
Abstract
The short- and long-term neuroprotective effects of 2-iminobiotin, a selective inhibitor of neuronal and inducible nitric oxide synthase, were studied in 12-day-old rats following hypoxia-ischemia. Hypoxia-ischemia was induced by occlusion of the right carotid artery followed by 90 minutes of hypoxia (FiO2 0.08). Immediately on reoxygenation, 12 and 24 hours later the rats were treated with vehicle or 2-iminobiotin at a dose of 5.5, 10, 30, or 60 mg/kg per day. Histologic analysis of brain damage was performed at 6 weeks after hypoxia-ischemia. To assess early changes of cerebral tissue, levels of HSP70, nitrotyrosine, and cytochrome c were determined 24 hours after reoxygenation. Significant neuroprotection was obtained using a dose of 30 mg/kg per day of 2-iminobiotin. Levels of HSP70 were increased in the ipsilateral hemisphere in both groups (P<0.05), but the increase was significantly (P<0.05) less in the rats receiving the optimal dose of 2-iminobiotin (30 mg/kg per day). Hypoxia-ischemia did not lead to increased levels of nitrotyrosine, nor did 2-iminobiotin influence levels of nitrotyrosine. In contrast, hypoxia-ischemia induced an increase in cytochrome c level that was prevented by 2-iminobiotin. In conclusion, 2-iminobiotin administered after hypoxia-ischemia provides long-term neuroprotection. This neuroprotection is obtained by mechanisms other than a reduction of nitrotyrosine formation in proteins.
Introduction
Activation of nitric oxide synthase (NOS) contributes to hypoxia-ischemia-induced cerebral injury in the perinatal period (Hamada et al, 1994; Ikeno et al, 2000; Ishida et al, 2001; Tsuji et al, 2000; van den Tweel et al, 2002). Inhibition of both neuronal NOS (nNOS) and inducible NOS (iNOS) using 2-iminobiotin resulted in short-term neuroprotection, whereas simultaneous inhibition of all three isoforms of NOS, that is, nNOS, iNOS, and endothelial NOS (eNOS), which regulates cerebral blood flow, did not (Groenendaal et al, 1999; Marks et al, 1996; Peeters-Scholte et al, 2002). The latter indicates that nNOS and iNOS play a prominent role in the reperfusion/reoxygenation injury of the brain after perinatal hypoxia-ischemia. It has been suggested that neuroprotective effects of nNOS and iNOS inhibition are obtained by a reduced production of peroxynitrite, a highly reactive free radical that can cause nitrotyrosine formation (Koppenol et al, 1992; Numagami et al, 1997). Recent studies indicate that NO may also have direct negative effects on mitochondria independent of nitrotyrosine formation, leading to release of cytochrome c and activation of caspases, which results in apoptosis (Brown and Borutaite, 2001). It is unknown whether reduction of nitrotyrosine formation or direct effects of NO on mitochondria contribute most to the neuroprotective effect of combined nNOS and iNOS inhibition. Furthermore, studies in newborn lambs have demonstrated that neuroprotective effects of NOS inhibition are dose dependent, and become less obvious at higher doses (Dorrepaal et al, 1997).
In the present study, we determined whether 2-iminobiotin, a selective inhibitor of nNOS and iNOS, provides long-term neuroprotection after hypoxia-ischemia in the 12-day-old rat. If so, we wanted to determine the optimal dose of 2-iminobiotin using histology. It has been demonstrated in the same model of hypoxia-ischemia that cerebral levels of HSP70 protein may serve as an early marker of brain injury (Ferriero et al, 1990; van den Tweel et al, 2002). Therefore, HSP70 expression was determined 24 hours after the insult as an indicator of early neuroprotection. To assess the contribution of nitrotyrosine formation to the therapeutic effect of 2-iminobiotin, cerebral nitrotyrosine levels were measured after hypoxia-ischemia.
