
Abstract
The heme oxygenase (HO) enzymes catalyze the rate-limiting step in the breakdown of heme to iron, carbon monoxide, and biliverdin. A prior cell culture study demonstrated that deletion of HO-2, the isoform constitutively expressed in neurons, attenuated hemoglobin (Hb) neurotoxicity. The present study tested the hypothesis that HO-2 gene deletion is cytoprotective in a model of Hb toxicity in vivo. Stereotactic injection of 6 μL stroma-free Hb (SFHb) into the striatum significantly increased protein oxidation in wild-type mice at 24 to 72 h, as detected by an assay for carbonyl groups. At 72 h, carbonylation was increased 2.5-fold compared with that in the contralateral striatum. In HO-2 knockout mice, protein oxidation was not increased at 24 h, and was increased by only 1.7-fold at 72 h. Similarly, striatal lipid peroxidation, as detected by the malondialdehyde assay, was significantly greater in the SFHb-injected striata of wild-type mice than in knockout mice. Striatal cell viability, determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay, was 45.0%±6.3% of that in contralateral striata in wild-type mice at 72 h; it was increased to 85%±8% in knockouts. Heme oxygenase-2 gene deletion did not alter weight loss or mortality after SFHb injection. Baseline striatal HO-1 expression was similar in knockout and wild-type mice; induction after SFHb injection occurred more rapidly in the latter. These results suggest that HO-2 gene deletion protects striatal cells from the oxidative toxicity of Hb in vivo. Pharmacologic or genetic strategies that target HO-2 may be beneficial after central nervous system hemorrhage, and warrant further investigation.
Introduction
Central nervous system (CNS) hemorrhage is the primary event in approximately 15% of strokes, and is associated with most traumatic injuries requiring hospitalization. Although its deleterious effect has often been attributed to compressive ischemia produced by a hematoma (Bullock et al, 1988; Nath et al, 1986), a growing body of evidence suggests that blood components may directly injure surrounding tissue and thereby worsen outcome (Qureshi et al, 2001). Early edema formation appears to be mediated in part by the proteolytic activity of thrombin (Xi et al, 1998); hemoglobin (Hb) and its metabolites have a later cytotoxic effect, at least in rodents (Huang et al, 2002).
In prior cell culture studies, Hb has been shown to be an oxidative neurotoxin (Regan and Panter, 1993; Wang et al, 2002). This effect is at least partly due to iron release from heme, because it was prevented with very high efficacy by iron chelators (Regan and Rogers, 2003). Heme degradation leading to iron release is catalyzed by the heme oxygenase (HO) enzymes (Abraham et al, 1996), and also occurs by reaction with peroxides and other reactive oxygen species (ROS) (Gutteridge, 1986; Nagababu and Rifkind, 2004). The former process appears to predominate in heme-mediated injuries to cultured neurons. Targeted deletion of the HO-2 gene, the isoform that is constitutively expressed in neurons (Ewing and Maines, 1997), attenuated ROS formation, protein oxidation, and cell death due to Hb or hemin (Regan et al, 2004; Rogers et al, 2003). These observations are consistent with reports that HO inhibitors attenuated edema and neuronal death after injection of whole blood or Hb into the rat striatum (Huang et al, 2002), rabbit thalamus (Koeppen et al, 2004), or pig frontal lobe (Wagner et al, 2000).
Although pharmacologic inhibition of HO has provided key information about its proxidant and antioxidant effects, currently available inhibitors have nonspecific effects that complicate data interpretation. Protoporphyrin derivatives are highly reactive compounds that alter the activity of a variety of cell enzymes and membrane channels (Grundemar and Ny, 1997; Linden et al, 1993; Luo and Vincent, 1994; Meffert et al, 1994). Some may also have an antioxidant effect that is unrelated to HO inhibition (Wagner and Dwyer, 2004). To further delineate the role of HO in hemorrhagic injury, we have developed an in vivo model that facilitates the quantitative investigation of Hb toxicity in the mouse striatum. In the present study, we tested the hypothesis that mice lacking the HO-2 gene are less vulnerable to Hb.
Materials and methods
Animal breeding and housing
All mice were obtained from our breeding colony and were maintained under standard conditions with a 12 h light/dark cycle; they were provided with unlimited access to food and water. Heme oxygenase-2 deficient mice with a B6/129 background were descended from those generated by Poss et al (1995), as previously described. Both the HO-2 knockouts and the wild-type controls used in all experiments were the first- or second-generation offspring of heterozygous breeders. Animal care and treatments were in accordance with guidelines as described in ‘Principles of Laboratory Animal Care’ (NIH publication No. 80–23, revised 1996) and approved by the Institutional Animal Care and Use Committee. Genotype was determined by polymerase chain reaction (PCR) using genomic DNA isolated from tail clippings and previously published primers (Rogers et al, 2003).
Preparation of Murine Stroma-Free Hb
After mice were euthanized with isoflurane, blood was withdrawn by cardiac puncture using aseptic techniques. After centrifugation (2500 r.p.m.) for 5 mins at 4°C, the supernatant was removed and the cell pellet was washed three times with sterile saline. Cells were then collected, suspended in sterile saline, and lysed by two freeze—thaw cycles. The sample was then centrifuged, the supernatant was removed, and Hb concentration was determined spectrophotometrically using the method of Winterbourn (1990). Stroma-free Hb (SFHb) was then diluted with sterile saline to 2 mmol/L (expressed as the concentration of the Hb tetramer), which approximates its concentration in whole blood. It was aliquoted and stored at −70°C until used.
Hemoglobin Injection
Mice were anesthetized with 2% isoflurane in a mixture of 100% oxygen delivered by face mask. They were then placed into a stereotactic frame (David Kopf Instruments). A 30-gauge stainless-steel needle was introduced through a burr hole into the right striatum at the following coordinates relative to bregma: 2 mm lateral, 1 mm anterior, 3.5 mm below the surface of the skull. Each mouse received 6 μL of 2 mmol/L SFHb injected over a period of 30 mins with a microinfusion apparatus (1 μL per 5 mins). The injection needle was slowly withdrawn 15 mins later, and the wound was sutured. After recovery, food and water were provided ad libitum. Control mice were subjected to the same surgical procedure, but were injected with an equal volume of sterile saline instead of SFHb.
Assessment of Cell Injury
In initial experiments, brains were removed 72 h after SFHb injection, sectioned, and stained with 1% 2,3,5-triphenyl tetrazolium chloride (TTC), which is reduced by viable cells to a red formazan product (Lundy et al, 1986). However, in this model an area of nonstaining tissue was often surrounded by an irregular area of tissue with visible but reduced staining; lesion boundaries could not be precisely defined. To quantify tissue injury more accurately, we modified the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay, which has been used for many years to assess the viability and proliferation of cultured cells (Carmichael et al, 1987). We made use of the fact that CNS tissue can be dissociated into individual cells or small groups of cells by gentle trituration, without loss of cell viability as determined by trypan blue exclusion or the ability to reduce tetrazolium salts to formazan (Rose et al, 1993). Dissociation allows a uniform concentration of tetrazolium to be delivered to all cells.
