
Abstract
The intracellular mechanisms that regulate neurogenesis remain unclear. Using neurospheres isolated from the subventricular zone (SVZ) of the adult rat, we investigated the effect of cyclic guanosine monophosphate (cGMP) and its signaling pathway on the induction of neurogenesis. Neurospheres expressed phosphodiesterase 5 (PDE5) and treatment of neurospheres with Sildenafil, a specific inhibitor of PDE5, significantly increased cGMP levels and neurogenesis. In addition, incubation of neurospheres with Sildenafil significantly phosphorylated Akt, which was associated with an increase of phosphorylation of glycogen synthase kinase 3 (GSK-3), a downstream target of Akt. Coincubation of neurospheres with Sildenafil and LY 294002, a pharmacological inhibitor of PI3-K/Akt, abolished Sildenafil-induced phosphorylated Akt and GSK-3. Furthermore, LY 294002 blocked Sildenafil-increased SVZ cell proliferation. These data suggest that Sildenafil-enhanced neurogenesis likely occurs through activation of the PI3-K/Akt/GSK-3 pathway.
Introduction
Cyclic guanosine monophosphate (cGMP) is generated by cytoplasmic-soluble guanylate cyclases and degraded by phosphodiesterase 5 (PDE5) (Ignarro, 1991; Corbin and Francis, 1999). Cyclic GMP effector molecules include cGMP-dependent protein kinases (PKGs), cGMP-regulated phosphodiesterases (PDEs), and cGMP-gated ion channels (Butt et al, 1993). Cyclic GMP is a molecular messenger involved in diverse cellular processes including regulation of cellular proliferation (Firestein and Bredt, 1998). Increases in cGMP levels enhance proliferation of endothelial cells and motor neurons (Zhang et al, 2003; Hood and Granger, 1998). Inhibition of type-I PKG attenuates cGMP-induced sensory neuron precursor proliferation, suggesting that PKGs contribute to the effects of cGMP on cellular function (Firestein and Bredt, 1998).
We have previously reported that treatment of stroke with Sildenafil, a specific inhibitor of PDE5, increased brain cGMP levels and enhanced neurogenesis (Zhang et al, 2002). However, the direct effects of cGMP signaling on neurogenesis in adult subventricular zone (SVZ) cells remain to be determined.
Akt is a downstream target of phosphatidylinositol 3-kinase (PI3-K), and the PI3-K/Akt pathway mediates a cGMP-dependent survival role in cerebellar granule neurons (Ciani et al, 2002) and regulates neurogenesis during Xenopus development (Peng et al, 2004).
Neuronal progenitor and stem cell populations reside within the SVZ and hippocampus of the rodent and human brain (Doetsch et al, 1999; Eriksson et al, 1998; Reynolds et al, 1992; Scheffler et al, 1999). Stroke enhances neurogenesis (Arvidsson et al, 2002; Iwai et al, 2002; Jin et al, 2003; Parent et al, 2002; Zhang et al., 2004, 2001). Precursor cells isolated from the adult SVZ exhibit neural stem cell characteristics in vitro; they can form spheres (self-renew) and differentiate into neurons, astrocytes, and oligodendrocytes (multipotential) (Gritti et al, 1996; Reynolds and Weiss, 1992; Morshead et al, 1994). In the present study, we isolated SVZ cells from the adult rat and tested the hypothesis that inhibition of PDE5 activity enhances neurogenesis. We also investigated the potential mechanisms underlying the effects of PDE5 inhibitor on neurogenesis.
Materials and methods
All experimental procedures were approved by the Institutional Animals Care and Use Committee of Henry Ford Hospital.
Neurosphere Culture
Subventricular zone cells were dissociated from normal (n = 6) male Wistar rats (3 to 4 months), as previously reported (Morshead et al, 1994; Chiasson et al, 1999). The cells were plated at a density of 2 × 104 cells per milliliter in DMEM-F-12 medium (Invitrogen Corporation, Carlsbad, CA, USA) containing 20 ng/mL of epidermal growth factor (EGF, R&D System, Minneapolis, MN, USA) and basic fibroblast growth factor (bFGF, R&D System, Minneapolis, MN, USA). DMEM-F-12 medium contains
Experimental Protocol
(1) To examine the effects of Sildenafil and cGMP on neurosphere proliferation, neurospheres were incubated in the presence of Sildenafil (300 nmol/L, Pfizer Inc., Dublin, Ireland), 8-Br-cGMP (100 μmol/L, Sigma, St Louis, MO, USA), a stable analog of cGMP, with or without LY294002 (a selective phosphatidyl 3-kinase inhibitor, 20 μmol/L, Calbiochem, San Diego, CA, USA) and 1H-[1,2,4]oxadiazolo-[4,3-a]quinoxalin-1-one (ODQ, an inhibitor of guanylate cyclase, 10 μmol/L, Sigma, St Louis, MO, USA). Bromodeoxyuridine (BrdU, 50 μg/mL, Sigma, St Louis, MO, USA) was added 24 h before the assessment of the numbers of neurospheres. Measurements of the numbers and size of neurospheres were performed after 7 days in culture.
