours of reperfusion (Fig. 3H). Although no LIF immunoreactivity was observed in the hippocampus (Fig. 3J), bilateral hippocampal neurons (CA1, CA2, CA3, and dentate gyrus) and their dendrites also expressed LIF protein from 12 hours of reperfusion (Fig. 3K). The LIF immunoreactivity was not detected when anti-LIF antibody was preincubated with recombinant mouse LIF (Fig. 3I and L) or when the primary antibody was omitted. GABAergic neurons were detected in the periischemia region (Fig. 3M). Taken together with the immunoreactivity of neurons for LIF on the adjacent sections, some LIF-immunopositive neurons in the periischemia region were considered to be GABAergic. On the other hand, apparent parvalbumin immunoreactivity was not detected in the ischemic core (Fig. 3N). Cresyl violet staining revealed neurons in the periischemia region could survive up to 28 days following 90 minutes of ischemia (Fig. 3O).
The results of MAP2/LIF, GFAP/LIF, and GSAI-B4/LIF double-staining immunohistochemistry are shown in Fig. 4. Intense MAP2 immunoreactivity was detected especially in the periischemia region from 12 hours of reperfusion onward, as previously reported (Li et al., 1998). The LIF immunoreactivity was clearly co-localized with MAP2 in this region (Fig. 4A and B). Although the GFAP-labeled astrocytes proliferated around the surface of the ischemic cortex beginning at 24 hours of reperfusion, LIF was not expressed in the astrocytes at 24 or 48 hours of reperfusion. At 96 hours of reperfusion, LIF immunoreactivity was co-localized with markedly proliferating astrocytes showing thick processes (Fig. 4C and