At 24 or 72 h after SFHb injection, mice were again anesthetized with isoflurane and were then killed by cervical dislocation. Brains were rapidly removed and placed into a 60 mm culture dish containing 3 mL Hank's balanced salt solution (HBSS). Injected and contralateral striata were excised under a dissecting microscope, minced, and placed into separate centrifuge tubes containing 1 mL HBSS. Tissue was then dissociated by gentle trituration through a Pasteur pipette, followed by passage through another pipette with narrowed tip. 1 mL of 0.25 mg/mL MTT in Dulbecco's mimimal essential medium (DMEM) was added to cells, resulting in a final concentration of 0.125 mg/mL MTT. The cell suspension was then placed in a 37°C water bath for 4 mins. Cells were then collected by low-speed centrifugation for 2 mins, and the supernatant was removed and discarded. Formazan was then extracted from cells by adding 2 mL isopropanol containing 0.04 N HCl, and vortexing. After centrifugation to remove cellular debris, absorbance of the supernatant was determined at 562 nm with a reference wavelength of 650 nm. The formazan signal in the injected striatum was expressed as a percentage of that in the contralateral striatum.
Protein Oxidation Assay
At the end of the SFHb exposure interval, injected and contralateral striata were rapidly removed and were homogenized in ice-cold cell lysis buffer containing 210 mmol/L mannitol, 70 mmol/L sucrose, 5 mmol/L HEPES, 1 mmol/L EDTA, 0.1% sodium dodecyl sulfate (SDS). After sonication and centrifugation, the protein concentration was determined (Pierce BCA protein assay kit). Proteins were denatured in 12% SDS, and carbonyl groups were derivatized to 2,4-dinitrophenylhydrazone (DNP-hydrazone) by reaction with 2,4-dinitrophenyl-hydrazine, using the Oxyblot™ kit (Chemicon, Inc., Temecula, CA, USA) and following the manufacturer's instructions. Proteins were then separated on a 12% polyacrylamide gel and were transferred to a polyvinylidene difluoride Imobilon-P transfer membrane filter (Millipore, Billerica, MA, USA) using a semidry transfer apparatus (Bio-Rad, Hercules, CA, USA). Carbonylated proteins were detected with rabbit anti-DNP (1:150, Chemicon) followed by goat anti-rabbit IgG (1:300). Immunoreactive proteins were visualized using Super Signal West Femto Reagent (Pierce, Rockford, IL, USA) and Kodak ImageStation 400.
Malondialdehyde Assay
Three days after SFHb injection, injected and contralateral striata were removed, dissociated in 5% trichloroacetic acid, homogenized, and sonicated on ice. After centrifugation, the supernatant was collected, and a thiobarbituric acid/acetate solution was added to give a final concentration of 0.3% (pH 3.5). Tubes were placed into a boiling water bath for 15 mins, and then were cooled to room temperature. Fluorescence was determined with a fluorescence spectrometer (Perkin-Elmer) with an excitation wavelength of 515 nm, an emission wavelength of 553 nm, and a slit width of 5. Fluorescence intensity was compared with that of a malondialdehyde standard curve.
Cell Counts
Animals were deeply anesthetized with pentobarbital (75 mg/kg), and then were perfused with 4% paraformaldehyde in 0.1 mol/L PBS (pH 7.4). After postfixation in the same solution at 4°C for 48 h, brains were embedded in paraffin. They were then cut coronally through the needle entry site (identifiable on the brain surface), as well as 2 mm anterior and 2 mm posterior to that plane. Sections (5 μm) were deparaffinized with xylene and graded alcohol and then stained with hematoxylin and eosin. Sections were examined by light microscopy (BX 51, Olympus); the needle track and injection site were visualized, and 200× images were captured. Injured neurons were identified by their condensed and irregular nuclei and/or eosinophilic cytoplasm. Morphologically normal neurons were quantified in a reactive zone that was 360 to 600 μm from the injection site; neurons in the corresponding area of the contralateral striatum were also counted.
HO-1 and HO-2 Immunoblotting
Striata were dissected free of other tissue, dissociated, and sonicated in ice-cold lysis buffer as described above. Proteins (15 μg/sample) were separated on a 15% polyacrylamide gel and were transferred onto a polyvinylidene difluoride Immobilon-P transfer membrane filter (Millipore) using a semidry transfer apparatus (Bio-Rad). Nonspecific sites were blocked by incubating with 5% nonfat dry milk in buffer containing 20 mmol/L Tris, 500 mmol/L NaCl, and 0.1% Tween 20 (pH 7.5) for 1 h at 37°C. Membranes were incubated at 4°C overnight with a 1:1,000 dilution of rabbit anti-HO-1 or anti-HO-2 (Stressgen Biotechnologies, Victoria, BC, Canada), followed by horseradish peroxidase-conjugated goat anti-rabbit IgG at 1:2,000 dilution at 37°C for 1 h. Immunoreactive proteins were visualized as described above.
Statistical Analysis
Data were analyzed with one-way analysis of variance. Differences between groups were determined with the Bonferroni multiple comparisons test. Significance was assigned to a P-value <0.05.
Results
Mice used for all experiments were 3 to 6 months old. Genotype was determined by PCR at 3 to 5 weeks; the knockout PCR product is 90 base pairs larger than the wild type, and migrated separately on a 2% agarose gel (Figure 1).
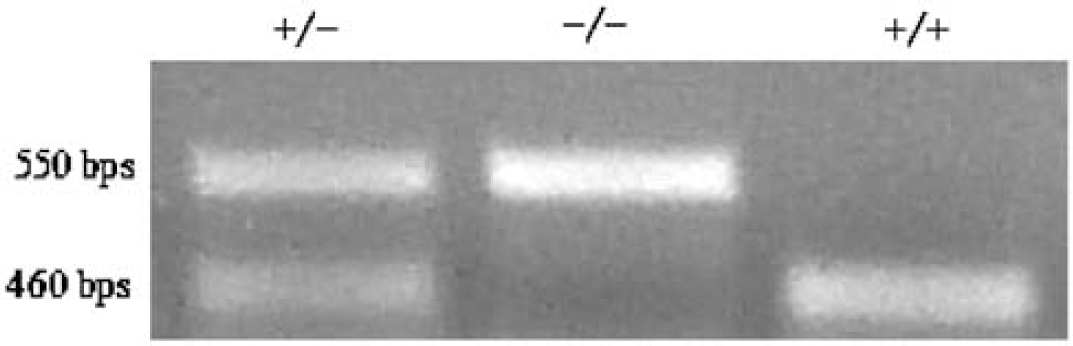
Representative 2% agarose gel, showing separate migration of wild-type (+) and HO-2 knockout (−) PCR products.
Preinjection mean weights were not significantly different between groups (Table 1). Striatal injection of 6 μL SFHb resulted in significant weight loss in both wild-type and knockout animals by 3 days. No significant weight loss was observed in control animals injected with 6 μL sterile saline. Four of 47 (8.5%) wild-type animals and 3 of 47 (6.4%) knockout animals died within 3 days of SFHb injection (P = 0.70). Large parenchymal hemorrhages, which are easily distinguished from injected SFHb by clot formation and bright red color, were seen at autopsy in each. No deaths occurred in control animals injected with saline.
