
Abstract
We recently mapped the visual recognition memory network in the behaving baboon using a positron emission tomography (PET) activation paradigm with 18F-fluoro-deoxyglucose during a visual delayed matching-to-sample task. This study confirmed the key role of the perirhinal cortex and documented an unexpected left-sided advantage. Specific contribution of each subdivision of the perirhinal cortex has, however, never been investigated. Furthermore, although alteration to the perirhinal cortex has been implicated in several brain disorders, putative plasticity within the entire brain network after perirhinal damage remains largely unknown. To confirm our previous data and to investigate these latter issues, we used our PET activation paradigm on a second healthy baboon before and after 16 months after bilateral excitotoxic lesions of the perirhinal cortex. Activation common to our two healthy baboons occurred only in the left rostroventral perirhinal cortex (i.e., areas 36pm and rostral 36r) and insular cortex. Although histologic analysis disclosed that the perirhinal lesions achieved in the present baboon were essentially caudal to this preoperatively activated area, memory performance was severely impaired. Concomitant with this long-lasting cognitive deficit, changes in the neural network implicated in the task were observed, involving disappearance of the preoperative activations and appearance of a significant activation of the frontal and occipital cortices. However, different activation patterns were found in the first and last eight postoperative months. These findings highlight the functional heterogeneity of the perirhinal cortex and evidence progressive plasticity after perirhinal cortex damage.
Keywords
Introduction
Brain structures underlying visual recognition memory have been extensively investigated in nonhuman primates, especially so by lesion and recording studies with the use of delayed matching-(or non-matching-) to-sample (DMS/DNMS) tasks (Mishkin and Murray, 1994). We recently developed a positron emission tomography (PET) activation paradigm in the behaving baboon with 18F-fluorodeoxyglucose (FDG) during a computerized visual DMS task that allowed to confirm the recognition memory network classically acknowledged (Blaizot et al, 2000). Within the medial temporal lobe, the rostroventral part occupied by the perirhinal cortex (Brodmann areas 35 and 36) was the most activated area, consistent with its pivotal anatomic position and central role in visual recognition memory (Meunier et al, 1993; Buffalo et al, 1999; Malkova et al, 2001; Murray and Richmond, 2001).
Several issues concerning the functional contribution of the perirhinal cortex to visual memory are, however, little known. Firstly, specific involvement of each subdivision of the perirhinal cortex, well-defined by their cytoarchitectonic features (Suzuki and Amaral, 1994; Blaizot et al, 2004), has never been investigated. Nevertheless, functional heterogeneity was suggested in both our PET activation study (see Figure 2 in Blaizot et al, 2000) and a 2DG autoradiography investigation in macaques (Davachi and Goldman-Rakic, 2001). Secondly, putative hemispheric specialization for memory function, well-documented in humans, has been little addressed in the nonhuman primate. Our previous PET study in the baboon revealed an unexpected left-sided advantage, including the perirhinal cortex, which however requires further investigation. Thirdly, the effects of perirhinal damage on functional neural networks are unknown, although it may have implications in several brain disorders. Indeed, the perirhinal cortex is one of the areas earliest and most severely affected by the neurofibrillary tangles in both normal aging and Alzheimer's disease (AD) (Braak and Braak, 1991; Delacourte et al, 1999), and damage to the rostral perirhinal cortex (i.e., the temporopolar cortex) has been implicated in anomia (Damasio et al, 1996) and semantic dementia (Hodges et al, 1992). Furthermore, we previously reported that lesions of the rhinal cortex (i.e., entorhinal and perirhinal cortices) in the baboon induced both severe memory impairment and a pattern of brain glucose hypometabolism similar to that found in AD (Meguro et al, 1999; Chavoix et al, 2002; Blaizot et al, 2002). Selective perirhinal lesions also decreased resting-state cerebral glucose consumption, the hypometabolic profile being closer to that observed in normal aging than in AD (Millien et al, 2002). Although these findings indicate that declarative memory impairment induced by perirhinal and entorhinal damage would result from functional disruption of distributed networks, the impact of perirhinal alteration on the visual recognition memory network, including cerebral reorganization and putative compensations, remains unknown.