D). Lectin histochemistry detected some morphologically activated microglia and monocytes/macrophages in the ischemic core. As reported by Zhang et al. (1997), round types and highly ramified types of microglia gradually increased by 96 hours of reperfusion. The distribution of reactive microglia and monocytes/macrophages was clearly different from that of LIF immunoreactivity (Fig. 4E and F). The LIF/IL-6 double-staining showed that some cortical neurons in the periischemia region expressed LIF and IL-6 simultaneously (Fig. 4G to Fig. 4I).
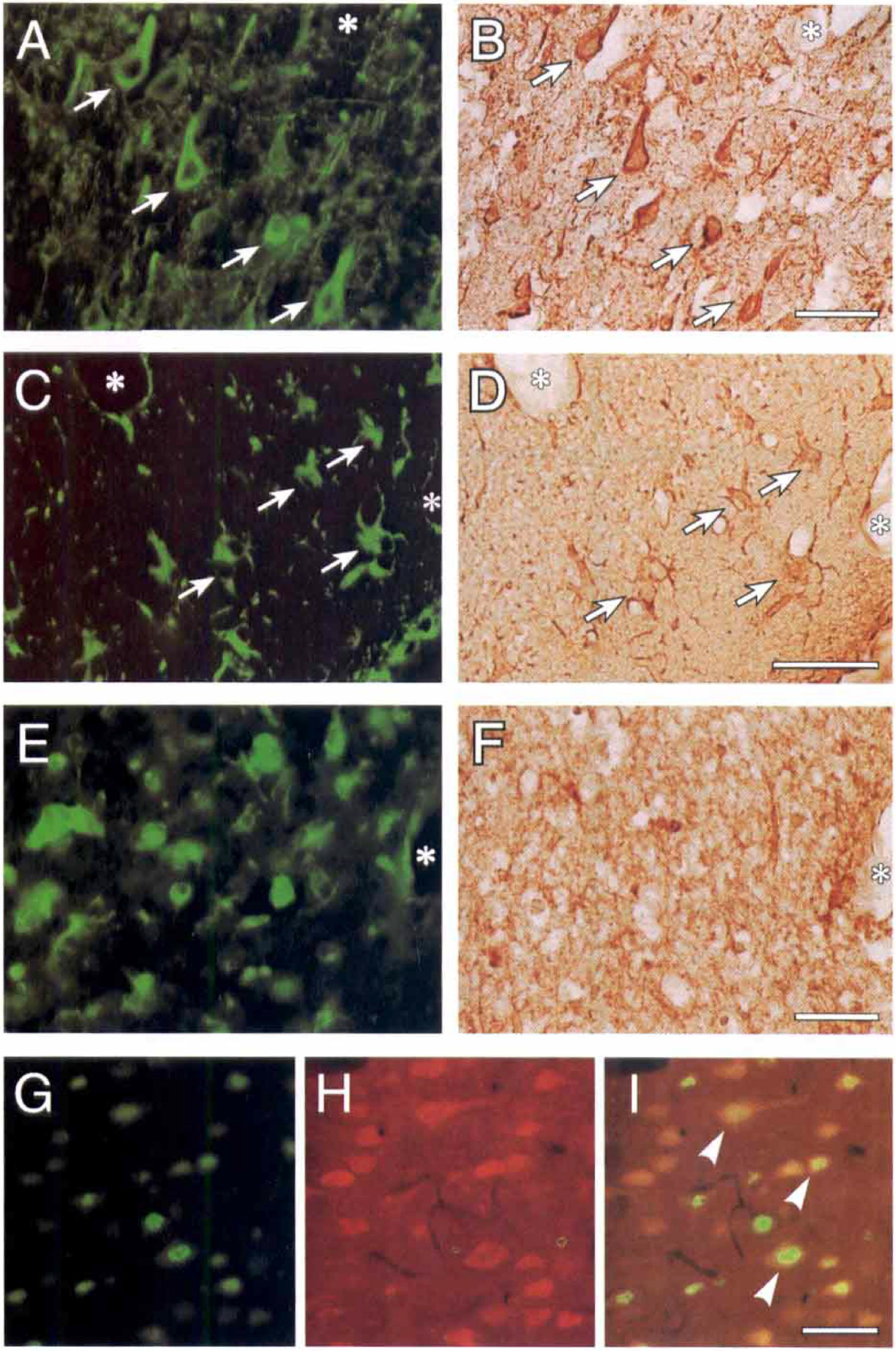
Figure 5 summarizes the number of LIF-immunopositive neurons in the three different regions: ischemic core, periischemia region, and contralateral cortex. In the sham group and the MCAO group with 3.5 hours of reperfusion, the number of immunopositive cells was similar in each region. At 12 hours of reperfusion, the number of immunopositive cells was markedly increased in all regions examined, but the increase in the periischemia region was most prominent, and the number peaked at 24 hours of reperfusion. The numbers of LIF-immunopositive cells in each region remained significantly higher than in the sham group until 96 hours of reperfusion.
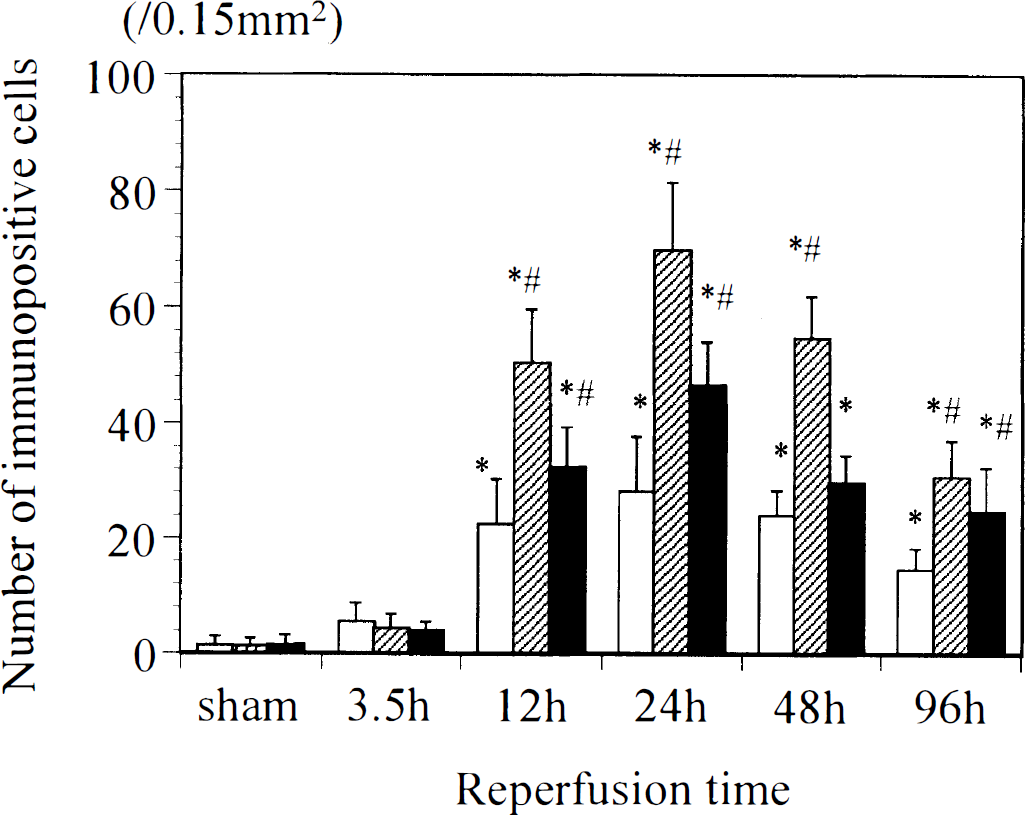
Temporal profiles showing the numbers of leukemia inhibitory factor-immunopositive cells in three different regions of the cerebral cortex. The locations of the ischemic core (open columns), periischemia region (shaded columns), and contralateral cortex (filled columns) are shown in Fig. 1. *P < 0.01 versus sham group; #P < 0.01 versus ischemic core.