Mean weight (±S.E.M., n = 12 to 37/condition) of mice immediately before striatal injection of 6 μL stroma-free Hb solution or saline, and 3 days later
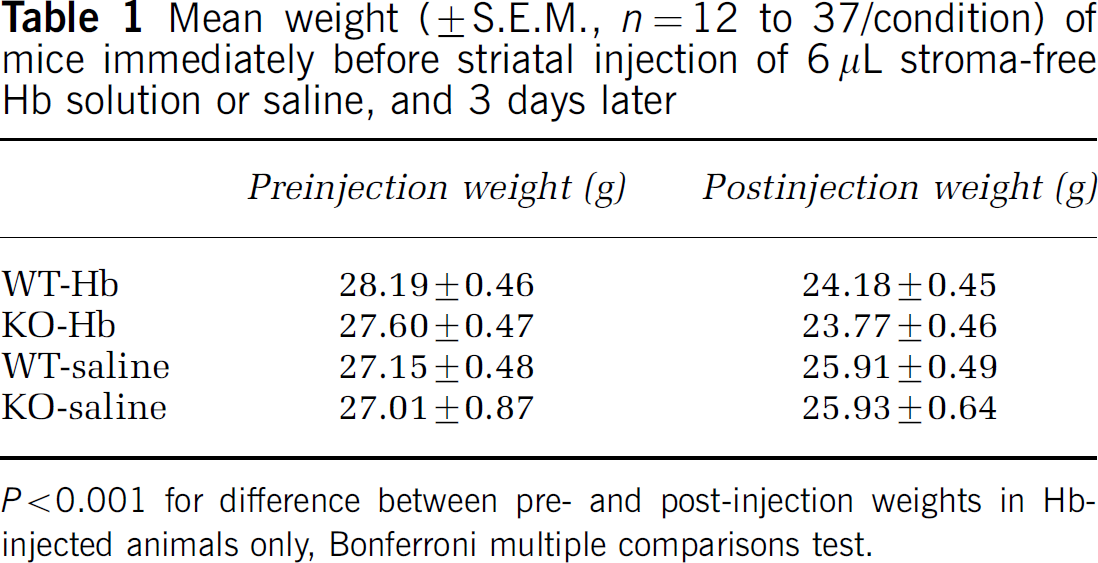
P <0.001 for difference between pre- and post-injection weights in Hb-injected animals only, Bonferroni multiple comparisons test.
HO-2 Gene Deletion Reduces Cell Death After SFHb Injection
In initial experiments, brains were removed 72 h after SFHb injection, sectioned, and stained with TTC (Figure 2), which is reduced by viable cells to a red formazan product. This produced areas completely devoid of staining and areas with clearly visible but attenuated staining. Because lesions boundaries could not be delineated, injury was quantified by cell dissociation and the MTT assay. In mice injected with 6 μL saline, the mean ratio of formazan absorbance in the injected striatum compared with the contralateral side 72 h later was 0.98 ± 0.06 (Figure 3). This was reduced to 0.45 ± 0.06 in wild-type mice injected with 6 μL SFHb. In contrast, the ratio in HO-2 knockout mice injected with SFHb was 0.85 ± 0.08.
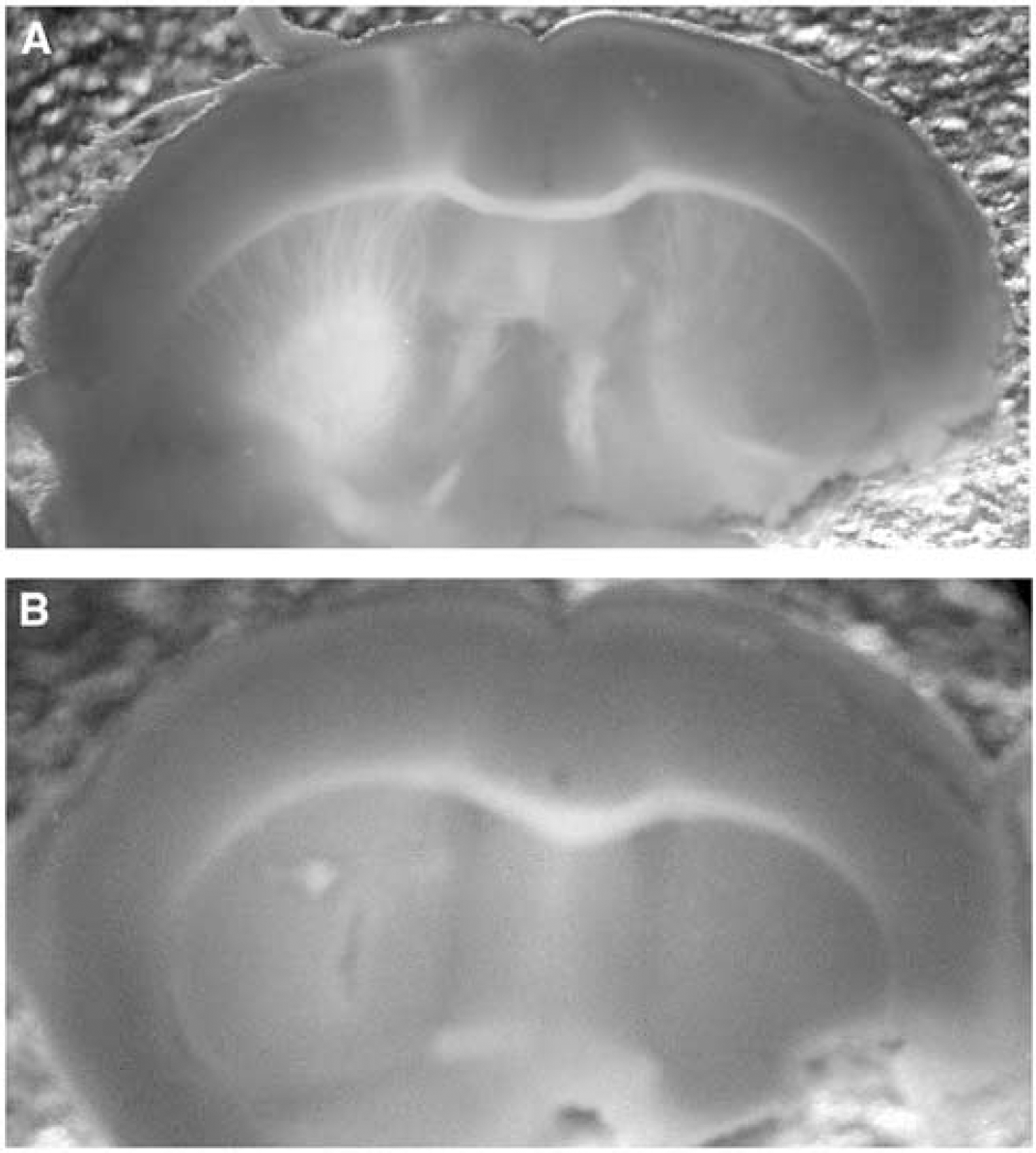
Heme oxygenase (HO)-2 gene deletion attenuates stroma-free hemoglobin toxicity in striatum. Sections of brain from wild-type (
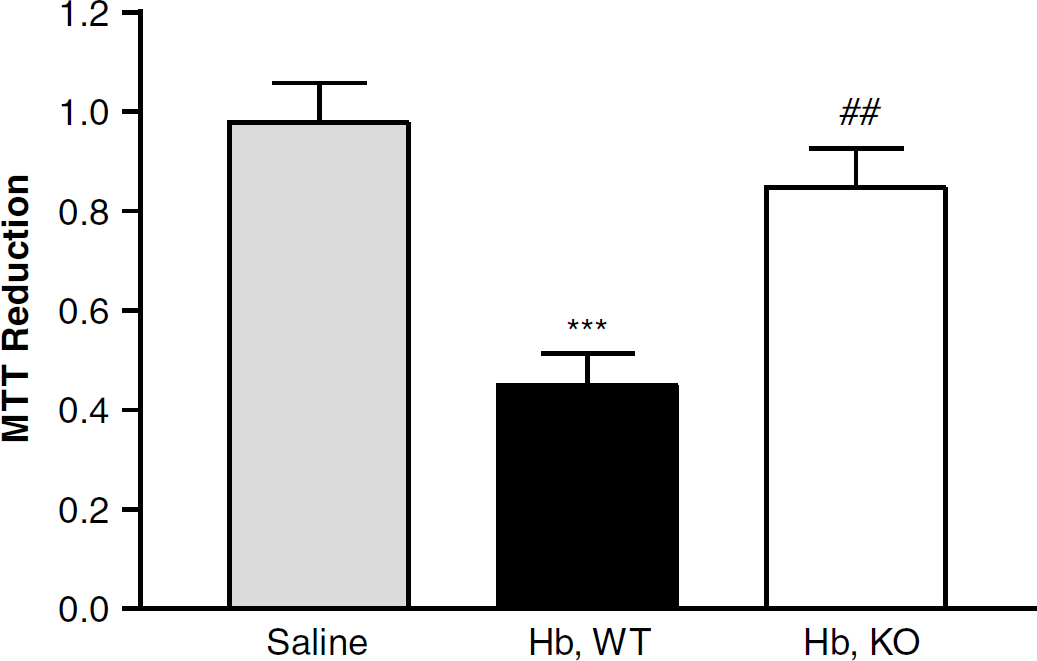
Heme oxygenase (HO)-2 gene deletion reduces the cytotoxic effect of stroma-free hemoglobin in vivo. Bars represent striatal cell viability 72 h after Hb or saline injection, as assessed by MTT reduction to formazan. MTT reduction (mean ± S.E.M., n = 10/condition) is expressed as a fraction of that in the hemisphere contralateral to the injection site. The saline group contained five knockout and five wild-type mice. ***P <0.001 versus ratio in mice injected with saline, ##P <0.01 versus that in wild-type (WT) mice.