(2) To examine the effects of Sildenafil and cGMP on neurosphere differentiation, neurospheres (approximately 80 μm in diameter) were plated directly onto laminin-coated glass cover slips in DMEM-F-12 medium containing 2% FBS, which is referred to as differentiation medium, in the presence of various concentrations of Sildenafil (0, 37.5, 75, 150, and 300 nmol/L), 8-Br-cGMP (0, 1, 10, 50, 100 μmol/L). Every 4 days, one-half of the medium was replaced with fresh medium. Incubation was terminated 14 days after plating, and immunostaining for neuronal and astrocyte markers was performed for evaluation of differentiation.
(3) To examine the effects of Sildenafil on levels of intracellular cGMP, neurospheres were washed twice in Locke's solution, pH 7.4 (composition in mmol/L: NaCl 140, KCl 4.4, CaCl2 2.5, MgSO4 1.2, KH2PO4 1.2, NaHCO3 4, glucose 5.6, EDTA 0.01, glycine 0.003 and HEPES 10) and kept in this medium for 60 min at 37°C. The cells were then preincubated for 30 min at 37°C in Locke's solution containing 0.5 mmol/L 3-isobutyl-1-methyxanthine (IBMX) (Sigma, St Louis, MO, USA) and Sildenafil (300 nmol/L), or Sildenafil (300 nmol/L) and ODQ (10 μmol/L). Levels of cGMP were measured using ELISA.
(4) To examine whether the Akt pathway is involved in Sildenafil-enhanced neurogenesis, neurospheres were left untreated or pretreated with LY294002 (20 μmol/L) for 1 h followed by treatment with or without Sildenafil (300 nmol/L) for 1 h. Levels of Akt and Akt kinase activity were measured using Western blot analysis.
Immunohistochemistry and Quantification
Single or double immunofluorescent staining of cultured cells was performed as previously described (Zhang et al, 2001, 2003). The following primary antibodies were used in the present study: mouse anti-BrdU (1:1,000, Boehringer Mannheim, Indianapolis, IN, USA), mouse anti-β-tubulin III (TuJ-1, 1:1,000, Covance Inc., Princeton, NJ, USA), rabbit anti-glial fibrillary acidic protein (GFAP, 1:500, Dako Cytomation California Inc., Carpinteria, CA, USA), goat anti-GFAP (1:500, Santa Cruz Biotechnology, Inc., Santa cruz, CA, USA); nestin (1:100, BD Biosciences Pharmingen San Diego, CA, USA), rabbit anti-phosphodiesterases (PDE5, 1:500, FabGennix International Inc., Shreveport, LA, USA). Cultured cells were fixed in 4% paraformaldehyde for 15 to 20 mins at room temperature. Nonspecific binding sites were blocked with 5% normal goat serum in MEM/HEPES medium for 30 mins at room temperature. The cells were then incubated with the primary antibodies listed above and with FITC- or Cy3-conjugated secondary antibodies. Nuclei were counterstained with 4', 6'-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA, USA).
The number of BrdU immunoreactive cells, TuJ1-positive neurons, GFAP-positive astrocytes, nestin and PDE5-positive cells, and total DAPI cell number per neurosphere were counted and the percentage of each cell type per sphere was determined. Approximately 30 spheres per group were analyzed.
ELISA for cGMP
Cyclic GMP levels of the cultured cells were determined using a commercial cGMP ELISA kit according to the manufacturer's instruction (R&D System, Minneapolis, MN, USA) (Zhang et al, 2003).
Western Blot Analysis
Cell extracts for Western blot analysis were prepared by washing the cells three times with PBS and lysing them in lysis buffer containing 200 mmol/L Tris–HCL, pH 7.4, 1 mmol/L EDTA, 0.1% sodium dodecyl sulfate (SDS) and 100 mmol/L NaCl, 1% nonidet P-40, 1% deoxycholic acid, 10% glycerol, 1 mmol/L NaVO3, 50 mmol/L NAF, and a protease inhibitor cocktail (Boehringer Mannheim, Indianapolis, IN, USA). The lysates were collected in micro***centrifuge tubes, sonicated for 10 secs and centrifuged at 14,000g for 20 mins. Protein concentration in the supernatants of cell extract was determined using a BCA protein assay kit (Pierce, Rockford, IL, USA). Equal amount of proteins were loaded on 10% SDS-polyacrylamide gel and after eletrophoresis, the proteins were transferred to nitrocellulose membranes. The blots were subsequently probed with the following antibodies: phosphospecific Akt (Ser473, Cell Signaling Technology, Inc., Beverly, MA, USA), phosphospecific Erk (Santa Cruz Biotechnology, Inc., Santa cruz, CA, USA). For detection, horseradish peroxidase (HRP-)- conjugated secondary antibodies were used (1:2,000) followed by enhanced chemiluminescence (ECL) development (Amersham, Buckinghamshire, UK). Normalization of results was ensured by running parallel Western blots with phosphorylation-independent antibodies or α-actin antibody. The optical density was quantified using an image processing and analysis program (Scion image, Ederick, MA, USA).