Figure 6 summarizes the number of LIF-immunopositive neurons in the pyramidal cell layer of the CA1. Similar to Fig. 5, LIF immunoreactivity became positive at 12 hours of reperfusion and peaked at 24 hours.
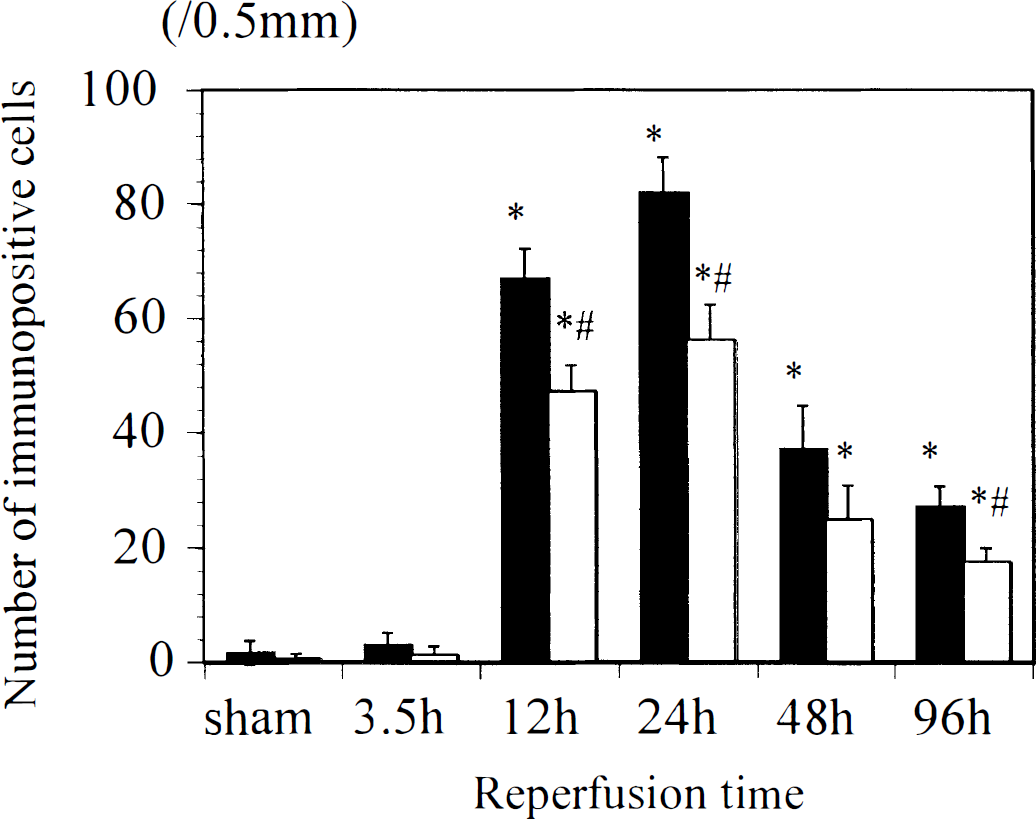
Temporal profiles showing the numbers of leukemia inhibitory factor-immunopositive cells in the bilateral hippocampal CA1. Presented are the ischemic side (filled columns) and the contralateral cortex (open columns). *P <0.01 versus sham group; #P < 0.01 versus ischemic side.
DISCUSSION
The neuropoietic cytokine LIF can act as a trophic factor, enhancing neuronal survival and differentiation (Hilton, 1992). Although several cytokines are known to be up-regulated in the brain in response to ischemia (Barone et al., 1997; Davies et al., 1999), there have been no reports on LIF expression after cerebral ischemia. In the present study, we have demonstrated that LIF protein was substantially expressed in the cortical and hippocampal neurons from 12 to 96 hours of reperfusion following 90-minute focal ischemia. At 96 hours of reperfusion, proliferating astrocytes around the surface of the infarcted cerebral cortex also express LIF. The prominent up-regulation of LIF could be detected in both mRNA and protein levels by RT-PCR and western blot analysis, respectively.