MATERIALS AND METHODS
Animal Protocol
The animal care committee of University Medical Center Utrecht approved all animal experiments. A total of 93 Wistar rat pups of both sexes were used (Harlan, Zeist, The Netherlands). At postnatal day 12, pups were exposed to a hypoxic-ischemic insult according to an adaptation of the Vannucci-Rice model (van den Tweel et al, 2002). Briefly, the pups were anesthetized with halothane (5.0% induction, 2.0% maintenance) in a mixture of N2O and O2 (1:1). The right common carotid artery was exposed and ligated. The duration of the surgery was 5 to 7 mins, after which the pups were allowed to recover for at least 60 minutes. Subsequently, the pups were placed in an incubator with a constant temperature (37°C) and humidity. Hypoxia-ischemia was induced by lowering FiO2 to 0.08 in N2 for 90 mins. After the hypoxic-ischemic insult, the pups received treatment and were returned to their dams. Eighteen rats underwent anesthesia and neck incision without carotid artery occlusion; these animals served as sham-operated controls. The animals were kept at room temperature with a light:dark cycle of 12:12 hours and had access to food and water ad libitum.
Selective Nitric Oxide Synthase Inhibition
2-Iminobiotin (Sigma-Aldrich, St. Louis, MO, USA) was dissolved in 0.01 mol/L HCl and subsequently the pH was set at 4.2 with 0.1 mol/L NaOH. The animals that were subjected to hypoxia-ischemia were assigned to five groups. Animals were randomly given vehicle (n = 24) or 2-iminobiotin 5.5 (n = 11), 10 (n = 10), 30 (n = 20), and 60 (n = 10) mg/kg per day subcutaneously. The first dose was administered immediately after hypoxia-ischemia, and subsequent doses at 12 and 24 hours.
Histology
Histologic studies were performed 6 weeks after hypoxia-ischemia in 63 rats (sham-operated n = 8, vehicle-treated n = 14, 5.5 mg/kg per day n = 11, 10 mg/kg per day n = 10, 30 mg/kg per day n = 10, 60 mg/kg per day n = 10). After intracardial perfusion with 4% paraformaldehyde, the brains were removed, postfixed, and embedded in paraffin by standard histological procedures. Eight-micrometer-thick sections were cut approximately –3.30 mm from the bregma and mounted on coated slides and stained with hematoxylin and eosin (Klinipath, Duiven, The Netherlands). The histopathology was scored in the regions CA1, CA2, CA3, CA4 of the hippocampus and in the dentate gyrus, and in 6 areas of the parietal cortex on a 4-point scale, ranging from 3: normal; 2: few neurons damaged; 1: moderate number of neurons damaged; 0: almost all neurons damaged and cystic infarction. The maximum cumulative score was 33 points and the minimal score was 0. Subsequently, full section images were captured with a Nikon D1 digital camera (Nikon, Tokyo, Japan) and processed in Adobe Photoshop 5.0 (Adobe Systems Inc., San Jose, CA, USA). The sizes of the ipsilateral and contralateral hemispheres were subsequently measured in arbitrary units and the ratio of ipsilateral and contralateral hemispheres was calculated. The proportion of neuronal damage was expressed by this ratio.
Western Blotting
In a second experiment, P13 sham rats (n = 10) and rats treated with 2-iminobiotin (Sigma-Aldrich, Steinheim, Germany) 30 mg/kg per day (n = 10) or vehicle (n = 10) were killed 24 hours post-hypoxia-ischemia. The brains were rapidly removed and divided into two hemispheres, and the cerebellum was snap-frozen in liquid nitrogen and stored at –80°C. The left and right hemispheres were homogenized in a buffer (50 mmol/L Tris, 5 mmol/L EDTA, and 150 mmol/L NaCl) supplemented with protease inhibitors (leupeptin 10 μg/mL, pepstatin 5 μg/mL, STI 10 μg/mL, orthovanadate 1 mmol/L, and 1 mmol/L PMSF). The homogenates were centrifuged at 13,000 g for 15 mins. Supernatants were collected and stored at –80°C. Protein concentration was determined using Bio-Rad protein assay using bovine serum albumin (BSA) as standard.