Two wild-type and two knockout mice injected with SFHb for this cell viability experiment died before 72 h, and were not included in the above data analysis. If each is assumed to have a ratio of 0 (i.e., 100% striatal cell death in the injected side), then the mean ratios would be reduced to 0.38 ± 0.07 and 0.71 ± 0.11 for the wild-type and knockout groups, respectively (P <0.05, Bonferroni multiple comparisons test).
HO-2 Gene Deletion Reduces Striatal Protein and Lipid Oxidation After SFHb Injection
To determine if the protective effect of HO-2 gene deletion was associated with evidence of reduced oxidative stress, striatal malondialdehyde and protein carbonyl content were determined. In wild-type mice, SFHb injection was associated with a 1.6-fold increase in protein carbonylation at 24 h and 2.5-fold increase at 72 h, compared with the contralateral striatum (Figure 4). In HO-2 knockout mice, no increase was observed 24 h after SFHb injection, and a 1.7-fold increase was observed at 72 h. Consistent with this observation, striatal malondialdehyde was increased 3.5-fold in wild-type mice 72 h after SFHb injection, but was increased only two-fold in knockouts (Figure 5).
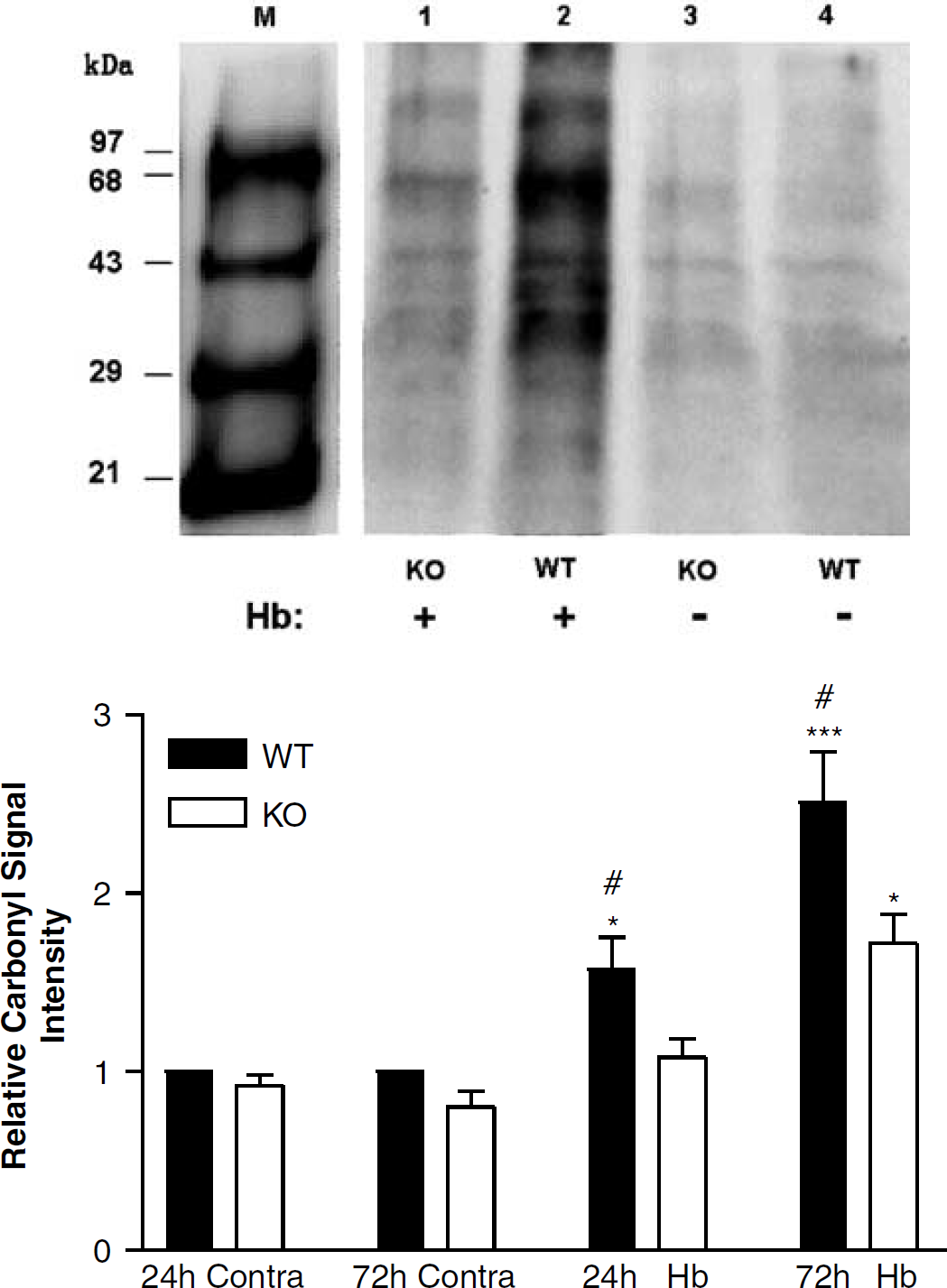
Striatal protein oxidation after stroma-free hemoglobin injection is reduced by HO-2 gene deletion. Top: representative immunoblot of proteins from wild-type (WT) and HO-2 knockout (KO) mice. M: molecular weight standard with attached DNP residues. Lanes 1, 2: striatal proteins from HO-2 knockout and wild-type mice, 24 h after injection of 6 μL stroma-free Hb; lanes 3, 4: respective contralateral striata. Bottom: mean carbonyl signal intensities (±S.E.M., n = 4/condition) in Hb-injected and contralateral (contra) striata, 24 and 72 h after Hb injection. *P <0.05, ***P <0.001 compared with corresponding contralateral striatum, #P <0.05 compared with signal in HO-2 knockout mice at 24 or 72 h.