Assay of Akt Kinase Activity
Akt kinase activity of the cultured cells was determined using a commercial Akt kinase activity kit, according to the manufacturer's instruction (Cell Signaling Technology, Inc., Beverly, MA, USA). Briefly, the cultured cells were lysed and the protein concentration of the supernatants was determined using the Bio-Rad protein assay reagent. The proteins (250 μg) were incubated with gentle rocking at 4°C overnight with immobilized anti-Akt antibody crosslinked to agarose hydrazide beads. After Akt was selectively immunoprecipitated from the cell lysates, the immunoprecipitated products were resuspended in 40 μL of kinase assay buffer containing 200 μmol/L ATP and 1 μg of glycogen synthase kinase 3 (GSK-3) fusion protein (Cell Signaling Technology, Inc., Beverly, MA, USA). The kinase reaction was allowed to proceed at 30°C for 30 mins and stopped by the addition of SDS sample buffer. Reaction products were resolved by 12% SDS-PAGE followed by Western blotting (Hayakama et al, 1999) with an anti-phospho-GSK-3α/β antibody as described previously (Hayakawa et al, 2000).
Real-Time RT-PCR
Quantitative PCR was performed using SYBR Green real-time PCR method. Total RNA was isolated from neurosphere cultures using the Stratagene Absolutely RNA MicroRNA isolation kit (Stratagene, La Jolla, CA, USA). Quantitative RT-PCR was performed on an ABI 7000 PCR instrument (Applied Biosystems, Foster City, CA, USA) using 3-stage program parameters provided by the manufacturer, as follows: 2 mins at 50°C, 10 mins at 95°C, and then 40 cycles of 15 secs at 95°C and 1 min at 60°C. Specificity of the produced amplification product was confirmed by examination of dissociation reaction plots. A distinct single peak indicated that single DNA sequence was amplified during PCR. PCR products were run on 2% agarose gels to confirm that correct molecular sizes were present. Each sample was tested in triplicate using quantitative RT-PCR, and samples obtained from three independent experiments were used for analysis of relative gene expression data using the 2−-ΔΔCT method (Livak and Schmittgen, 2001). The following primers for real-time PCR were designed using Primer Express software (ABI): GAPDH (FWD: AGA GAG AGG CCC TCA GTT GCT, REV: TTG TGA GGG AGA TGC TCA GTG T), PDE5 (FWD: GAT TGC TGA TTG CCG CTT TAA, REV: CGC TCC GCT GTA TGT ATG AGTT), PKGII (FWD: TCA CAG ACG CCC TGA ATA AGA AC, REV: ATG CAT TCC ACC ATG TCT TTG AT).
Statistical Analysis
One-way analysis of variance (ANOVA) followed by Student–Newman–Keuls test was used. The data were presented as mean+s.d. A value of P<0.05 was taken as significant.
Results
PDE5 Inhibitor Increases Levels of cGMP and Enhances Neurogenesis
Previous studies have shown the presence of PDE5 in Purkinje neurons of the adult mouse brain (Shimizu-Albergine et al, 2003). To examine whether SVZ cells of the adult rat express PDE5, we measured mRNA and protein levels of PDE5 in neurospheres derived from the adult rat. Real-time RT-PCR and Western blot analysis showed the presence of PDE5 mRNA and proteins, respectively, in SVZ cells (Figure 1A and 1B). Double immunostaining revealed that TuJ1 (Figure 1D and 1E) or nestin (Figure 1G and 1H) positive cells were PDE5 immunoreactive (Figure 1C and 1F). We therefore, examined the effect of a PDE5 inhibitor on proliferation of SVZ cells. Single cells from the adult rat at a density of 20 cells/μL were incubated in the growth medium containing a specific PDE5 inhibitor, Sildenafil. At 7 days in vitro (DIV), the total numbers of neurospheres were measured. Sildenafil significantly (P<0.05) increased the numbers and size of neurospheres (Figure 2B and 2D) compared with that in the control group (Figure 2A and 2D). To confirm that the Sildenafil-increased number of neurospheres was because of an increase of cell proliferation and not to an increase of cell aggregation, BrdU labeling was performed and Sildenafil significantly (P<0.05) increased the number of BrdU-positive cells (Figure 2F and 2(H). Together, these results show that inhibition of PDE5 activity induces SVZ cell proliferation.