Leukemia inhibitory factor mRNA has been shown to be predominantly expressed in both the cholinergic and the GABAergic neurons in the normal adult rat brain (Lemke et al., 1996, 1997). It was also demonstrated not to be induced in activated microglia and reactive astrocytes in response to basal forebrain cholinergic lesion created by injection of 192IgG-saporin (Lemke et al., 1999). Banner et al. (1997), on the other hand, demonstrated an increase in LIF mRNA, localized mainly in astrocytes, within 6 hours of a surgical lesion of the cortex in adult rat brain. In contrast, we observed that LIF was not expressed in astrocytes until 48 hours of reperfusion after MCAO. Although GSAI-B4 also stains cells of the monocyte lineage and endothelial vessels, we think that GSAI-B4 detects mostly reactive microglia and monocytes/macrophages. Our observations have shown that no reactive microglia and monocytes/macrophages in any region expressed LIF protein at any time during the reperfusion. Although the cellular origin of the LIF in the affected brain depends on the nature of the brain injury, we think that neurons are the major source of LIF protein after ischemic insults.
As we did not directly examine the role of LIF in the present study, the marked neuronal expression of LIF protein in the periischemia region is of great interest. The rat MCAO model produced by the suture method is the most popular model and has been repeatedly utilized for experimental cerebral ischemia (Belayev et al., 1996). The periischemia region in this model is considered to be a penumbra area that can be rescued by pharmacological agents (Memezawa et al., 1992). Accordingly, numerous histological and neurochemical reactions in this region triggered by reperfusion after cerebral ischemia have been attracting the interest of many investigators. The neuroanatomical remodeling in the neocortex may be involved in the mechanisms underlying the recovery of nervous function after rat MCAO (Stroemer et al., 1995). In fact, an immunohistochemical study with neuronal markers has shown neurite regrowth in dendrites and axons and neuronal plasticity in this region (Li et al., 1998). In addition to its neurotrophic effects, LIF can enhance neurotrophin expression and corticospinal axonal growth (Blesch et al., 1999). We observed that the dendrites and axons in the periischemia region were clearly stained with anti-LIF antibody. We speculate that the neuronal LIF expression in this region may be closely associated with the compensatory and repair mechanisms against ischemia-induced damage in neurons. In fact, we also observed that neurons in the periischemia region could survive during longer perfusion periods (up to 28 days following 90 minutes of ischemia).
The family of cytokines signaling through the common receptor subunit gp130 comprises IL-6, IL-11, LIF, oncostatin M, ciliary neurotrophic factor, and cardiotrophin-1 (Hibi et al., 1996; Heinrich et al., 1998). These so-called IL-6-type cytokines play an important role in the regeneration of complex cellular processes. The gp130 is widely distributed in both glial and neuronal cells in the rat brain (Watanabe et al., 1996). However, as most LIF receptor mRNA has been detected on cortical and hippocampal neurons (Yamakuni et al., 1996), neurons can be considered the main target of LIF.
We previously demonstrated that IL-6 protein is expressed in cerebral neurons after transient forebrain ischemia in gerbils and after focal ischemia in rats (Suzuki et al., 1999a, 1999b). The induction of IL-6 started earlier after MCAO, at 3.5 hours, than the induction of LIF. The IL-6 protein was also expressed in reactive microglia in addition to neurons. Interleukin-6 and LIF have some similar functional properties, and they stimulate the same intracellular signals (Heinrich et al., 1998). An antibody to gp130 can block the effect of these cytokines (Guo et al., 1999). Interleukin-6 synthesis is regulated by other members of the IL-6 cytokine family, such as LIF (Villiger et al., 1993). As IL-6 and LIF have different kinetic patterns after MCAO, both cytokines may have the same role in ischemic insults. In fact, intraventricular administration of recombinant IL-6 can prevent the delayed neuronal loss in gerbils and reduce the ischemic damage in rats (Matsuda et al., 1996; Loddick et al., 1998). In the present study, we were able to demonstrate that some neurons in the periischemia region simultaneously express both IL-6 and LIF. Moreover, LIF was expressed in the contralateral hemisphere like the expression of IL-6, which has been reported previously (Suzuki et al., 1999b). We suspect that spreading depression or ischemia-induced depolarization may be one of the most important factors for contralateral expression of LIF. Taking these results together, we speculate that neurons may be able to produce LIF in response to ischemic stress and that LIF may act as a neuroprotective mediator in a paracrine manner.