
Abstract
Postischemic cerebral inflammation has been reported to contribute to ischemic brain damage. During inflammation, constituents of the extracellular matrix such as fibronectin and laminin are recognized by certain integrins or proteoglycans and play an important role in the cell adhesion process. The purpose of this study was to evaluate the efficacy of peptides derived from laminin on leukocyte accumulation, infarct size, and neurological outcome in rats subjected to 1 h of cerebral ischemia and 48 h of reperfusion. Forty-four animals were included in this study: transient ischemia without treatment (Group I), treatment with TG-1 peptide (Group II), GD-1 peptide (Group III), and GD-6 peptide (Group IV). Group II showed a significant reduction of the leukocyte accumulation (p < 0.001) and infarct size (p = 0.015) when compared with Group I. The neurological grade of Group II was also significantly better than in Group I at 48 h after reperfusion (p = 0.012). Based on these data, which are the first to explore the therapeutic potential of this peptide in cerebral ischemia, laminin peptide may offer a novel therapeutic approach to allaying injury in ischemic stroke.
Cellular inflammation plays a critical role in the elaboration of a variety of types of central nervous system injury. Polymorphonuclear leukocytes (PMN) and monocytes are directly involved in the pathogenesis and development of ischemia-reperfusion injury by reducing microvascular blood flow, initiating thrombosis, and releasing chemical mediators such as free oxygen radicals (Schmid-Schönbein and Engler, 1986; Kochanek and Hallenbeck, 1992; Matsuo et al., 1995). One therapeutic approach targeting cell-mediated inflammation has been to block the function of specific adhesion molecules using selective antibodies. Much emphasis has been placed on the interaction between β2 integrin (Mac-1) and intercellular adhesion molecule-1 in mediating leukocyte recruitment to sites of cerebral infarction and the potential value of blocking this interaction in the control of PMN emigration (Clark et al., 1991; Bowes et al., 1993; Chen et al., 1994).
The extracellular matrix also contributes to the cell adhesion process during inflammation (Butcher, 1991). Constituents of the extracellular matrix such as fibronectin and laminin are recognized by certain integrins and proteoglycans (Skubitz et al., 1991; Gehlsen et al., 1992; Iida et al., 1992; Woods et al., 1993). We recently have demonstrated that synthetic peptides derived from fibronectin are effective in brain protection after ischemia/reperfusion by preventing leukocyte accumulation in the ischemic tissue (Yanaka et al., 1996a, d 1997a,b. It has been shown that the extracellular matrix proteins such as fibronectin and laminin interact with the same families of receptors (integrins and proteoglycans). The positive results that we obtained in those studies led us to study peptides derived from laminin.
During an inflammatory response, laminin also plays an important role in leukocyte recruitment (Akiyama and Yamada, 1987; Martin and Timpl, 1987; Beck et al., 1990). Laminin is a multifunctional basement membrane glycoprotein, composed of three large polypeptide chains (α, β1, and β2) held together by disulfide bonds (Timpl et al., 1979). Laminin has a cruciform configuration with two or three small globular domains on the short arms and one large globular domain at the distal end of the long arm (Fig. 1) (Engel et al., 1981). Several functional attributes of laminin have been elucidated. These include mediating the attachment and migration of cells, participating in cellular differentiation and tumor metastasis, and interacting with other basement membrane components such as type IV collagen and proteoglycans (Chung et al., 1979; Terranova et al., 1983; Goodman et al., 1991). Polymorphonuclear leukocytes move through the basement membrane and interact with laminin during inflammation. Laminin has been found to promote the adhesion of PMN (Terranova et al., 1986; Bryant et al., 1987; Suchard et al., 1991; Nourshargh et al., 1992). Polymorphonuclear leukocytes contain a variety of proteolytic enzymes such as elastase that specifically degrade laminin into smaller fragments (Heck et al., 1990). Peptide fragments resulting from the proteolytic degradation of laminin may exist in high concentrations during PMN migration through the basement membrane (Harvath et al., 1994). Furthermore, we have previously shown that several laminin peptides promote the chemokinesis of PMN (Harvath et al., 1994).