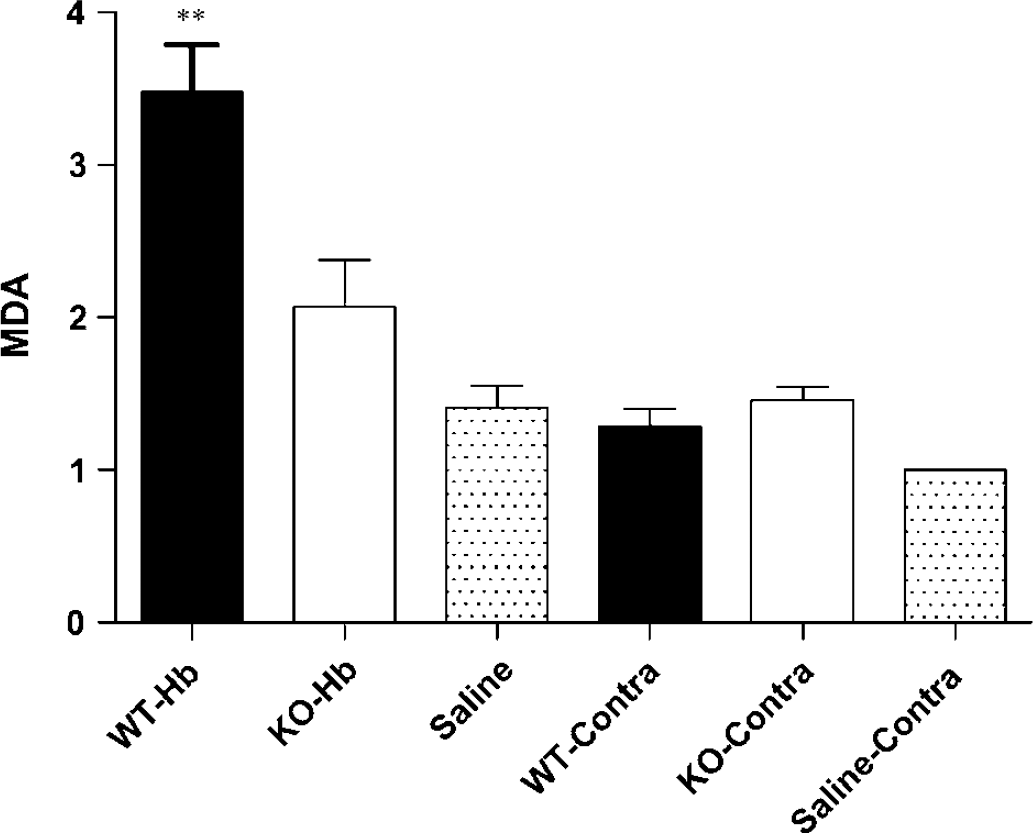
Striatal lipid peroxidation after stroma-free hemoglobin injection is attenuated by HO-2 gene deletion. Mean malondialdehyde content in injected or contralateral (contra) striata (±S.E.M., n = 6/condition), 72 h after injection of 6 μl stroma-free Hb or saline. Malondialdehyde levels were normalized to that in the striatum contralateral to saline injection in control mice (= 1.0).
Effect of HO-2 Gene Deletion on Neuronal Morphology After SFHb Injection
At 3 days after SFHb injection, most neurons that were immediately adjacent to the injection site had an abnormal appearance, with condensed nuclei and absent or eosinophilic cytoplasm (Figure 6). The number of morphologically normal neurons progressively increased with increasing distance from the injection site. In a zone 360 to 600 μm from the injection site, they were quantified by cell counts. The number of morphologically normal neurons per field was significantly greater in HO-2 knockout mice (Figure 7).