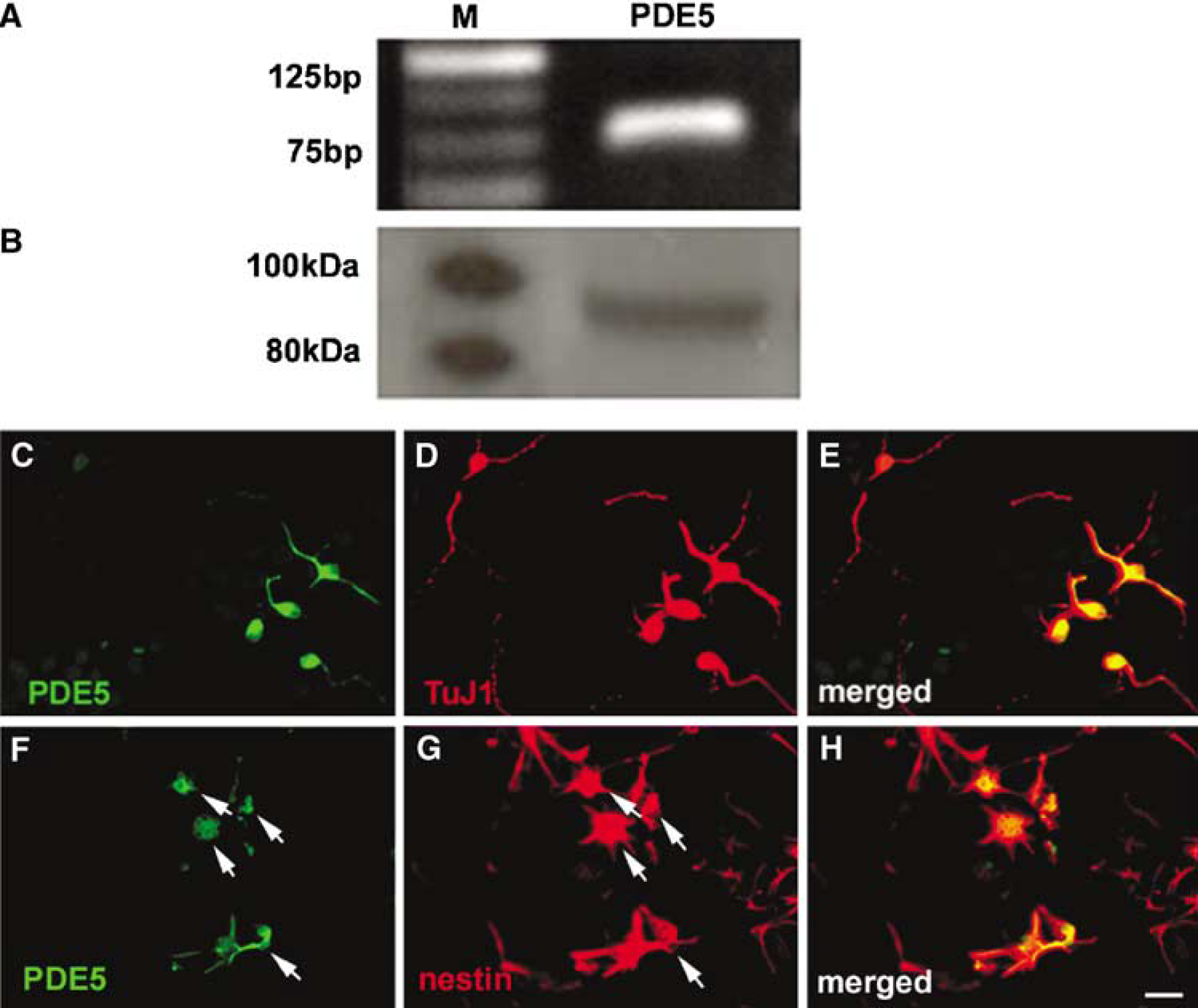
Neurospheres express phosphodiesterase 5 (PDE5). Real-time RT-PCR (
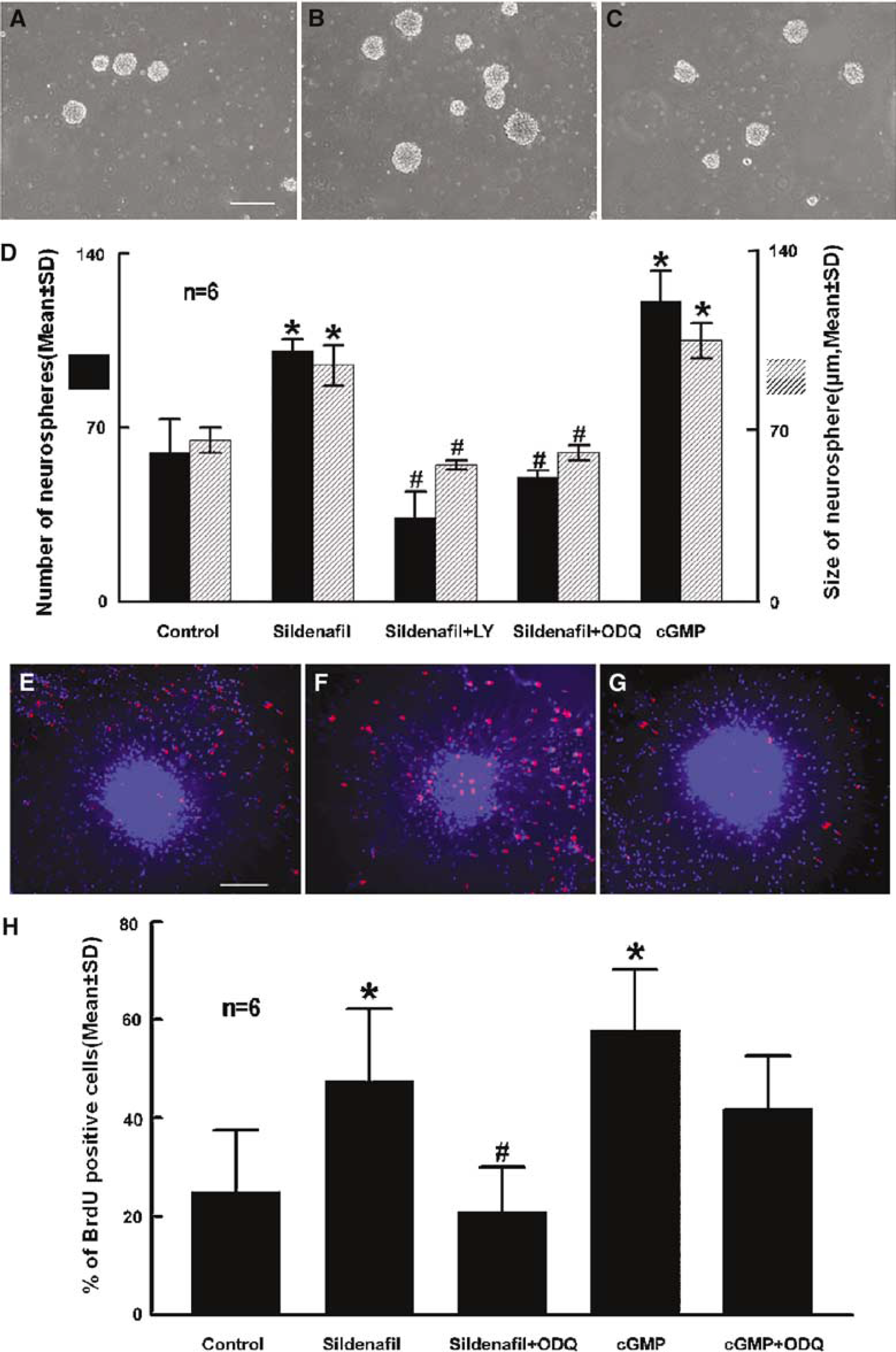
Inhibition of phosphodiesterase 5 (PDE5) activity increases subventricular zone (SVZ) cell proliferation. Panels
To examine whether inhibition of PDE5 increases cGMP levels, ELISA for cGMP levels in neurospheres was performed. ELISA measurements revealed that incubation of neurospheres in the presence of Sildenafil (300 nmol/L) significantly (P<0.05) increased cGMP levels from undetectable to 2.55+1.63 pmol/mg. To confirm the effect of cGMP on SVZ proliferation, neurospheres were incubated with 8-Br-cGMP (100 μmol/L), a stable analog of cGMP. Incubation with 8-Br-cGMP significantly (P<0.05) increased the numbers of neurospheres and BrdU-positive cells, which mimics the effect of Sildenafil on SVZ cell proliferation (Figure 2D and 2(H).
Cyclic GMP is generated by cytoplasmic soluble guanylate cyclases (sGC). To examine whether endogenous levels of sGC are involved in Sildenafil-increased cell proliferation, neurospheres were incubated with Sildenafil (300 nmol/L) in the presence of ODQ (10 μmol/L), a potent inhibitor of sGC (Garthwaite et al, 1995). 1H-[1,2,4]Oxadiazolo-[4,3-a]quinoxalin-1-one significantly (P<0.05) inhibited Sildenafil-induced cell proliferation (Figure 2D, 2G, and 2(H), indicating that proliferation by Sildenafil is dependent on basal activity of sGC in the neurosphere. 