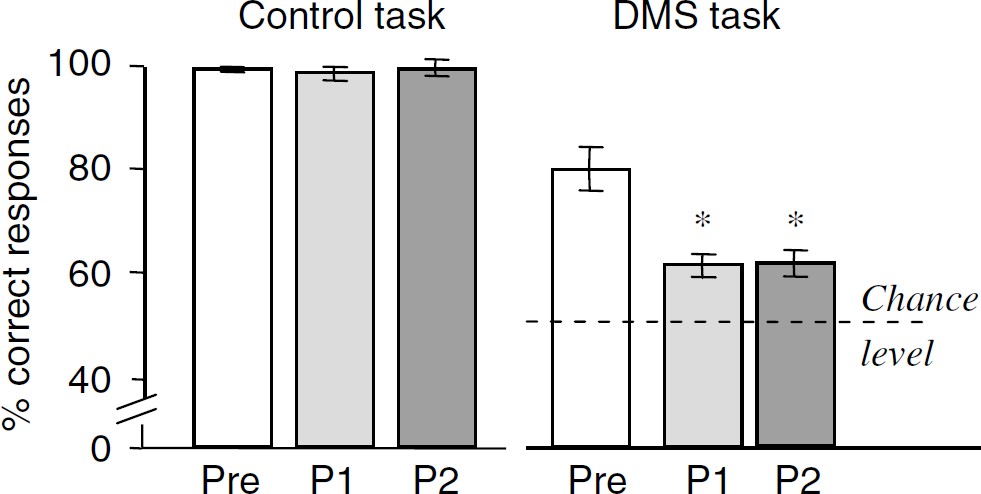
Pre-, early (P1; first 8 months) and late (P2; last 8 months) postoperative performance on the DMS and control tasks. For each task, performance is the mean ± s.d. over 6 and 8 PET sessions in the preoperative phase and each postoperative subperiod, respectively. *P<0.01 (Mann–Whitney U-test) as compared with preoperative performance.
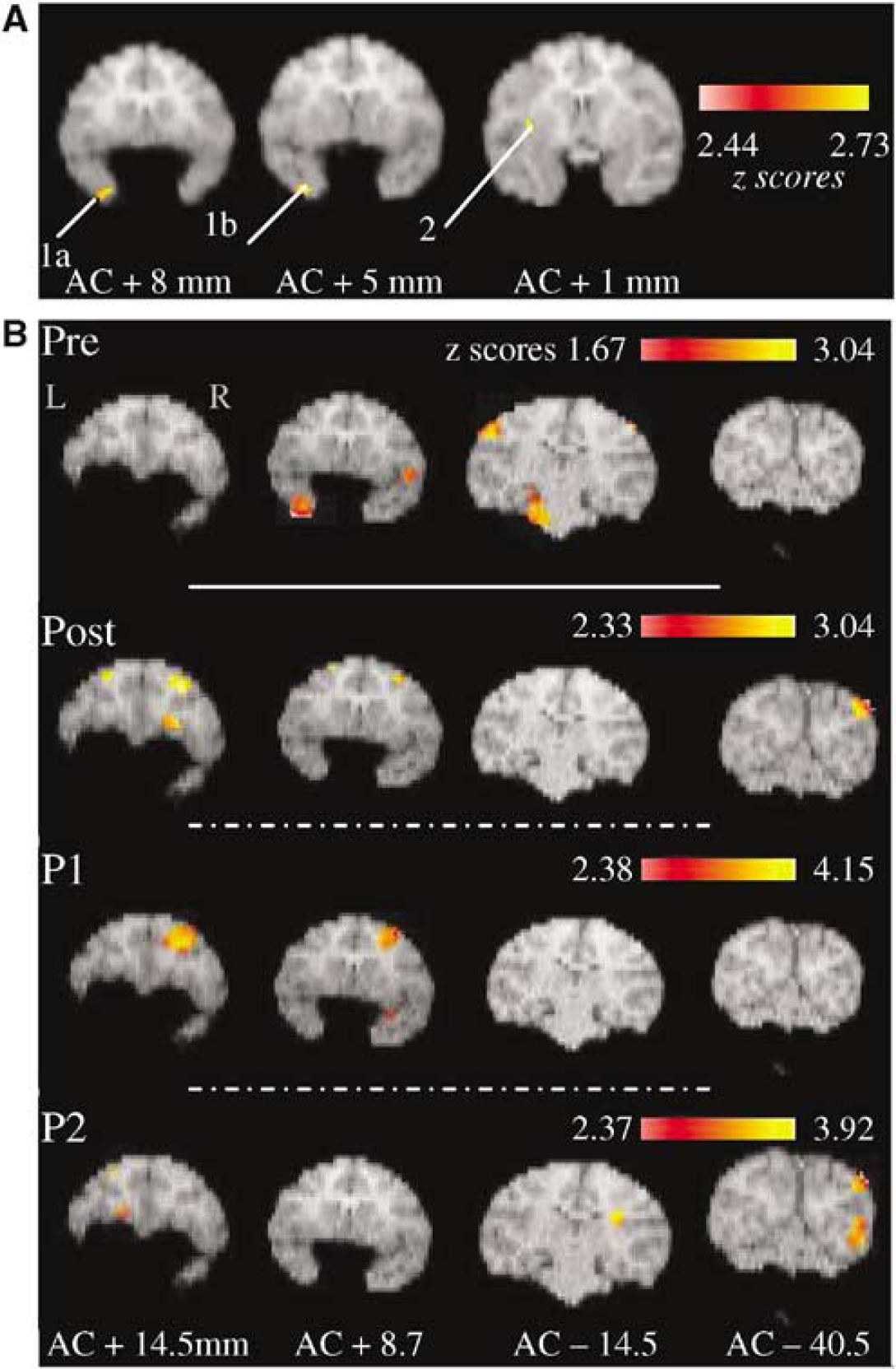
Significant activations for the tasks comparison ‘DMS minus Control’ for four illustrative coronal levels: (
The aim of the present study was therefore twofold: (1) to confirm, using our PET activation paradigm on a second healthy baboon, the functional neuroanatomy of visual recognition memory, including the issue of any preferential left-sided activation and a more detailed analysis of the medial temporal lobe activation sites by examination of postmortem histologic and cytoarchitectonic data; and (2) to investigate the functional impact of perirhinal lesions on the visual recognition memory network over an extended period of time.