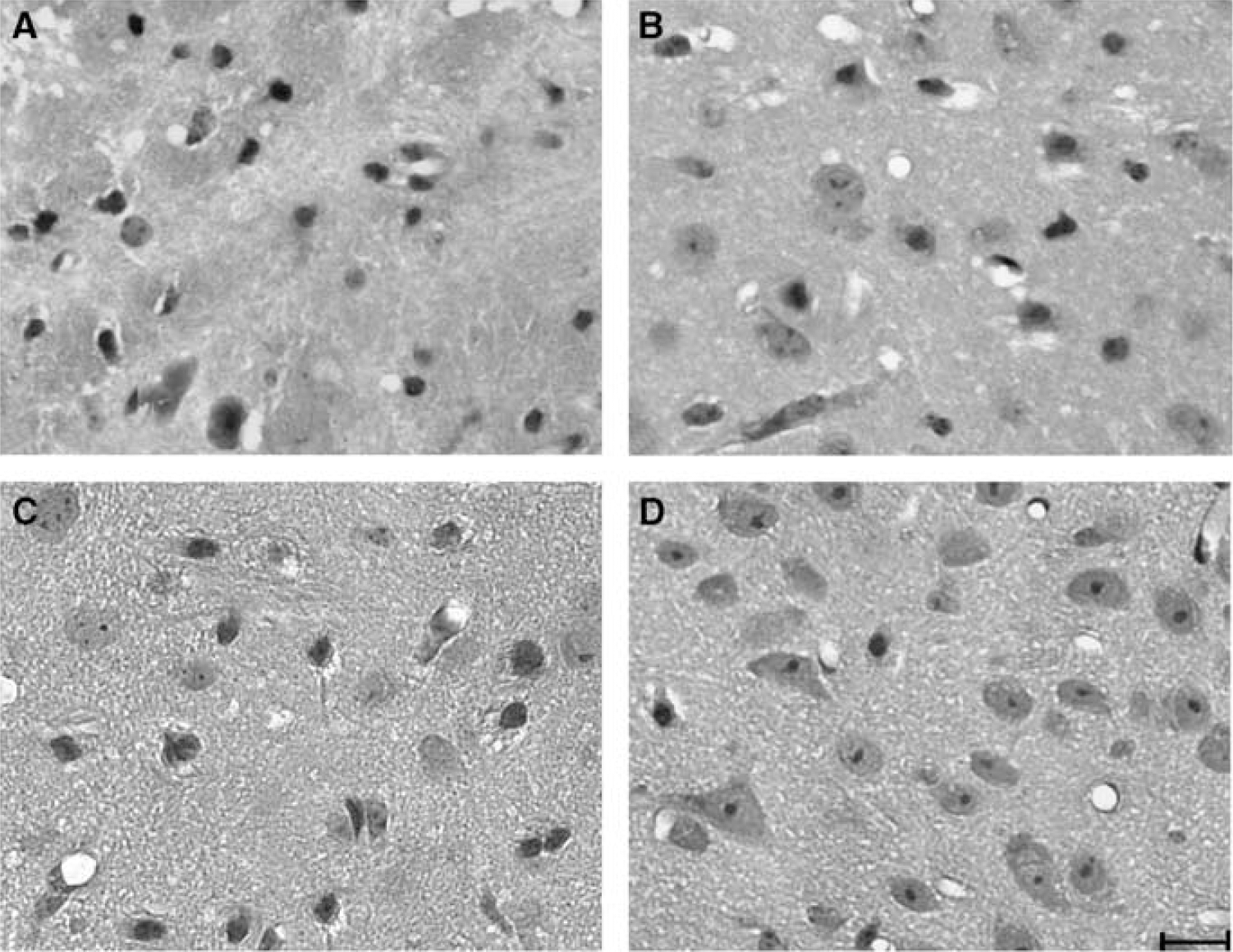
Morphologic appearance of striatal cells, 72 h after stroma-free hemoglobin injection, in wild-type (
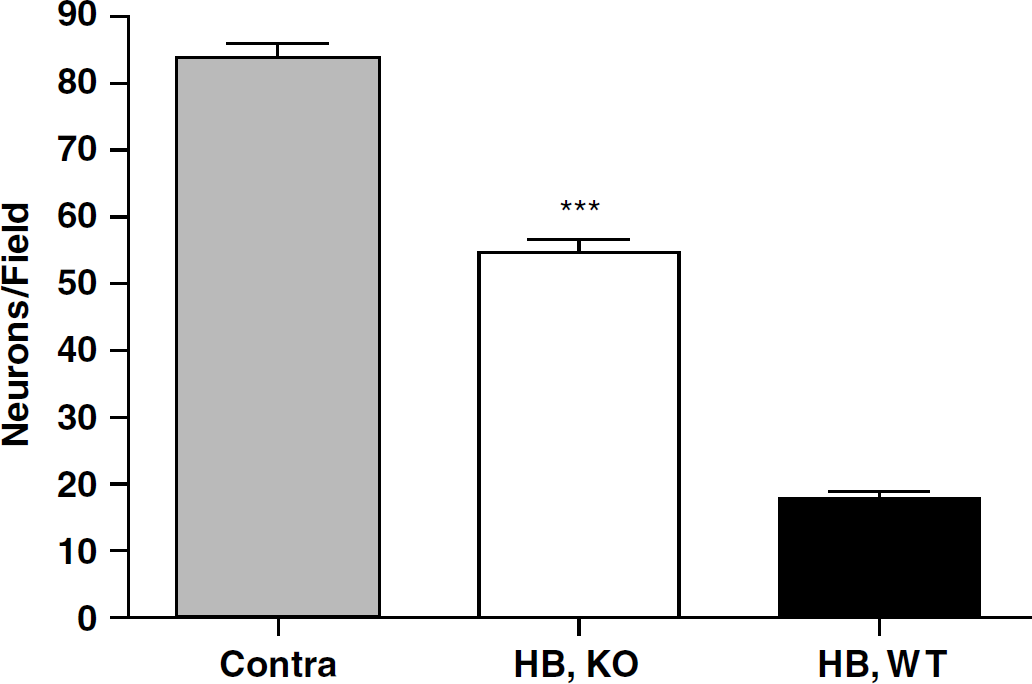
Bars represent mean number of morphologically normal neurons per 320 × 240 μm field, captured at an inner boundary that was 360 μm from the injection site, 72 h after stroma-free hemoglobin injection. ***P <0.001 versus mean number in wild-type mice, n = 3 mice/condition.
Effect of HO-2 Gene Deletion on HO-1 Expression
One limitation of experimental studies involving targeted gene deletion is that cells may compensate by increasing expression of another protein that has the same function as the target gene product. This may be particularly problematic for studies using HO-2 knockouts, because HO-1 is readily induced by a variety of stimuli including oxidative stress. Striatal HO-1 expression was therefore compared in HO-2 knockouts and wild-type controls by Western blot analysis. Consistent with prior observations (Turner et al, 1998), SFHb increased HO-1 expression in the injected striata of both wild-type and HO-2 knockout mice. The HO-1 band density was significantly greater in the former at 6 and 24 h after injection (Figure 8), but had equalized by 72 h (Figure 9). Heme oxygenase-1 expression was not significantly altered by saline injection.