1H-[1,2,4]oxadiazolo-[4,3-a]quinoxalin-1-one did not significantly block 8-BrcGMP-induced proliferation (Figure 2H), confirming that this effect is independent of sGC activation.
Neurospheres can differentiate into neurons, astrocytes, and oligodendrocytes in the differentiation medium (Reynolds and Weiss, 1992, 1996). To examine the effect of Sildenafil on the fate of neurospheres, neurospheres were incubated in the differentiation medium with various concentrations of Sildenafil. To measure phenotypes of neurospheres, immunostaining with antibodies against TuJ1 (a marker for neuroblasts), GFAP (a marker for astrocytes) was performed on neurospheres at 14 DIV. Immunostaining revealed that 12%+4.0% and 31+9.1% cells were TuJ-1 and GFAP-positive cells in neurospheres, respectively. Incubation of neurospheres with Sildenafil significantly (P<0.05) increased the numbers of TuJ1 in a dose-dependent manner, with maximal increases at 300 nmol/L (Figure 3A and 3C). However, Sildenafil did not significantly change percentage of GFAP positive cells (31+9.1 for control versus 22+3.9 for 150 nmol/L and 25+8.8 for 300 nmol/L). Incubation of 8-Br-cGMP (100 μmol/L) also significantly increased the numbers of TuJ1-positive cells (Figure 3D).