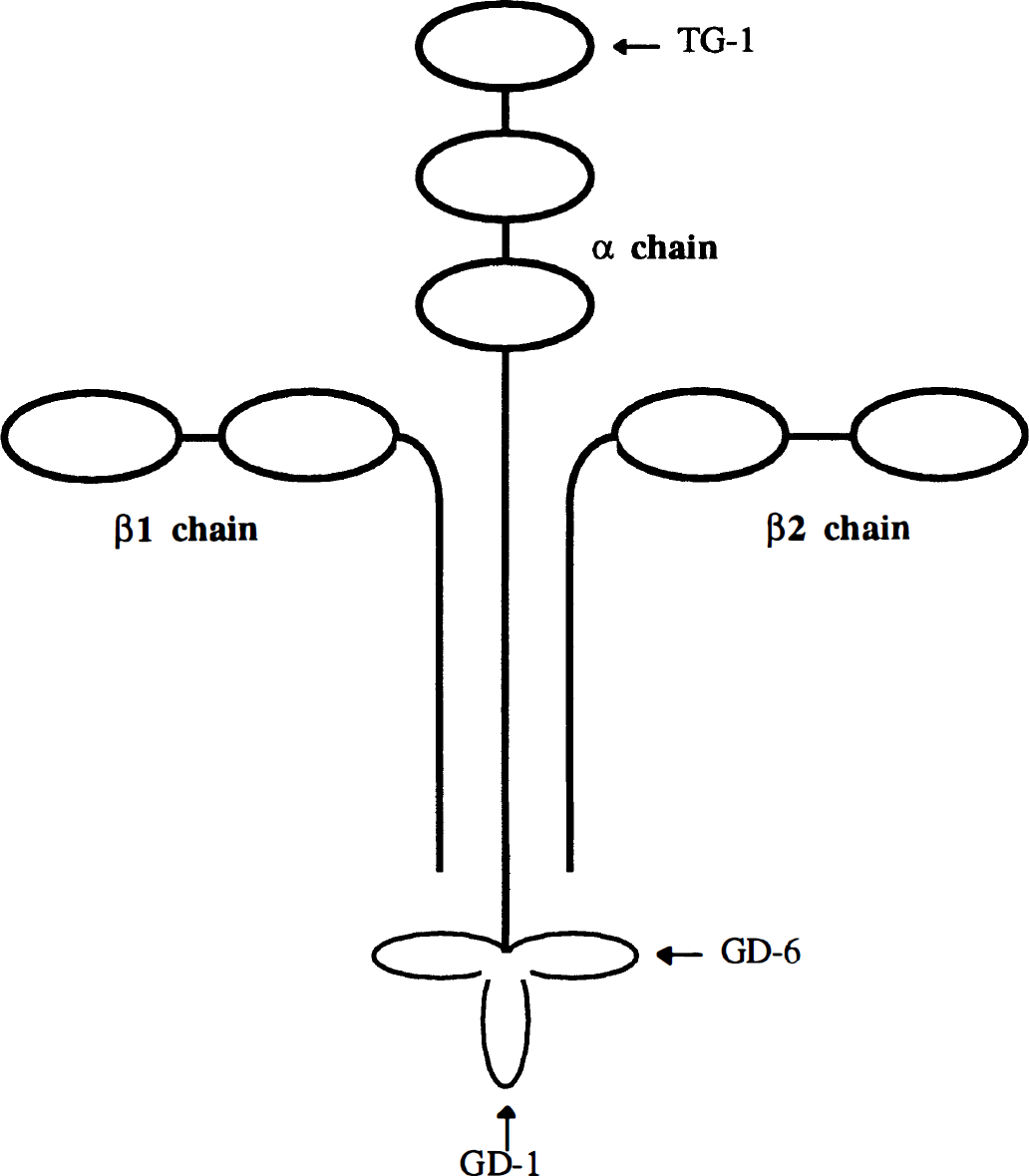
Sketch of the laminin molecule illustrating regions corresponding to synthetic peptides.
The present study was undertaken to evaluate the efficacy of peptides derived from the α chain of laminin on brain protection after transient focal cerebral ischemia in rats.
METHODS
General protocol
Fifty-two male Sprague-Dawley rats, weighing 270–350 g, were used and 44 animals were included in this study. All procedures were approved by the institutional animal care committee and performed under the auspices of Research Animal Resources, an American Association for the Accreditation of Laboratory Animal Care-approved facility.
The animals were allocated to one of four groups: transient cerebral ischemia without treatment (Group I), treatment with intravenous administration of TG-1 peptide (Group II), treatment with intravenous administration of GD-1 peptide (Group III), and treatment with intravenous administration of GD-6 peptide (Group IV). The animals in Groups II, III, and IV received intravenous administration of 150 µl of saline containing individual peptide (5 mg/kg) four times: 30 min prior to ischemia, at the time of reperfusion, 3 h after reperfusion, and 24 h after reperfusion. Eight animals were excluded from the study: One animal from Group I, one from Group III, and two from Group IV were excluded because of the presence of subarachnoid hemorrhage. One animal from Group I and one from Group II were excluded because of the absence of neurological deficits at 3 h after the induction of ischemia. Two animals from Group III were also excluded because of the failure of maintaining appropriate PCO2 level during surgery.