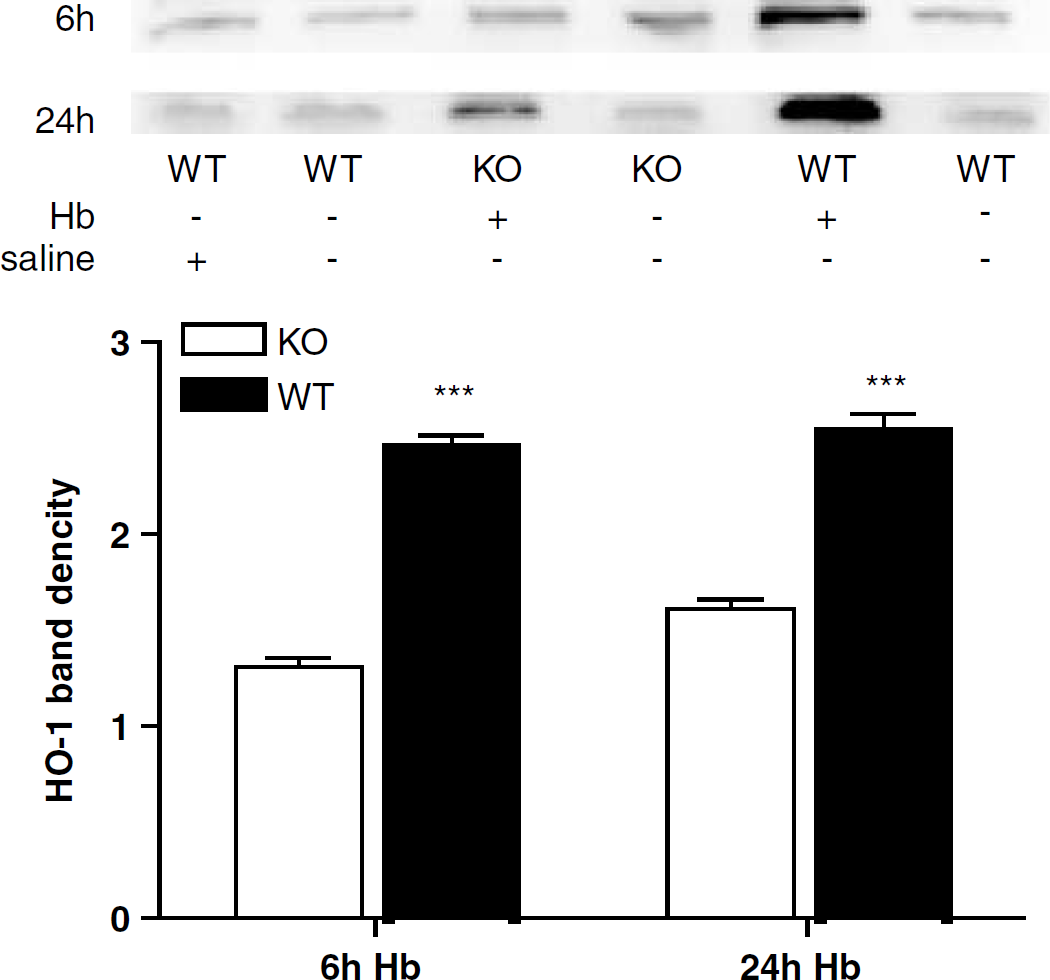
Greater HO-1 induction in wild-type mice than in HO-2 knockout mice at 6 and 24 h after stroma-free hemoglobin injection. Immunoblots of lysates from wild-type and HO-2 knockout striata were stained with polyclonal antibodies to HO-1. The mean band densities (±S.E.M., n = 3/conditon) were normalized to that of contralateral striata of WT mice injected with Hb (= 1.0). ***P <0.001 versus mean value in KO mice at same time point.
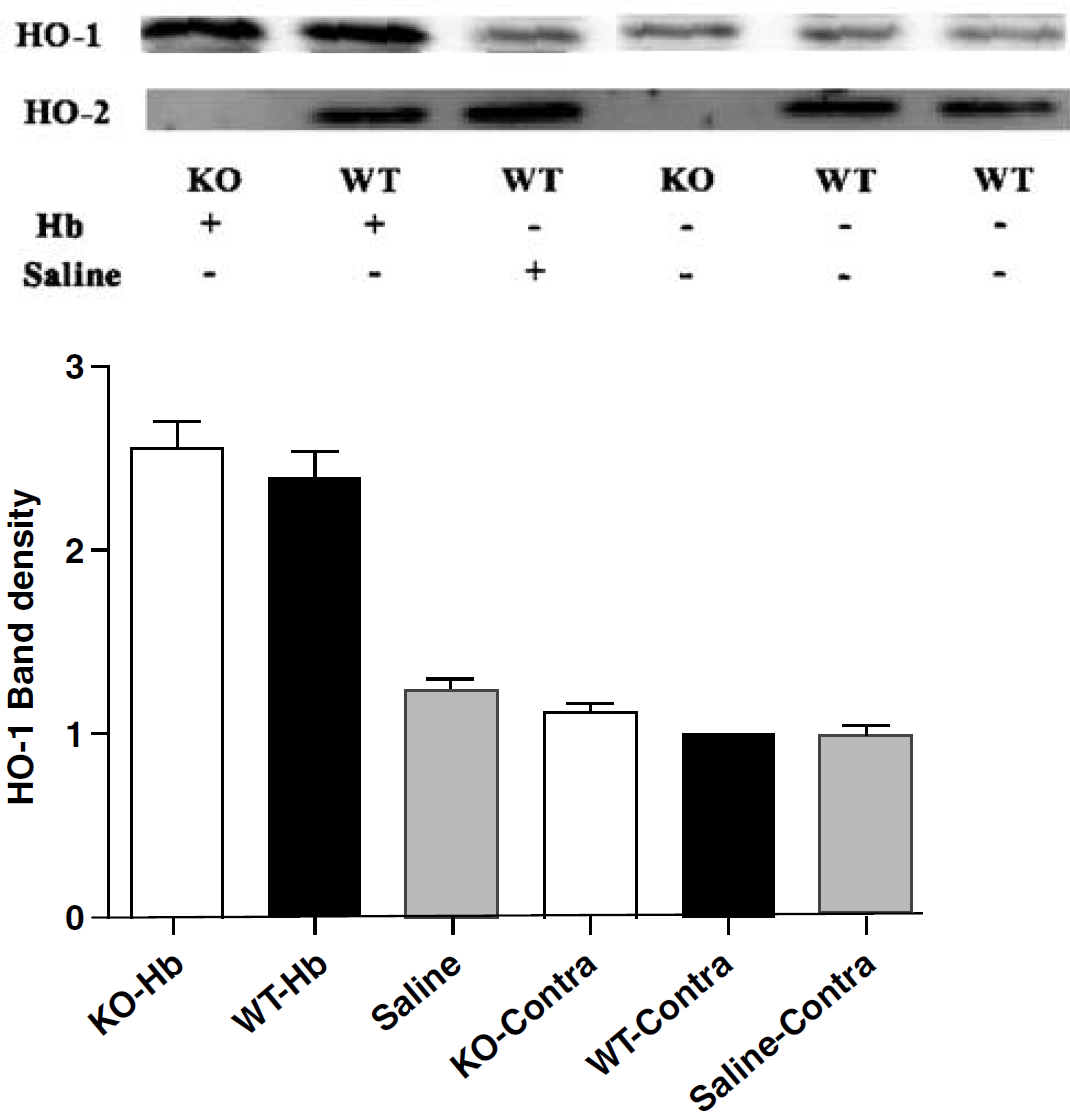
Heme oxygenase (HO)-1 expression is similar in wildtype and HO-2 knockout mice 72 h after stroma-free hemoglobin injection. Immunoblots of lysates from wild-type and HO-2 knockout striata 72 h after Hb or saline injection, stained with polyclonal antibodies to either HO-1 or HO-2. The mean HO-1 protein band densities (±S.E.M., n = 3 each) in the bar chart were normalized to that of contralateral striata of wildtype mice injected with Hb (= 1.0).
Discussion
The present study accomplishes two ends. First, we have shown that injection of a physiologic concentration of SFHb into the mouse striatum produced a 2.5-fold increase in protein oxidation and a 3.5-fold increase in lipid peroxidation within 3 days. Striatal cell viability, as detected by the ability to reduce MTT, was decreased by 55% in this same time interval. These results support the hypothesis that extravascular Hb is an oxidative neurotoxin in the CNS, and are consistent with prior observations in cell culture models and in vivo (Huang et al, 2002; Regan and Panter, 1993; Sadrzadeh et al, 1987; Wang et al, 2002). Second, we have observed that this injury was significantly attenuated in mice lacking the HO-2 gene, in agreement with the protective effect of HO inhibitors in models of Hb or whole blood toxicity (Huang et al, 2002; Koeppen et al, 2004; Wagner et al, 2000).