Proteins from both hemispheres of the three groups were separated by 10% SDS-PAGE. Proteins were transferred to nitrocellulose membranes (Hybond-C, Amersham, UK) by electroblotting. Membranes were incubated overnight at 4°C with the following antibodies: mouse-anti-HSP70 (Stressgen Biotech, Victoria, Canada), mouse-anti-Nitrotyrosine (Biomol, Plymouth, PA, USA), mouse-anti-cytochrome c (Pharmingen, San Diego, CA, USA). This was followed by incubation for 1 hour with the secondary antibody goat-α-mouse (A3682, Sigma-Aldrich, Steinheim, Germany). To test specificity of the mouse-anti-nitrotyrosine antibody, nitrated BSA and normal BSA were loaded and blotted. Specific bands were visualized by using the enhanced chemiluminescence detection system (ECL, Amersham, UK) and exposure to X-ray film. Autoradiographs were scanned using a GS-700 Imaging Densitometer (Bio-Rad, Hercules, CA, USA).
Statistics
Data are presented as mean ± s.d. SPSS software version 10 (SPSS, Chicago, IL, USA) was used to calculate significance. A Kruskal-Wallis test was used to analyze differences between the treatment groups. Subsequently, Mann-Whitney U-tests were used for further analysis of the difference between vehicle and each dose of 2-iminobiotin. For Western blot analysis, Mann-Whitney U-tests were used to analyze the treatment effect and Wilcoxon tests were used to analyze differences between the contralateral and ipsilateral hemispheres within one group. A P-value <0.05 was considered to be statistically significant.
RESULTS
Histology
The results of histologic examination are presented in Figure 1. The combined neuropathology score of the hippocampus and cortex of the ipsilateral hemisphere of vehicle-treated animals was significantly lower than that of the ipsilateral hemisphere of sham-treated animals (Figure 1A; P<0.05). In addition, the combined neuropathology score of the hippocampus and cortex of the ipsilateral hemisphere of animals that had received 10 or 30 mg/kg per day 2-iminobiotin was significantly higher than that of vehicle-treated animals (Figure 1A; P<0.05). No significant effects of 2-iminobiotin were demonstrated for a dose of 5.5 mg/kg per day or for the highest dose tested of 60 mg/kg per day. Analysis of the neuropathology score of the hippocampus and cortex separately shows that the neuroprotective effect of 10 and 30 mg/kg per day 2-iminobiotin is more pronounced in the cortex than in the hippocampus (Figures 1B and 1C).