Materials and methods
The experimental procedure used for this study was in accordance with the European Union rules for care and use of laboratory animals (directive 86/609/CEE). A total of 44 PET scans were acquired in a young-adult male Papio anubis baboon (3 years old, 10 kg weight), before and after bilateral lesions of the perirhinal cortex. The PET paradigm extended over 27 months, with pre- and postoperative periods lasting 10 and 16 months, respectively. Behavioral training started 1 year earlier.
Surgical and Magnetic Resonance Imaging (MRI) Procedures
Both procedures have been previously reported in detail (Blaizot et al, 1999; Millien et al, 2002). Briefly, excitotoxic lesions of the perirhinal cortex were performed bilaterally in two stages (left hemisphere 6 days after the right hemisphere) by local injections (13 target sites per side) of ibotenic acid (1.2 μl 12 to 15 μg/μl at pH 7.4 in PBS) using a methodology that combines MR techniques and stereotaxic neurosurgery.
Behavioral Testing
Testing was conducted in a sound-proof, darkened room equipped with a computerized test apparatus, as previously described (Chavoix et al, 2002). The animal faced a colour monitor fitted with a touch screen (27 × 20 cm, Sony CPD-1420E Trinitron) onto which visual stimuli were displayed, and the food reward was automatically delivered onto a cup located at the bottom right side of the screen.
The baboon was tested on the two activation tasks previously described in detail (Blaizot et al, 2000), using visual stimuli that appeared pseudorandomly in a 3 × 3 grid: a visual DMS task with trial-unique stimuli that required the baboon to memorize lists of five meaningless bicolour drawings and a visuomotor control task. For the DMS task, the five drawings of a given list were shown successively, followed by the presentation of these same drawings, each paired with a new one and a white square; the baboon was rewarded for touching the previously seen drawing. In the control task, both drawings of the choice phase were new and the white square was the rewarded choice. Before the PET study, the baboon was trained on both tasks, with stable performance over the last 5 days.
Postoperative assessment began 23 days after left perirhinal lesion. Behavioral procedure was similar to the preoperative one except that retraining on the DMS task always started with sessions requiring the memorization of single stimuli with a 1-sec delay (to a criterion >85% correct responses over 90 trials) to ensure that the animal had not forgotten the rule.
Positron Emission Tomography Procedure and Image Analysis
Positron emission tomography data of FDG brain uptake were collected according to a 63-cut three-dimensional volume axial acquisition with the high-resolution PET device ECAT HR+ (isotropic resolution [x, y, z]: 4.6 × 4.2 × 4.2 mm). The baboon underwent 12 preoperative scans (6 per task) and 32 postoperative scans (16 per task) starting 1 month postoperatively. Pre- and postoperative periods were divided into 3-week blocks, with alternation between blocks of DMS and control tasks. Each block consisted of 1 week of training (or retraining) on the proper task, followed by 2 weeks of testing during which two PET scans were acquired at approximately 7-day intervals.
The PET procedure was the same as in Blaizot et al (2000). On each PET day, the baboon was administered a bolus of approximately 2 mCi of FDG, 3 mins after the beginning of the 28-mins task. The baboon was then anesthetized (ketamine–xylazine: 6–0.6 mg/kg i.v.) and transferred to the PET device for a 10-mins PET scan and a 15-mins transmission scan, initiated 50 and 120 mins after the FDG injection, respectively.
For image analysis, a T1-weighted volumetric MR scan was acquired. Edition of the MR volume, reslicing of the volumetric PET data into coronal cuts, coronal MRI-PET coregistration, masking of the PET data, smoothing and normalization of the data for global activity were obtained as previously described (Blaizot et al, 2000).