Animal preparation and monitoring
All animal procedures were done under complete general anesthesia. Following the induction of anesthesia with intraperitoneal injection of a mixture of 87 mg/kg ketamine and 13 mg/kg xylazine, the animals were ventilated through a face mask with a mixture of oxygen and air. The arterial PCO2 was maintained between 35 and 40 mm Hg. The rectal temperature was maintained between 37 and 38°C with heating pads. The right femoral artery and vein were cannulated for measuring blood gases and collecting blood samples and for drug administration, respectively. Peripheral blood samples were obtained from all animals included in the study before induction of ischemia and at 48 h after reperfusion for measurement of the number of peripheral white blood cells and differentials (Cell-dyne 3500; Abbott Diagnostics, San Jose, CA, U.S.A.).
Transient focal cerebral ischemia in the area perfused by the middle cerebral artery was induced as described (Yanaka et al., 1996a1996b, and 1996c). Briefly, the right carotid bifurcation was exposed through a midline skin incision of the neck. The internal carotid artery was followed rostrally, and the pterygopalatine artery was ligated. The branches of the external carotid artery were dissected and divided. The occluder, 4–0 nylon suture with a silicone-coated tip, was then advanced from the external carotid artery into the lumen of the internal carotid artery until it blocked the origin of the middle cerebral artery. The average size of the silicone-coated portion was 0.25 mm in diameter and 12.0 mm in length. We chose an occluder that blocked the middle cerebral artery at the depth of 19–20 mm. Reperfusion was accomplished by withdrawal of the suture. The animals underwent ischemia for 1 h and reperfusion for 48 h. Postoperative recovery typically occurred within 2 h after the onset of ischemia. After surgery, the rats were allowed free access to food and water. Neurological deficit characterized by left-sided hemiparesis and right Horner's syndrome were used as criteria for ischemic insult. We also excluded any animals that had subarachnoid hemorrhage or lacked neurological deficits at 3 h after the induction of ischemia.
A neurological examination was performed 12, 24, and 48 h after occlusion by a blinded observer. A standard scoring scale was used: 0, normal; 1, failure to extend the left forepaw; 2, circling to the left; 3, falling to the left; and 4, does not walk spontaneously and exhibits a consciousness disturbance (Zea Longa et al., 1989).
Laminin peptide synthesis and treatment
Three synthetic peptides were selected for this study based on their previously reported ability to interact with PMN (Harvath et al., 1994). The peptides were derived from various regions of the murine laminin α chain: peptide TG-1 from the amino-terminal top globule and peptides GD-1 and GD-6 from the carboxy-terminal globular domain (Fig. 1). The amino acid sequence and net charge for each peptide are summarized in Table 1. Peptides were synthesized and purified as described (Wilke and Skubitz, 1991; Gehlsen et al., 1992; Harvath et al., 1994). Briefly, laminin peptides were synthesized at the microchemical facility of our institution using a Beckman system 990 peptide synthesizer (Beckman Instruments, Fullerton, CA, U.S.A.). Lyophilized crude peptides were purified by preparative reverse-phase HPLC on a C-18 column and were eluted with a linear gradient of acetonitrile (0–60%) containing 0.1% trifluoroacetic acid in water. Peptide composition was verified by amino acid analysis before use. Peptide purity was >95%, based on analytical HPLC.
Laminin peptides

Each of the amino acids are demonstrated as a single letter abbreviations. G. glycine; A, alanine; V, valine; L, leucine; I, isoleucine; F, phenylalanine; M. methionine; C, cysteine; S, serine; T, threonine; K, lysine; R, arginine; D, aspartate; E, glutamate; N, asparagine; Q, glutamine; P, proline.
On the basis of the sequence reported by Sasaki et al., 1988.