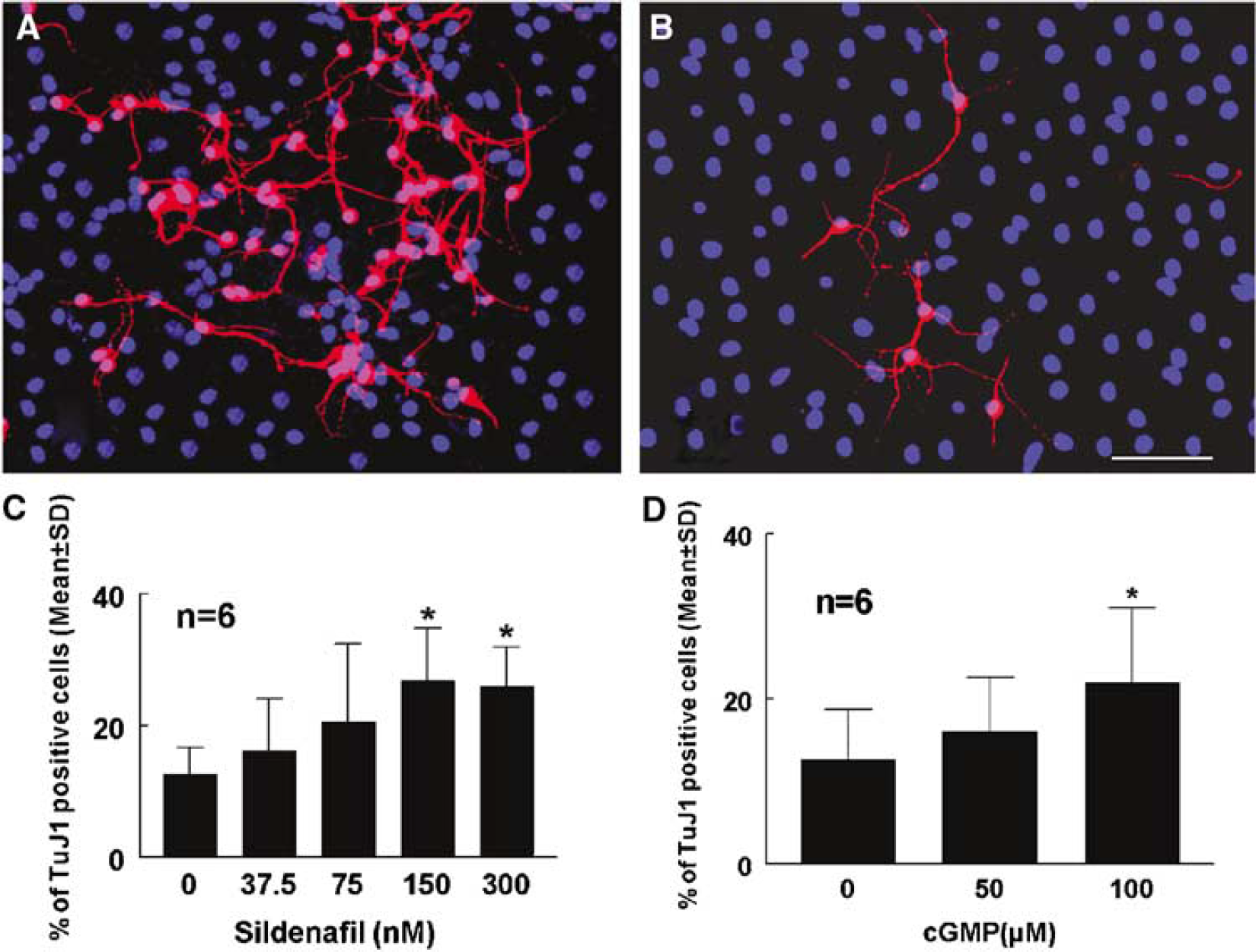
Sildenafil enhances neuronal differentiation. Panels
Blocking PI-3K/Akt Pathway Abolishes Sildenafil-Increased SVZ Cell Proliferation
The Akt pathway mediates a cGMP-dependent survival role in cerebellar granule neurons (Ciani et al, 2002). To explore whether the Akt pathway is involved in Sildenafil-enhanced neurogenesis, we measured phosphorylated Akt and the ability of Akt to phosphorylate its downstream target, GSK-3. Incubation of neurospheres with Sildenafil markedly (P<0.05) increased phosphorylated Akt compared with the control (Figure 4). Interestingly, Sildenafil did not activate phosphorylation of extracellular signal-regulated kinase ERK 1/2 (Figure 4). Phosphorylation of Akt by Sildenafil was associated with increases of phosphorylation of GSK-3 (Figure 4). Incubation of neurospheres with Sildenafil in the presence of LY294002, a selective PI3-K inhibitor, abolished Sildenafil-induced phosphorylated Akt and phosphorylation of GSK-3 (Figure 4). Furthermore, incubation of neurospheres with LY294002 blocked Sildenafil-increased numbers of neurospheres (Figure 2C and 2D). These data suggest that Sildenafil induces neurogenesis through activation of PI3-K/Akt/GSK-3 pathway.