The two questions asked in the present study were tested as follows. The functional neuroanatomy of visual recognition memory in healthy conditions was specified with a conjunction analysis between the preoperative memory and control condition data sets from the present baboon and those from another healthy baboon in which 15 PET scans were performed (8 and 7 for the DMS and control tasks, respectively) (Blaizot et al, 2000). For each animal, the PET data sets were coregistered with the subject's MRI, and an average PET image was obtained. Spatial normalization of this average image was then performed in SPM99 using the template baboon (Black et al, 2001), and parameters from this normalization were further applied to all coregistered PET–MRI data sets. Smoothing and normalizing the PET data for global activity were performed afterwards. Changes in the visual recognition memory network after perirhinal lesions were assessed by difference analyses between pre- and postoperative data (i.e., (DMS Task—Control)post-op versus (DMS Task—Control)pre-op). As this analysis was performed in one animal only, it was performed in native space (i.e., no spatial normalization). Because network plasticity could be an ongoing process for months, the postoperative period was further divided into an early (Post 1) and a late (Post 2) equal subperiods of 8 months each, each including 16 PET scans (8 per task).
Both the conjunction and the difference analyses were performed voxel by voxel by means of the SPM99 software (Welcome Department of Cognitive Neurology, London), using ‘memory minus control’ data sets for each subject and each period (i.e., pre- and postoperative) and the design ‘single-subject: conditions and covariate’ with the performance as covariate. The voxels exceeding a statistical criterion of P<0.01 (uncorrected) were superimposed on the corresponding coronal MRI planes of the baboon, and activated brain areas were anatomically identified by one of us (RI). This rather liberal significance cutoff was justified by the unique use of histology combined with coregistered MRI-PET data in the present study, which enhances the biologic significance of the results. Specific attention was paid to the parahippocampal gyrus, for which precise identification of each area was performed using both our histology-MRI comparative method (Blaizot et al, 2004) and histologic sections of the present baboon.
Histologic Procedure
At 18 months after surgery, the baboon was deeply anesthetized and the brain was sequentially perfused transcardially with saline and a series of various concentrations of paraformaldehyde as previously described (Blaizot et al, 2004). The brain was then cryoprotected, and a block of each side containing the whole perirhinal cortex was sectioned coronally at 50 μm in a sliding microtome provided with a freezing unit (Microm, Heidelberg). A one-in-five series (250 μm interval) was mounted onto gelatin-coated slides and stained with thionin. Boundaries of the perirhinal cortex were determined using cytoarchitectonic criteria recently defined for the baboon (Blaizot et al, 2004). Damage was assessed every millimetre all along the rostrocaudal extent of the perirhinal cortex, including the temporal pole (i.e., over 18 mm), on each side and for each perirhinal division (i.e., areas 35, 36pm, 36pl, 36r and 36c), both in the superficial and deep layers separately. On each histologic section, areas with neuronal loss were plotted onto corresponding camera lucida drawings, and the area of damage was expressed as a percentage of the whole surface of the perirhinal cortex on the corresponding section. The volume of damage for a given perirhinal cortex division (expressed as a percentage) was then calculated across the 18 mm. Damage in the adjacent nontargeted areas was similarly examined, but not quantified. Atrophy resulting from the neuronal loss was stereologically measured on coronal T1-weighted MR images, by comparing, for each side, the preoperative volume of the medial temporal cortex (i.e., rhinal and piriform cortices plus amygdala and hippocampus when available) with that obtained 248 days after the lesion. However, because the degree of atrophy was less than 15%, difference was too small to provide reliable data on the quantification damage.
Results
Preoperative Behavioral Performance and Neuroimaging Data
Preoperative performance of the present baboon is shown in Figure 1. As illustrated in Figure 2A, the conjunction analysis revealed common activation in the perirhinal cortex (one peak in area 36pm (Z score = 2.73, P<0.01) and another in area 36r extending to area TE (Z score = 2.44, P<0.01)) and the insular cortex (Z score = 2.45, P<0.01) only, all on the left side.
Histologic Data
Neuronal loss was of similar distribution on both sides albeit more severe on the left side (Figure 3). Area 35 was very accurately and uniformly damaged (97% and 79% damage on the left and right side, respectively), whereas damage varied along the rostrocaudal extent of area 36. Area 36p was spared as a result of the high difficulty to reach this area using our surgical approach. Area 36r was significantly damaged (58% and 27% damage on the left and right side, respectively) and area 36c only slightly (11% and 8% damage, respectively) with complete sparing of the caudal two-fifths, the medial part being more severely affected than the lateral part for both areas. Furthermore, in contrast to area 35 in which all cell layers were destroyed, only layers V and VI were consistently damaged in area 36, further extending to layer IV in caudal 36r and to all cell layers in rostral 36r. The adjacent entorhinal cortex, TE, TH and TF areas were spared with, however, minimal damage in the right area TE. Moreover, the rostral subiculum and, to a lesser extent, the rostral CA1 field of the hippocampus were slightly damaged, the right side again less so than the left.