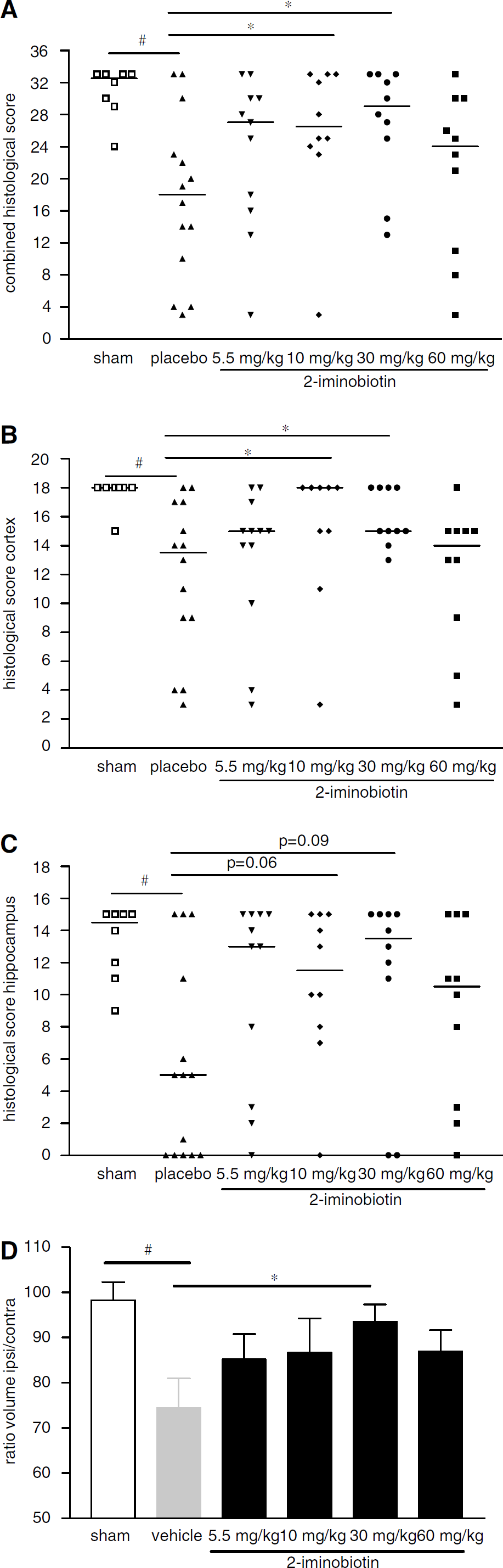
Histopathology (cumulative scores) of rats treated with vehicle (n = 14) or 2-iminobiotin at doses of 5.5 (n = 11), 10 (n = 10), 30 (n = 10), or 60 (n = 10) mg/kg per day and for sham-operated (n = 8) rats 6 weeks after hypoxia-ischemia. Cerebral damage was scored in 6 areas of the cortex and the CA1, CA2, CA3, CA4 of the hippocampus and dentate gyrus on a 4-point scale (range 0 to 3), resulting in a maximal score of 33. In addition, the ipsilateral and contralateral areas were measured (see Materials and methods). (
Analysis of the ratio of the area of the ipsilateral/contralateral hemisphere demonstrated a significant (P<0.01) reduction in the ratio of vehicle-treated rats compared with sham-treated rats (Figure 1D). The ratio of the hemispheres of animals receiving 30 mg/kg per day 2-iminobiotin was significantly (P<0.05) higher than that of vehicle-treated animals. No significant differences were found between the vehicle-treated animals and the animals receiving 2-iminobiotin doses of 5.5, 10 and again those receiving 60 mg/kg per day. A representative example of the histological protection obtained with 30 mg/kg per day 2-iminobiotin is given in Figure 2.
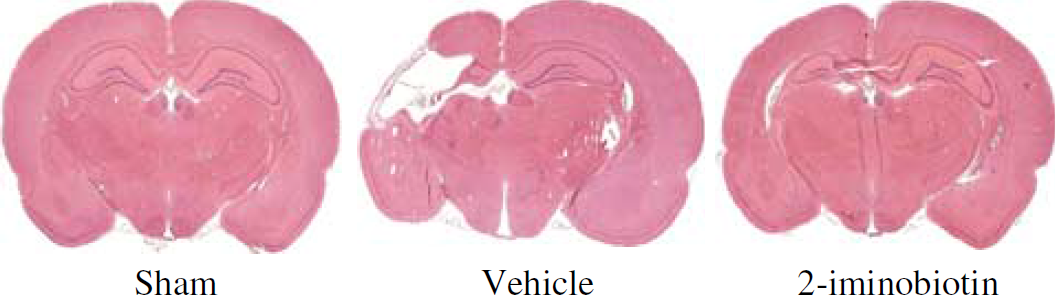
Example of HE-stained specimens of a sham-operated, a vehicle-treated, and an animal treated with the optimal dose of 2-iminobiotin (30 mg/kg per day). The animal treated with 2-iminobiotin shows moderate damage of the hippocampus and cortex.
Nitrotyrosine
Using Western blot analysis, a large number of nitrotyrosine bands were visible in the brain (Figure 3). We analyzed the density of bands of proteins with nitrotyrosine that were clearly visible and migrated at 30, 37, 45, and 70 kDa. No differences in nitrotyrosine staining were found between ipsilateral and contralateral hemispheres in any of the three groups (shown in Figure 3). Moreover, nitrotyrosine staining was not different between sham animals, vehicle-treated animals, or 30 mg/kg per day 2-iminobiotin-treated animals.