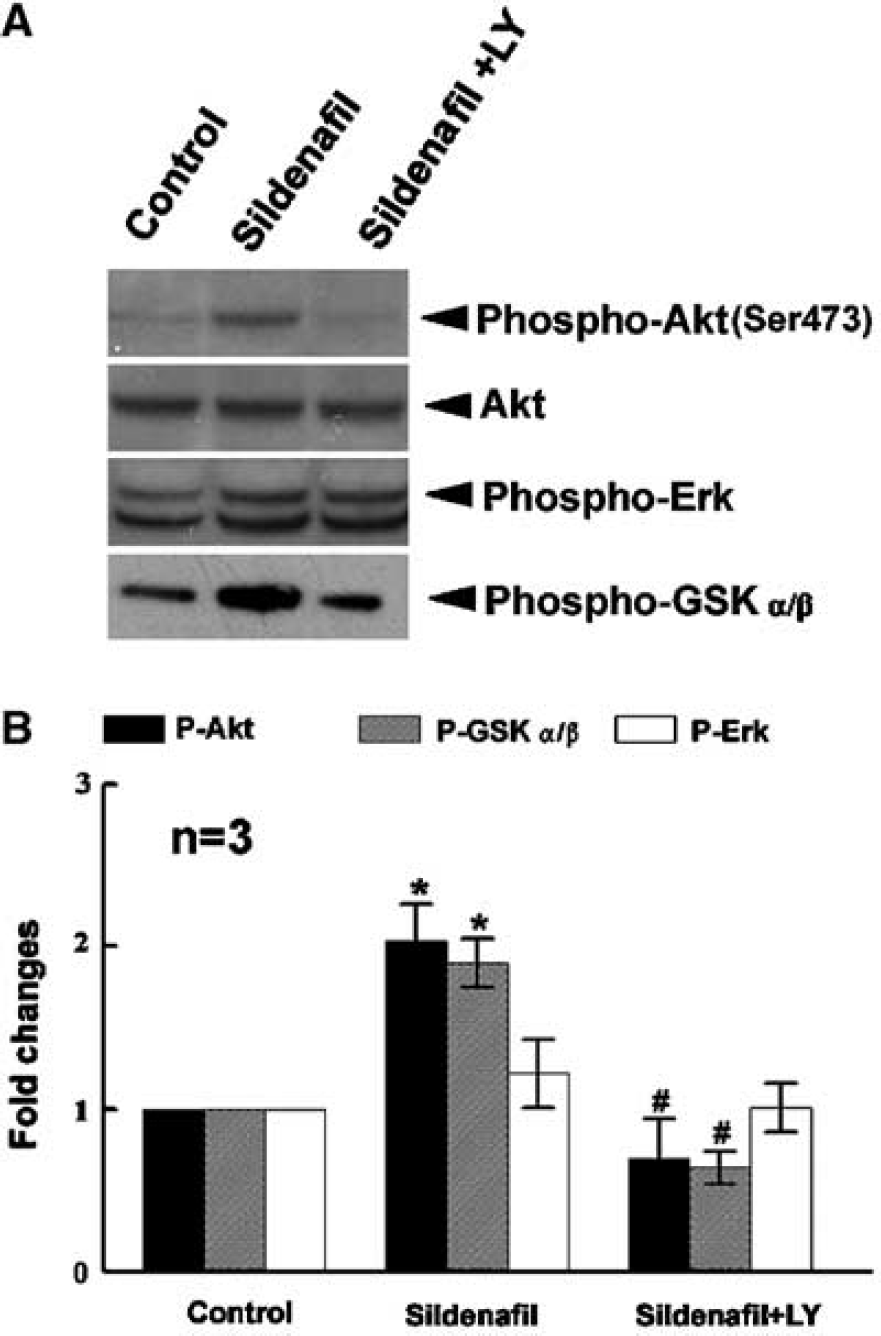
The effects of Sildenafil and LY 294002 on phosphorylation of Akt, GSK-3α/β, and ERK1/2. Western blot analysis (panel
Inhibition of PDE5 Activity Upregulates Expression of Type II cGMP-Dependent Protein Kinase
The intracellular actions of cGMP are primarily mediated by PKG (Lincoln et al, 2001). Sildenafil may regulate neurogenesis by modulating PKG expression. Therefore, using real-time RT-PCR, we quantified expression of PKG genes. Incubation of neurospheres with Sildenafil increased expression of type II PKG by 4- to 6- fold (P<0.05) (Figure 5) but not type I PKG (data not shown), suggesting that type II PKG may be involved in Sildenafil-induced neurogenesis.
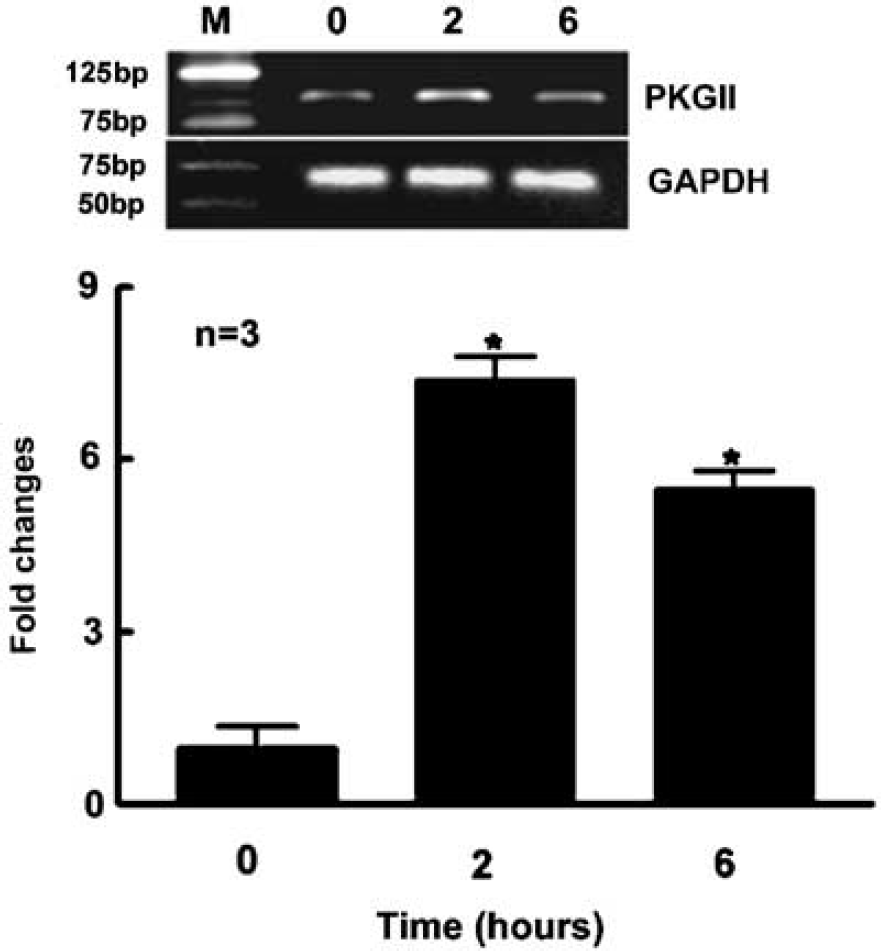
Inhibition of PDE5 activity upregulates expression of type II cyclic guanosine monophosphate (cGMP)-dependent protein kinase. Quantitative measurements of real-time RT-PCR show that incubation of neurospheres with Sildenafil significantly increased type II cGMP-dependent protein kinase (PKG) mRNA 2 and 6 h after incubation. GAPDH was used as an internal control. *P<0.05 versus the control group. The zero time point is the control group.