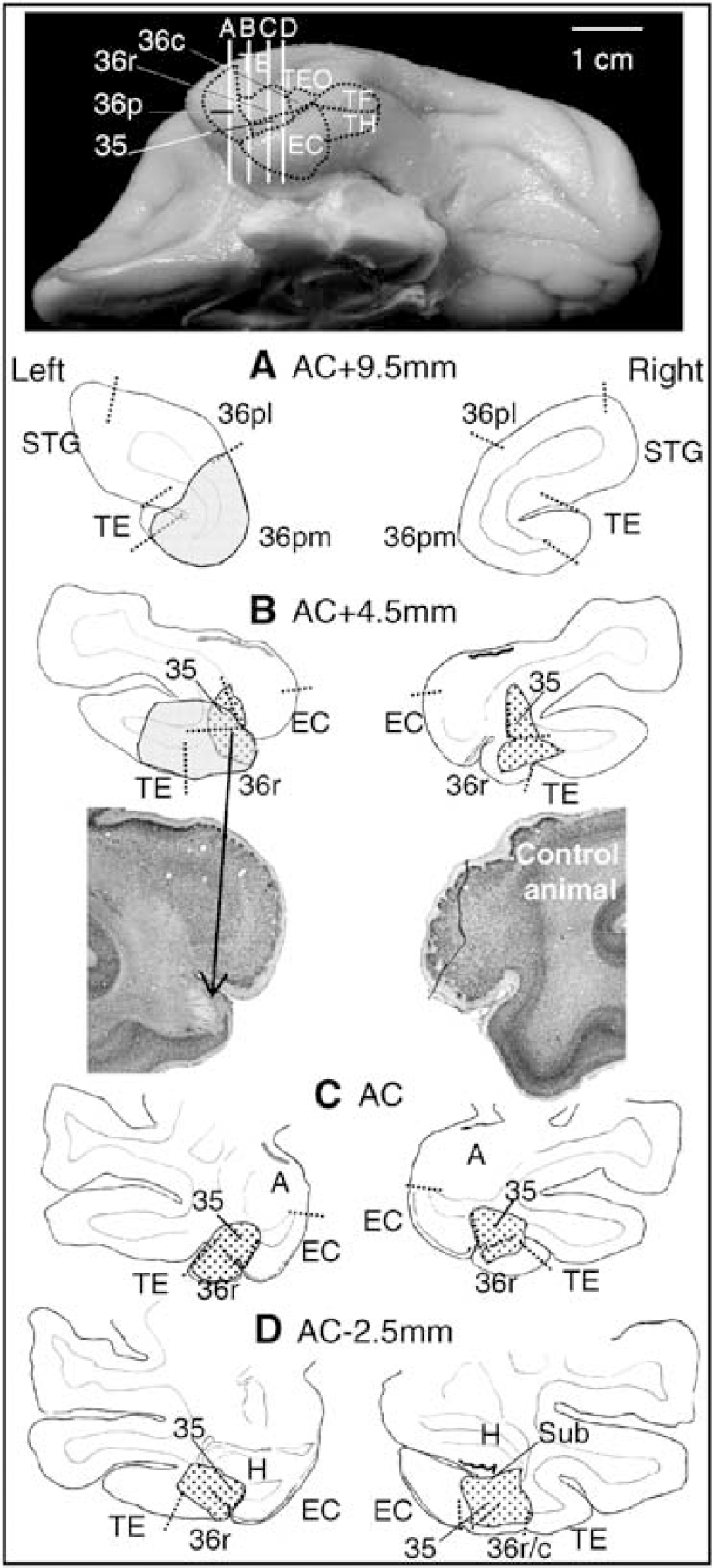
Superimposition of histologie damage and significant preoperative perirhinal activations of the present baboon. Top: location of the left perirhinal cortex and its subdivisions (areas 35, 36pm, 36r and 36c) on a ventral view of a baboon's brain. The vertical lines correspond to the four levels (
Postoperative Behavioral Performance
The animal relearned the DMS task almost immediately. However, postoperative DMS performance was significantly impaired as compared with preoperative one (59% ± 4% versus 77% ± 6% correct responses; P<0.01), whereas control performance remained normal (98% ± 2% versus 99% ± 1% correct responses). Delayed matching-to-sample performance was significantly impaired (P<0.01) at each postoperative subperiod and there was no significant difference between Post 1 and Post 2 performance in any of the tasks considered (Figure 1).