Myeloperoxidase activity assay
Inflammatory cell infiltration can be quantified by using an assay for myeloperoxidase (MPO), an enzyme found within the azurophilic granules of PMN (Bradley et al., 1982). The MPO activity assay has been found to be a reproducible and objective method to reliably estimate PMN infiltration and correlates well with other estimations of PMN movement into inflamed tissues (Bradley et al., 1982; Barone et al., 1991; Matsuo et al., 1994). For the biochemical determination of MPO activity, the animals (Group I: n = 6, Group II: n = 4, Group III: n = 4, Group IV: n = 4) were anesthetized at 48 h after reperfusion and perfused transcardially with 150 ml of physiological saline (25°C at a pressure of 100 mm Hg) before brain removal to flush all blood components from the vasculature. Brain samples of the ischemic and contralateral hemispheres were taken from the middle cerebral artery area, immediately frozen in powdered dry ice, and stored at −80°C for later biochemical analysis. The MPO assay for brain tissue was conducted as described earlier (Matsuo et al., 1994): Tissue segments were thawed on ice, the cortical tissues were dissected from the subcortical portion, and the wet weight of the cortical tissue in grams was rapidly measured. Each sample was homogenized in 5 mmol/L potassium phosphate buffer (pH 6.0, 4°C, 1:20 wt/vol) using an Overhead-Stirrer (three on/off cycles at 5-s intervals; Wheaton Instruments, Millville, NJ, U.S.A.) and centrifuged at 30,000 g (30 min, 4°C; JS-H2 Centrifuge, Beckman Instruments, Palo Alto, CA, U.S.A.). The supernatant was discarded and the pellet was washed again as described. After decanting the supernatant, the pellet was extracted by suspending the material in 0.5% hexadecyltrimethylammonium bromide (Sigma Chemical Co., St. Louis, MO, U.S.A) in 50 mmol/L potassium phosphate buffer (pH 6.0, 4°C) for ∼2 min at an original tissue wet weight-to-volume ratio of 1:10. The samples were immediately frozen on powdered dry ice and subjected to three freeze-thaw cycles, after which sonication was repeated between cycles. After the last sonication, the samples were incubated at 4°C for 20 min and centrifuged at 12,500 g (15 min, 4°C). Supernatant MPO activity was assayed as described (Bradley et al., 1982). Briefly, 0.1 ml of supernatant was mixed with 2.9 ml of 50 mmol/L potassium phosphate buffer, pH 6.0, containing 0.167 mg of o-dianisidine dihydrochloride (Sigma Chemical Co.) and 0.0005% hydrogen peroxide (Sigma Chemical Co.). The rate at which a colored product formed during the MPO-dependent reaction of o-dianisidine dihydrochloride was measured. The change in absorbance at 460 nm was recorded at 15-s intervals over 2 min using a spectrophotometer (DU-64; Beckman). One unit of MPO activity is defined as that which degrades 1 µmol of peroxide per minute at 25°C (Barone et al., 1991). The MPO activity assay was carried out by a blinded observer.
Measurement of infarct size
The animals were killed at 48 h after reperfusion (Group I: n = 5, Group II: n = 7, Group III: n = 7, Group IV: n = 7) for the volume assessment. The infarct size was measured as reported previously (Yanaka et al., 1996). Briefly, the brains were removed and kept at −80°C for 5 min. Each brain was then cut into 2-mm-thick coronal blocks, for a total of seven blocks per brain. The brain slices were incubated at 37°C for 30 min in 2% 2,3,5-triphenyl-2H-tetrazolium chloride. The unstained regions, which reflect mitochondrial damage, have been shown to correspond well to regions of histopathological infarction (Bederson et al., 1986). A tight correlation between histopathology and staining results within 3 days after the onset of ischemia in rat brain has been reported (Clark et al., 1993). The surface of each slice was digitized, and the total and infarcted volumes were calculated using three-dimensional reconstruction software (Jandel PC3D, Corte Madera, CA, U.S.A.) by a blinded observer.
Statistics
All values are expressed as mean ± SD. A one-way analysis of variance was performed on data of MPO activity, size of infarction, and peripheral leukocyte counts. A two-way analysis of variance with Tukey multiple comparisons was carried out to compare the neurological outcome. Differences were considered significant if p < 0.05.
RESULTS
Myeloperoxidase activity
The mean MPO activity (units/g wet tissue) in the ischemic cortex was as follows (Fig. 2): Group I: 0.21 ± 0.04, Group II: 0.09 ± 0.03, Group III: 0.22 ± 0.03, and Group IV: 0.24 ± 0.04. Group II showed a significant decrease in the MPO activity in the lesion when compared with Group I (p < 0.001). There was no significant difference in the contralateral cortex between untreated and treated groups.