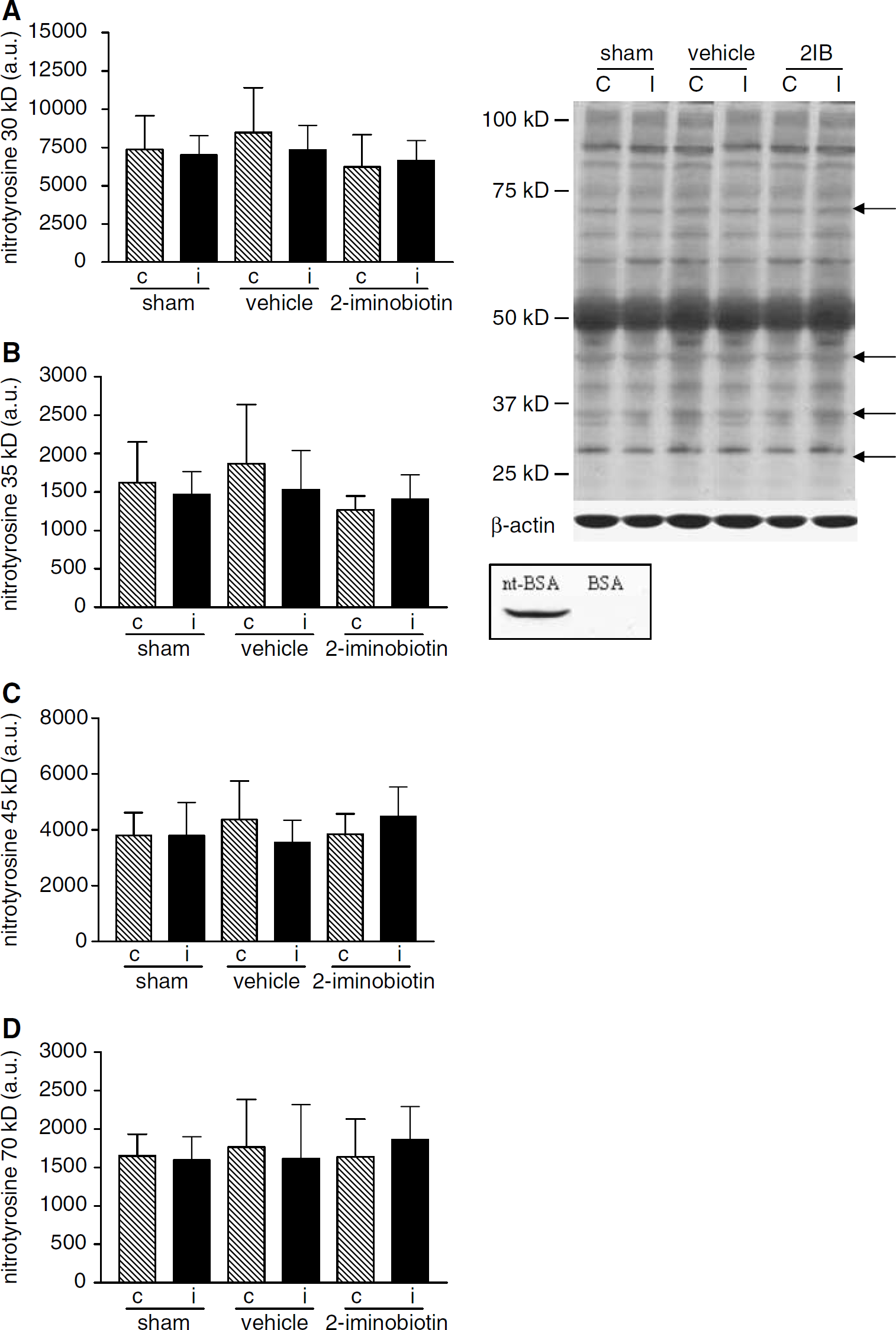
Western blot analysis for nitrotyrosine was performed in homogenates from sham-operated rats and rats treated with vehicle or 2-iminobiotin 30 mg/kg per day (n = 10 per group). We analyzed bands of nitrotyrosine migrating at 30, 37, 45, and 70 kDa. In the top inset a typical nitrotyrosine blot is shown; the arrows point to the analyzed bands. In graphs
HSP70
Using Western blot analysis, only a very faint band for HSP70 could be observed in the ipsilateral and contralateral hemispheres of sham-operated animals. In the ipsilateral hemisphere of both vehicle-treated animals and animals receiving 2-iminobiotin (30 mg/kg per day), a significant (P<0.001) increase in HSP70 protein expression compared with the contralateral hemisphere could be demonstrated (Figure 4). However, the ipsilateral hemispheres of the 2-iminobiotin-treated animals showed a significantly (P<0.05) lower level of HSP70 than the vehicle-treated animals.