Postoperative Neuroimaging Data
Brain areas significantly activated for the tasks comparison (DMS—Control) before and after perirhinal lesion in the present baboon are illustrated in Figure 2B. Preoperatively, significant activation was found in bilateral insular cortex, left area 36pm of the perirhinal cortex (also includes areas 36r, 35 and TE; see Figure 3 for details of these perirhinal activations as superimposed with histologic damage), posterior hypothalamus (including the mammillary bodies), pontine nuclei and cerebellum as well as in right retrosplenial cortex, inferior parietal gyrus (somatosensory area 3b) and mesodiencephalic junction. By contrast, significant postoperative activations were right-sided or bilateral, and involved the frontal and occipital cortices only. Interestingly, separate analyses at the early and late postoperative subperiods revealed significantly different patterns of network activation over time. These differences revealed by the tasks comparison led us to further justified investigating the changes that occurred after perirhinal lesion by means of a direct comparison to the preoperative period (i.e., the difference analysis, see Materials and methods). Significantly higher activation in the postoperative period (and each subperiod) as compared with the preoperative one are listed in Table 1.
Higher activations after perirhinal lesion (i.e., postoperatively and at each postoperative period) as compared to preoperatively
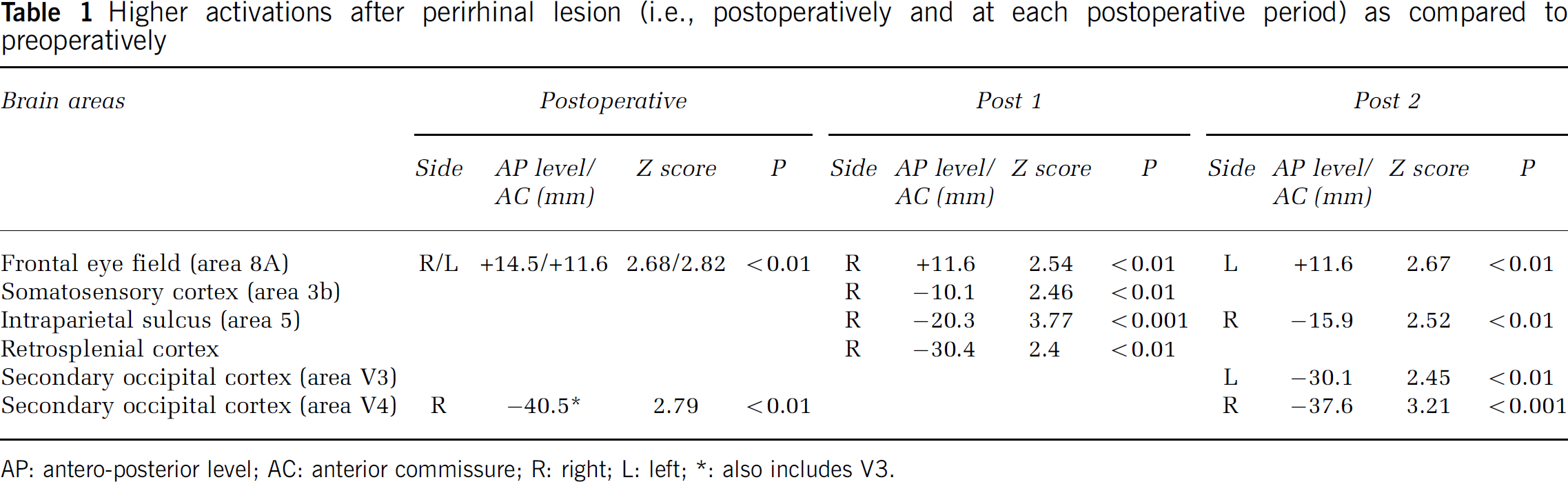
AP: antero-posterior level; AC: anterior commissure; R: right; L left;
also includes V3.
Discussion
The present findings give new insights into the neural network involved in a visual DMS task, especially regarding the functional heterogeneity of the perirhinal cortex, and provide the first account on changes occurring in the entire brain after selective lesions of this ventromedial temporal area, suggestive of plasticity in the neural network subserving visual recognition.
Concerning brain mapping in healthy baboons, common activation occurred in the perirhinal and insular cortices only. The former was the most significantly activated, in agreement with its key role in visual declarative memory (Murray and Richmond, 2001). By contrast, the finding concerning the insular cortex was rather unexpected. This somatic processing area is the final link between the sensory pathway and the limbic system, and has been considered analogous to area TE for tactile functions (Schneider et al, 1993). Activation in the anterior part of the insular cortex that mainly responds to stimulation of the face and intraoral structures (Schneider et al, 1993) probably reflects somatosensory processes related to the food reward. Interestingly, the insular cortex has significant interconnections with the perirhinal and entorhinal cortices (Mesulam and Mufson, 1982; Insausti et al, 1987; Suzuki and Amaral, 1994), which suggests high functional relevance of this neuroanatomic network for the DMS task.