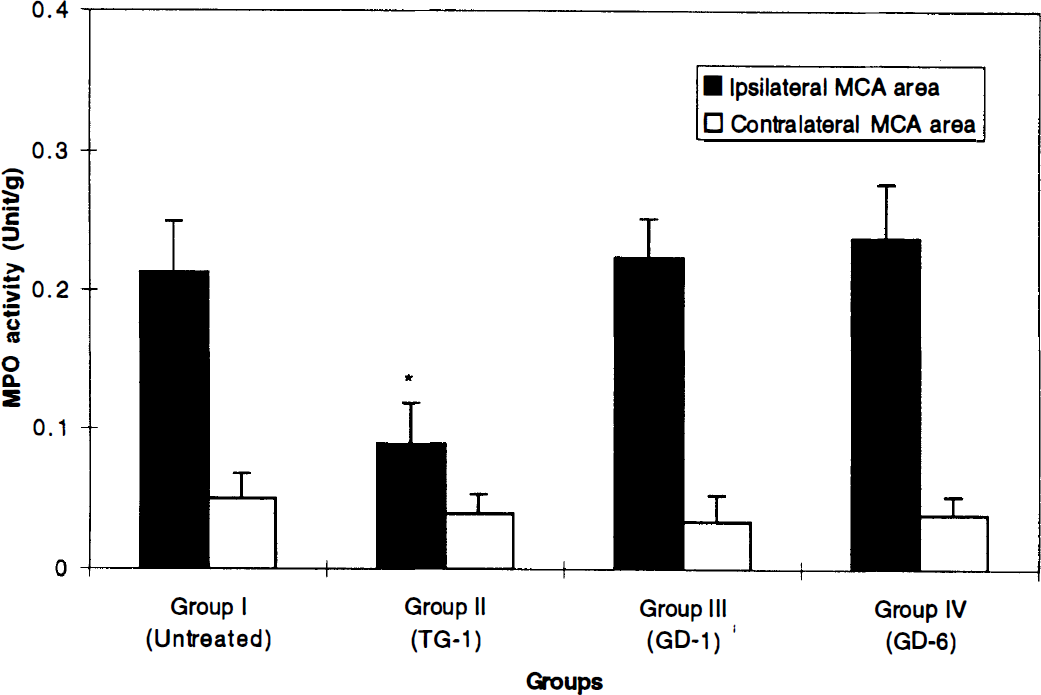
Graph showing myeloperoixidase (MPO) activity. All values are expressed as mean ± SD. *The MPO activity in the ischemic hemisphere of the Group II shows a significant decrease when compared with Group I (p < 0.001). MCA, middle cerebral artery.
Infarct volume
The mean infarct size, expressed as a percentage of the total hemispheric volume ± SD, was as follows (Fig. 3): Group I: 46.91 ± 4.51%, Group II: 31.44 ± 11.08%, Group III: 40.10 ± 9.15%, and Group IV: 40.34 ± 15.94%. Group II showed a significant decrease in the size of infarction when compared with Group I (p = 0.015).
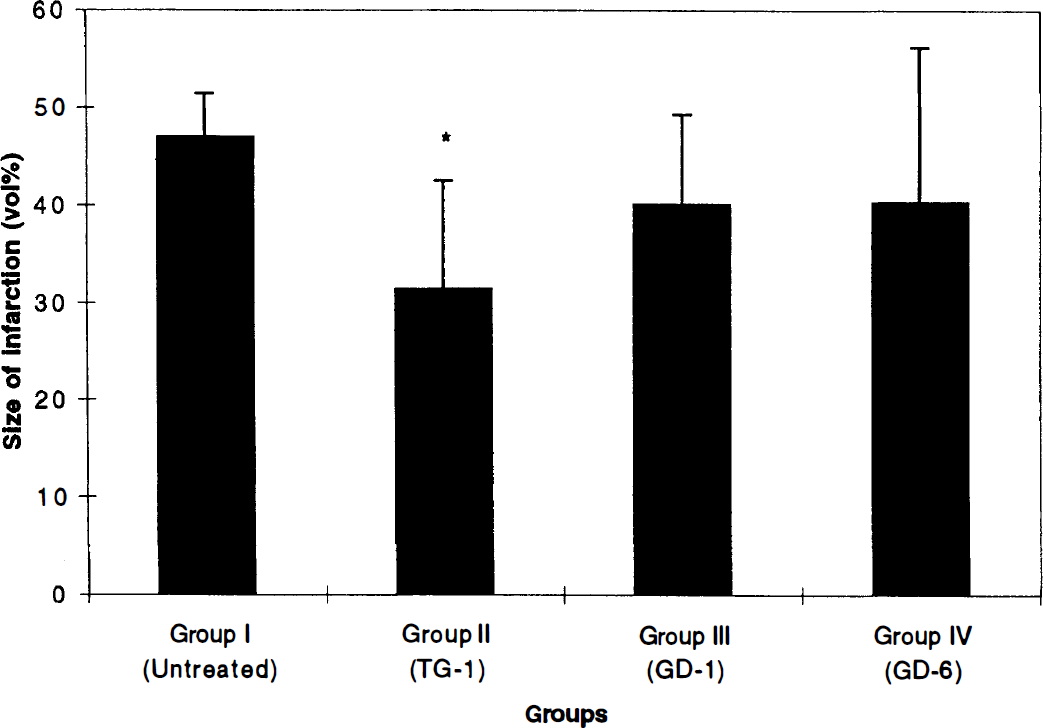
Graph showing mean infarct volume expressed as a percentage of the total hemisphere for each study group. All values are expressed as mean ± SD. Vertical bars indicate standard deviation. *Group II shows a significant decrease in the size infarction when compared with Group I (p = 0.015).