The effect of the HO enzymes on oxidative injury has been controversial in the CNS and elsewhere, as widely disparate results have been reported. It is becoming increasingly apparent that HO may have both proxidant and antioxidant effects (Ryter and Tyrrell, 2000), and that its net effect depends on both the cell type and the nature of the oxidative insult. Its antioxidant properties are likely mediated by its catalysis of heme breakdown to biliverdin, which is an antioxidant per se and is converted to another antioxidant, bilirubin, by biliverdin reductase (Stocker et al, 1987). This protective effect appears to predominate in models relevant to CNS ischemia and trauma (Chang et al, 2003; Doré et al, 1999a; Doré et al, 1999b; Panahian et al, 1999). However, in the presence of excess substrate, which occurs after hemorrhage, HO activity may enhance oxidation of cellular proteins and lipids (Lamb et al, 1999; Levere et al, 1989). This phenomenon is attenuated by deferoxamine (Lamb et al, 1999; Sadrzadeh et al, 1987), and is therefore likely mediated by release of iron from heme (Koeppen and Dickson, 2002).
Consistent with prior observations (Matz et al, 1997), HO-1 expression was increased by SFHb injection in both wild-type and HO-2 knockout mice; however, its upregulation was more rapid in the former. Heme oxygenase-1 is induced by a variety of oxidative cytotoxins in the striatum, and it appears to be a sensitive marker of early cell injury (Munoz et al, 2005). This phenomenon is likely due to the presence of the antioxidant response element in the 5' untranslated region of the HO-1 gene, which increases transcription in response to oxidative stress by binding the Nrf-2 transcription factor (Elbirt and Bonkovsky, 1999; Ishii et al, 2000). The attenuated HO-1 expression in HO-2 knockout mice at 6 and 24 h may merely reflect a slower progression of injury. In this regard, it is noteworthy that striatal protein oxidation was already detectable in wild-type mice by 24 h, but not in knockouts.
In addition to being a marker of oxidative cell injury, heme breakdown by induced HO-1 may also contribute to cytotoxicity (Lu and Ong, 2001; Schipper et al, 1999). In a prior cell culture study, HO-2 gene deletion attenuated but did not completely prevent the toxic effect of Hb on cortical neurons (Rogers et al, 2003). Heme oxygenase-1 was induced by Hb treatment in both wild-type and knockout cultures. Most of the neuronal death in knockout cultures was prevented by concomitant treatment with the HO inhibitor tin protoporphyrin IX, suggesting that heme breakdown by HO-1 is also deleterious. The delayed expression of HO-1 in HO-2 knockout mice in the present study may therefore account for part of the protection provided by HO-2 gene deletion.
In contrast to its effect on neuronal injury, prior studies suggest that HO decreases the vulnerability of astrocytes to Hb and hemin (Chen and Regan, 2004; Regan et al, 2000; Teng et al, 2004). Both wildtype and HO-2 knockout astrocytes are highly resistant to Hb in culture. This resistance is mediated at least in part by HO activity, because it is markedly diminished by HO inhibitors (Regan et al, 2000). Astrocytes are quite vulnerable to the rapid injury produced by hemin, the oxidized form of heme that accumulates to high micromolar concentrations in hematomas (Letarte et al, 1993). Increasing HO-1 by adenoviral gene transfer before hemin exposure is protective (Teng et al, 2004), while HO-2 gene deletion is deleterious (Chen and Regan, 2004). The disparate effect of HO on heme-mediated injury to neurons and astrocytes may limit the utility of HO inhibitor therapy for CNS hemorrhage. In animal models, its net effect is protective (Huang et al, 2002; Koeppen et al, 2004). It should be noted, however, that the cellular composition of the human and rodent striatum is quite different. Approximately two-thirds of cells in the mouse striatum are neurons (Sturrock, 1980). In contrast, 70% of cells in the striata of middle-aged humans dying of non-neurologic disease are glia (Heinsen et al, 1994). Because HO apparently protects astrocytes from heme-mediated injury but is deleterious to neurons, reducing its activity may have a different net effect on hemorrhagic striatal injury in humans and rodents.
The concentration and injection volume of SFHb were chosen to produce a consistent injury with minimal mortality. Nevertheless, a low mortality rate was observed that was similar in both knockout and wild-type mice. It was accompanied by a large intracranial hemorrhage in all animals. While these hemorrhages may have been produced by mechanical disruption of vessels during needle insertion or injection, examination of striata of control mice injected with saline suggests that this is unlikely. Of 35 mice in this latter group, only one punctate hemorrhage at the needle insertion site was seen when striata were inspected under a dissecting microscope; no deaths occurred. It therefore seems likely that hemorrhage may be a consequence of extravascular SFHb per se. Hemoglobin has been shown to be toxic to endothelial and vascular smooth muscle cells in culture (Comair et al, 1993; Motterlini et al, 1995), and to intact vessels in vivo (Mayberg et al, 1990a). Its toxicity to vascular cells has primarily been associated with arterial spasm and subsequent ischemia in models of subarachnoid hemorrhage (Mayberg et al, 1990b), and is attenuated by HO (Abraham et al, 1995; Ono et al, 2001). Further investigation of the effect of Hb on vessel integrity and rebleeding in intracerebral hemorrhage models seems warranted.
This study was designed to quantify oxidative cell injury in the striatum after SFHb injection, rather than assess metabolic end points. However, animal weight was recorded before injection and 3 days later, and was significantly decreased by Hb in both wild-type and knockout mice. Weight loss has commonly been observed after hemorrhagic and traumatic brain injury, in both animal models and humans (Roe and Rothwell, 1997; Touho et al, 1990). Its etiology has not been precisely defined, but it is likely due to both an increased metabolic rate and hypophagia. The results of this study suggest that this effect is not altered by expression of HO-2.
Intracerebral hemorrhage is a complex injury. Experimental evidence suggests that clot retraction, thrombin proteolysis, transferrin-bound iron toxicity, and a mass effect contribute to cell injury, in addition to the deleterious effect of extracellular Hb (Bullock et al, 1988; Nakamura et al, 2005; Xi et al, 2002; Xi et al, 2000). The model used in this study addresses only the latter mechanism, and therefore does not represent many key elements of clinical CNS hemorrhage. However, by facilitating the investigation of the toxicity of Hb in the intact CNS, without the confounding effects of other injury cascades, it will hopefully complement studies assessing the toxicity of whole blood.
In summary, the present results suggest that HO-2 increases local oxidative stress after injection of SFHb into the neuron-rich mouse striatum. Therapeutic strategies that decrease its expression or activity may protect neurons from heme-mediated injury after CNS hemorrhage. However, the consequence of such approaches on glial and vascular injury should be considered, in light of the protective effect of HO on heme-mediated injury to these cell populations.
Footnotes
Acknowledgements
The authors thank Dr Frank Sharp for providing the HO-2 knockout mice used to establish our colony, and Mr James O'Dowd for his technical assistance in this project.