Regarding the perirhinal cortex, our PET data in healthy baboons combined with histologic data evidenced two new findings: the functional heterogeneity of the perirhinal cortex subdivisions and the different implication of both sides. Within the perirhinal cortex, significant common activation was confined to areas 36pm and rostral 36r, that is, in the rostroventral-most perirhinal cortex, which corresponds mainly to the ventral part of area TG of von Bonin and Bailey (1947) and Brodmann's area 38 (Blaizot et al, 2004). This is consistent with other findings concerning visual recognition memory in monkeys: the implication of the ventral temporal pole (Horel et al, 1987), the predominant role of the rostral perirhinal cortex recently suggested (Malkova et al, 2001), and previous observations in activation studies (Blaizot et al, 2000; Davachi and Goldman-Rakic, 2001). However, to our knowledge, the present study is the first to clearly demonstrate that, within the perirhinal cortex, the rostroventral part is the most heavily engaged in visual recognition memory, probably because of its preferential reciprocal connections with both the caudalmost perirhinal cortex and anteroventral part of the visual association area TE (Moran et al, 1987; Kondo et al, 2003; Lavenex et al, 2004).
Comparison between damage and preoperative activation locations in the present baboon gives further insights into the functional role of perirhinal cortex subdivisions. Despite slight overlap only between both locations (in the rostral areas 36r and 35), the memory performance dropped dramatically as classically found when damage is mostly restricted to area 35 and caudomedial part of area 36 (Meunier et al, 1993; Buffalo et al, 1999; Malkova et al, 2001). Thus, severe memory impairment can occur despite substantial sparing of the brain area appearing as the most crucial for visual recognition memory. This could be explained by the neuronal pathway involved in visual memory processing. Area 36 receives strong input from area TE and projects to area 35 (Suzuki and Amaral, 1994). Since the deep layers that were more affected than the superficial ones are the main origin for feedback projections, it could be assumed that information processing was stuck within area 36 after damage to area 35, so that encoding was no longer possible. Although there exists direct projections from the entire rostrocaudal extent of area 36 to the entorhinal cortex (Insausti et al, 1987; Suzuki and Amaral, 1994) and reciprocal connections between perirhinal cortex and the hippocampus (Van Hoesen et al, 1979; Insausti and Muñoz, 2001), these connections do not seem sufficient to allow the transfer of visual information. In the light of our findings, we may hypothesize that rostral area 35 plays a prominent role in the encoding of visual information while rostroventral area 36 could be more crucial for visual associative memory and retrieval processes.
Although our hypotheses should be confirmed by further investigations on the functional role of each subdivision of the perirhinal cortex, several lines of evidence are consistent with the major role of the rostroventral perirhinal cortex in visual associative memory. Our DMS task performed on a touch-sensitive screen involves visual-reward and putative visuo-tactile associations, two forms of associative memory in which the perirhinal cortex has been implicated (Murray and Richmond, 2001). Moreover, impairment in visuo-tactile memory after perirhinal ablation has been observed despite sparing of its caudal half part (Goulet and Murray, 2001), and damage to the temporopolar cortex has been involved in brain disorders in which the ability to relate visual representation with the corresponding words is impaired such as anomia (Damasio et al, 1996), semantic dementia (Hodges et al, 1992) and AD (Arnold et al, 1994). Our hypothesis is further supported by the pivotal anatomic position of the rostroventral perirhinal cortex. This region is not only the end station of the ventral visual pathway, but also provides a privileged site for convergence of sensory and limbic inputs (Pandya and Kuypers, 1969; Suzuki and Amaral, 1994; Insausti and Muñoz, 2001; Lavenex et al, 2004) with predominance of the visual inputs in the ventral portion of the temporal pole and convergence of olfactory and visual inputs in its medioventral part (Moran et al, 1987).
Besides highlighting the heterogeneity of the perirhinal cortex, the present study confirms the greater left-sided activation during a visual DMS task we previously detected (Blaizot et al, 2000), which could indicate left predominance for visual declarative memory in old word monkeys. In humans, the left medial temporal lobe (even more the left temporopolar cortex) is both predominantly affected in anomia and semantic dementia (Hodges et al, 1992; Damasio et al, 1996) and preferentially activated during episodic encoding (Schacter and Wagner, 1999; Strange et al, 2002; Davachi et al, 2003) and naming (Damasio et al, 1996) in healthy subjects. Thus, the preferential left-sided activation in healthy baboons performing a visual DMS task, a fact that fits with a more pronounced lesion on the left side, stresses the need for functional comparison between both sides in the nonhuman primate, which has never been done so far.