Neurological assessment
The neurological grades of the animals are summarized in Fig. 3. The neurological grade of Group II was significantly better than in Group I (p = 0.012) at 48 h after reperfusion (Fig. 4).
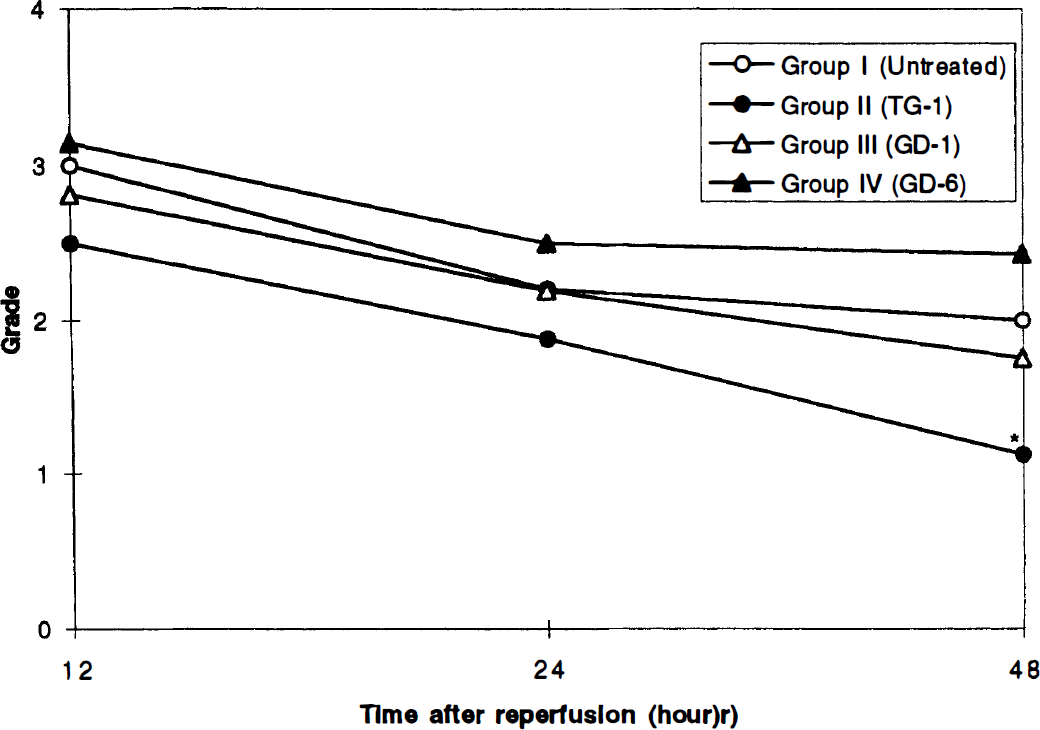
Graph showing the clinical outcome of the animals in each study group. See text for definition of grades. *The neurological grade of Group II is significantly better than in Group I (p = 0.012) at 48 hours after reperfusion.
Peripheral leukocyte counting
No significant differences were detected in the number of leukocytes and differentials in Groups I, II, and III. There was a significant reduction of the number of leukocyte and lymphocyte counts in Group IV after peptide administration (Table 2).
Peripheral white blood cell and differential values (×1,000 mL; mean ± standard deviation) before ischemia and after reperfusion.