Discussion
In the present study, we show that incubation of neurospheres with Sildenafil significantly increased cGMP levels and enhanced neurogenesis, which was associated with significant increases of phosphorylated Akt and Akt activation. In addition, inhibition of the PI3-K/Akt pathway abolished Sildenafil-induced neurogenesis. These data indicate that increases in cGMP levels via inhibition of PDE5 activity enhance neurogenesis in adult neural progenitor cells, and the PI3-K/Akt pathway mediates cGMP-enhanced neurogenesis.
Treatment of stroke with Sildenafil-enhances neurogenesis in the adult rat (Zhang et al, 2002). Consistent with these in vivo data, we now show that incubation of neurospheres with Sildenafil significantly increased neurogenesis. In addition, incubation of neurospheres with Sildenafil increased cGMP levels and 8-Br-cGMP mimicked Sildenafil enhanced neurogenesis, whereas inhibition of sGC, which catalyzes guanosine triphosphate into cGMP (Conran, 2004), reduced Sildenafil-enhanced neurogenesis. Although the biological activity of endogenous cGMP increased by Sildenafil may be different from the exogenous cGMP analog, our data suggest that Sildenafil enhances neurogenesis via increases of cGMP levels and endogenous basal levels of cGMP are required for neurogenesis. While cGMP is known to be involved in neuronal survival (Ciani et al, 2002), our results for the first time indicate that cGMP promotes cell proliferation and selective neurogenesis in neurospheres derived from the SVZ of the adult rat. Depending on the cell type, cGMP can have pro- or anti-proliferactive effects (Pilz and Casteel, 2003). Increases in cGMP levels augment endothelial proliferations (Zhang et al, 2003; Hood and Granger, 1998). In contrast, cGMP inhibits mesangial cell and various fibroblast proliferations (Hanada et al, 2001; Takizawa et al, 1997). Here, we show that nestin-positive neural progenitor cells and neurons express PDE5, which may account for the observation that Sildenafil selectively increased numbers of TuJ1-positive cells. Cyclic GMP-dependent protein kinases contribute to the effects of cGMP on cellular and physiologic function (Firestein et al, 1998). Our data show that the PDE5 inhibitor, Sildenafil, significantly increased expression of type II PKG, suggesting that type II PKG may be involved in cGMP-enhanced neurogenesis. To understand fully the role of PKG on neurogenesis, additional experiments are required such as measurements of PKG activity and inhibition of activation of PKG by antagonists of PKG.
The cellular signals mediating neurogenesis are largely unknown. In addition to its many functions, the PI3-K/Akt pathway has been recently shown to mediate neurogenesis during Xenopus development (Vojtek et al, 2003; Peng et al, 2004) Akt is expressed at high levels during neurogenesis (Vojtek et al, 2003; Owada et al, 1997). Glycogen synthase kinase 3 is a downstream target of Akt and activation of Akt inhibits GSK-3 activation by inducing its phosphorylation (Livingstone et al, 1995). In parallel, our data show that Sildenafil increased phosphorylated Akt, which was associated with increases of phosphorylated GSK-3. The effects of Sildenafil on phosphorylated Akt are specific, as Sildenafil did not change phosphorylated ERK1/2. Coincubation of neurospheres with Sildenafil and LY 294002, a pharmacological inhibitor of PI3-K/Akt, abolished Sildenafil-induced phosphorylated Akt and GSK-3, indicating that Sildenafil can functionally activate downstream targets of PI3-K such as Akt and GSK-3. Glycogen synthase kinase 3 is an important component of Wnt signaling, which regulates neurogenesis during development (Castelo-Branco et al, 2003). Furthermore, LY 294002 blocked Sildenafil-increased SVZ cell proliferation. Collectively, our data provide evidence that Sildenafil-enhanced neurogenesis likely occurs through activation of the PI3-K/Akt/GSK-3 pathway.
Footnotes
Acknowledgements
The authors thank Dr. Jieli Chen and Adam M. Robin for technical assistance.