Altogether, the above data indicate that interaction of the rostroventral perirhinal cortex with both area TE and the insula would be essential for visual recognition memory and visual stimulus-stimulus association memory involved in our DMS task. It is also possible that old world monkeys use a semantic system analogous to that of humans, with some representation of the visual stimuli to solve the task. This inference from humans is supported by the cytoarchitectonic organization of the temporal pole in baboons that is more developed than in macaques and closer to humans (Blaizot et al, 2004). If this is true, then it could be speculated that the left rostroventral part of the perirhinal cortex would play a key role in semantic memory. Further investigations are, nonetheless, needed to address this issue.
Before discussing the changes in the neural network involved in the DMS task that occurred after perirhinal lesions, it should be stressed that, because observed in a single animal and with a significant threshold that could be considered to lie on the liberal side, our results are mainly descriptive and apply only for the present animal with its own cognitive strategies, so that generalization is not warranted at this stage. This drawback, inherent to the complexity and duration of the FDG PET activation paradigm in nonhuman primates, is however largely compensated for by the high number of PET scans obtained in the same subject. Furthermore, combining histology with high-resolution coregistered MRI-PET data and working in native space allows precise localization of the activated areas, probably close to that obtained with 2DG autoradiography and considerably better than that obtained with functional imaging in groups of humans, which requires spatial normalization to a brain template.
The postoperative period was characterized by greater activation of the bilateral frontal eye field and right visual area V4 than in the preoperative period. This cerebral reorganization concomitant with the memory impairment could be related to increased involvement of visuospatial processes. However, these results tell only part of the story since subdividing the 16-month postoperative period revealed time-dependence of these changes. During the early postoperative period, the retrosplenial cortex and area 8a, involved in declarative memory and/or working memory (Valenstein et al, 1987; Chafee and Goldman-Rakic, 1998; Kobayashi and Amaral, 2003), showed increased activation. Higher load was also placed on visuospatial attention and sensorimotor processes as suggested by stronger activation of the intraparietal sulcus, area 8a and area 3b. Interestingly, these cognitive areas are connected only weakly with the perirhinal cortex but more strongly so with the parahippocampal cortex (Blatt et al, 2003) known to have very dense connections with both the perirhinal cortex and the hippocampal formation (Insausti et al, 1987) and to be engaged in a visuospatial memory network. Additional compensation might have occurred from recruitment of the right hemisphere, which contrasts with the preoperative left-sided activations. This finding is reminiscent of the reduction in hemispheric lateralization with age (Cabeza, 2002) and AD (Grady et al, 2003) as well as of the preserved verbal encoding associated with reorganization of function to the right medial temporal lobe in left medial temporal pathology (Richardson et al, 2003). Higher activations in the late postoperative period were found in less cognitive areas, namely in the anterior part of the right intraparietal sulcus implicated in eye–hand coordination (Pierrot-Deseilligny et al, 2004) and in the bilateral secondary occipital cortex known to engage visual processing. These changes in the activated network from early to late postoperative periods could reflect modifications in the cognitive strategies implemented to solve the task after ineffective compensation in the early postoperative period. Alternatively, it might result from successive modifications in the available neural networks, which would be in agreement with sequential changes in the pattern of resting-state hypometabolism reported after perirhinal lesions (Millien et al, 2002). Neuroanatomy studies of the remaining perirhinal connections and putative sprouting in the adjacent hippocampal and parahippocampal regions, as performed after entorhinal cortex ablation (Leonard et al, 1995), would be very useful to better understand this cerebral reorganization.
Reorganization of the memory networks with increased recruitment of the dorsolateral prefrontal and posterior parietal cortices, and sometimes of the retrosplenial cortex, has also been reported in aging (Cabeza, 2002), AD (Grady et al, 2003) and temporal epilepsy (Dupont et al, 2002). The perirhinal cortex alteration found in these subjects could, thus, be highly involved in their memory impairments. However, the distinct activation patterns over time evidenced in our study highlights the difficulty in interpreting some human data while individual subjects are at different stages from the initial brain damage. Finally, as illustrated by the present study, combined use of lesions and functional neuroimaging in the nonhuman primate should now afford inestimable tools to investigate the functional impact of focal lesions on the neural network involved in a given task, including cerebral reorganization and putative compensations.
Footnotes
Acknowledgements
The authors thank B Landeau for her precious contribution in the PET data analysis, I Millien, F Mézenge, A Brocquehaye, MH Noel, B Haelewyn and A Young for their technical assistance regarding surgery, behavioral and PET experiments, E Artacho-Pérula for help in stereological analysis as well as the Cyceron's cyclotron team.