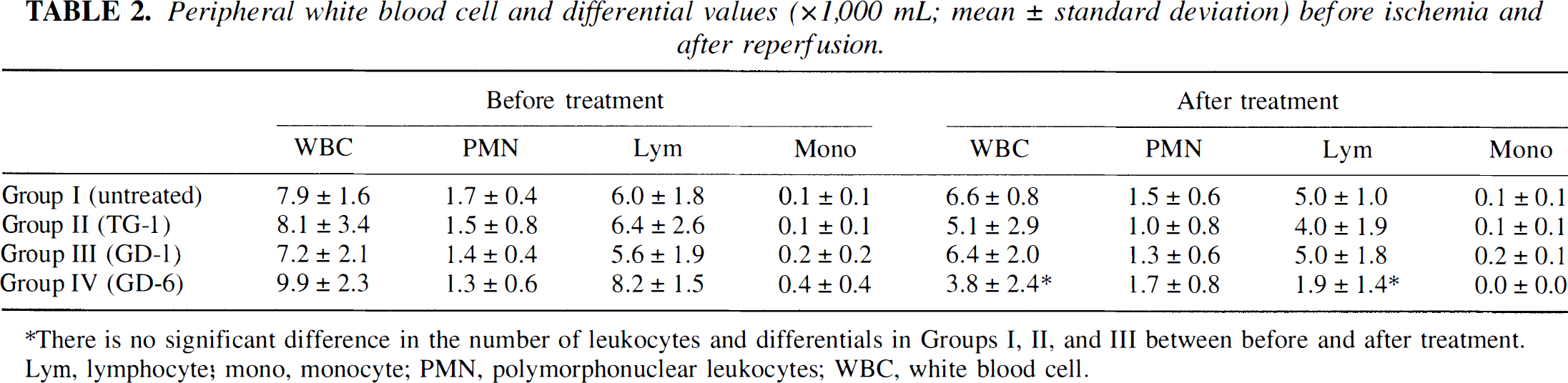
There is no significant difference in the number of leukocytes and differentials in Groups I, II, and III between before and after treatment. Lym, lymphocyte; mono, monocyte; PMN, polymorphonuclear leukocytes; WBC, white blood cell.
DISCUSSION
The current study convincingly demonstrates that intravenous administration of laminin peptide TG-1 significantly reduces leukocyte accumulation into the ischemic tissue, reduces the size of infarction, and improves neurological outcome after transient focal cerebral ischemia in rats.
Extracellular matrix proteins such as fibronectin and laminin play an important role in axonal regeneration, cerebral edema, and cerebral neoplasia (Rutka et al., 1988). Laminin binds to collagen IV, heparan sulfate proteoglycan, and to itself. It is also able to span the basement membrane and bind to various substances on the surface of cells via the interaction of membrane receptor with various other ligands (Martin and Timpl, 1987). Binding to laminin elicits cell-specific responses, causing secretory cells to become polarized, neural cells to extend axonlike processes, various cells to migrate, and a variety of cells to differentiate (Martin and Timpl, 1987). Laminin has been shown to promote or modulate PMN adhesion, motility, oxidant generation, degranulation, and phagocytosis (Suchard, 1993; Simms and D'amico, 1994). In addition, synthetic peptides derived from laminin such as TG-1 have been reported to stimulate neutrophil motility and migration (Harvath et al., 1994). Bryant et al. (1987) reported that PMN extravasation across basement membranes was aided both by reversible attachment of the cells to laminin in the matrix and by chemotaxis to a gradient of soluble intact and possibly degraded laminin. Furthermore, we have already studied peptides derived from fibronectin, and the positive results that we obtained in those studies led us to study peptides derived from laminin. It has been shown that the extracellular matrix proteins fibronectin and laminin interact with the same families of receptors, so it seems possible that if fibronectin peptides have an effect in this model system, laminin peptides may also have an effect.
The exact mechanism of action that laminin peptide TG-1 uses to cause the effect that it has in this animal model of focal cerebral ischemia has yet to be elucidated. Perhaps this peptide causes its effect by binding directly to PMN or other cells in the bloodstream. However, the cell surface molecule to which peptide TG-1 binds has not been determined. In earlier studies, we have reported that peptide TG-1 does not bind to [3H]heparin (Wilke and Skubitz, 1991), so it is unlikely that proteoglycans or glycosaminoglycans are the cell surface receptors for peptide TG-1. The two other laminin peptides used in this study, peptides GD-1 and GD-6, have previously been shown to bind [3H]heparin (Skubitz et al., 1991; Wilke and Skubitz, 1991), and neither of them had an effect in this study. We have previously published that the human melanoma cell line C8161 uses the α3β1 integrin receptor to bind to laminin peptide GD-6 (Gehlsen et al., 1992). Furthermore, we have preliminary data that the human HT-1080 fibrosarcoma cell line uses the α3β1 integrin as its cell surface molecule to bind to peptide GD-1 (L. T. Furcht, unpublished observation). Since neither peptide GD-6 nor peptide GD-1 was able to inhibit cerebral ischemia in this model, our results suggest that the α3β1 integrin receptor may not play an important role in this model system. However, it should be noted that other integrin subunits, such as α1β1, α2β1, α6β1, α7β1, α9β1, αvβ3, αIIbβ3, and α6β4, have been shown to serve as receptors for cells to adhere to laminin (Delwel and Sonnenberg, 1996). The integrin αvβ3 has been reported to be a major adhesion receptor for laminin and is expressed in cerebral microvessels after transient focal cerebral ischemia (Okada et al., 1996). Laminin interacts with β1-integrins, which were originally believed to be absent from PMN. Recent investigation has revealed β1-dependent adhesion pathway on neutrophils (Kubes et al., 1995). Therefore, it is not possible to rule out the entire family of integrins as possible cell surface receptors that may be important in this model system. Cells use many different surface molecules to adhere to laminin, in addition to proteoglycans and integrins, including a 67-kDa protein, cranin, lectins, galactosyltransferase, sulfated polysaccharides, and glycolipids (Meeham, 1991; Mercurio and Shaw, 1991). Since we do not know the receptor that cells use to adhere to peptide TG-1, it is possible that the PMN or other cells in the bloodstream use these other cell surface molecules as receptors to mediate the effects that were observed. Laminin has a variety of functional activities including promoting the adhesion, spreading, migration, and invasion of a variety of different cell types. Therefore, it is not surprising that a synthetic peptide derived from a functionally active domain of laminin has the ability to function as it did in this ischemia model.
The pharmacokinetics of the laminin peptides used in this study, specifically the half-lives of the peptides, have yet to be determined. The sequences of peptides selected for synthesis were all about the same size and of similar net charge. Furthermore, the three peptides used in this study were selected from functionally active sites of laminin (Skubitz et al., 1991; Wilke and Skubitz, 1991; Mercurio, 1995). Therefore, it seems that these peptides may have similar half-lives in vivo, although this would need to be determined. Again, these types of studies are beyond the scope of the present ischemia model study presented herein.
We are not able to provide a rationale as a decrease in the number of white blood cells was observed in those rats pretreated with peptide GD-6. Again, this peptide has been shown to use the α3β1, integrin as a receptor in certain cell lines, and it has been shown to bind [3H]heparin. Since [3H]heparin is negatively charged and peptide GD-6 is positively charged, it is possible that it binds to negatively charged constituents in the bloodstream and, in an indirect manner, has an effect to lower the white blood cell blood levels. It is difficult to hypothesize some toxic effects on PMN, since the levels of other cells in the bloodstream including lymphocytes and monocytes were not affected by peptide GD-6. Furthermore, all three peptides used in this study were positively charged, so it is unlikely that any hypothetical “toxic” effect that might be caused by peptide GD-6 could be due solely to charge alone. As stated in Methods, all three peptides were purified in exactly the same manner by HPLC and were then tested by amino acid analysis and mass spectrometry for purity. None of peptides contained impurities by these detection techniques.
Although the use of peptides as blocking agents after ischemia/reperfusion raises several interesting points, this approach also depends on the characteristics of the peptide itself, including receptor specificity and number and target organ. In addition, this peptide may produce unexpected actions since extracellular matrix proteins have been shown to participate in blood coagulation, angiogenesis, binding growth factors, and cytokines, which also can contribute to their effect on ischemic brain injury. The cell adhesion process after ischemia is still only partly understood; it cannot be excluded that the peptide may yet have unforeseen positive or negative effects. Further experiments are required to resolve these issues, and studies are currently underway to determine the mechanism of action of laminin peptides, including receptors on PMN that may be involved in interacting with the laminin peptides.
In conclusion, the data presented strongly demonstrated that the synthetic laminin peptide TG-1 was effective in brain protection following ischemic insult. Identification of binding sites in laminin peptide will help our understanding of laminin-mediated regulation of cellular adhesion process.