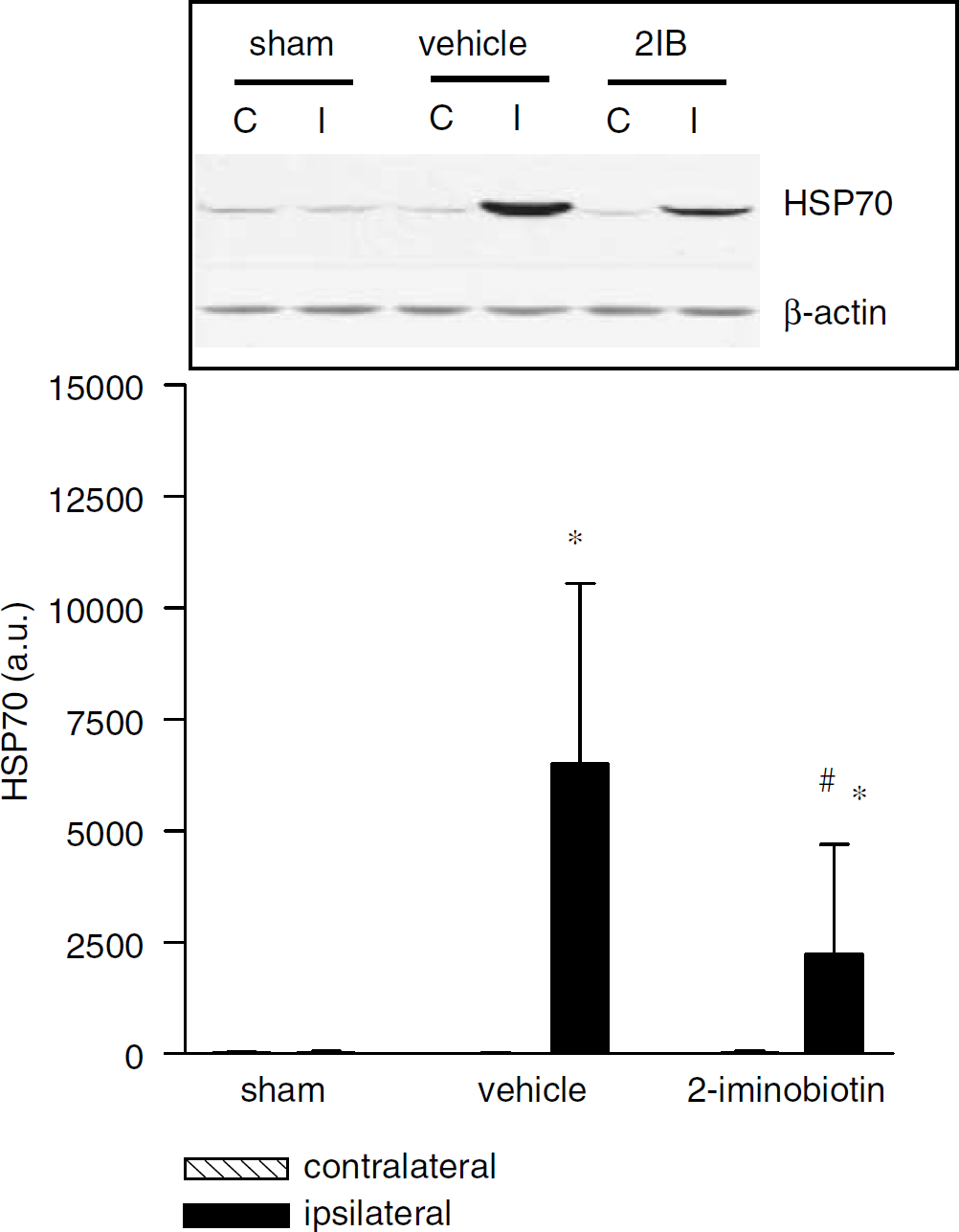
Western blot analysis was performed for HSP70 at 24 hours after hypoxia-ischemia in homogenates from sham-operated rats and rats treated with vehicle or 2-iminobiotin 30 mg/kg per day (n = 10 per group). An increase in HSP70 was observed ipsilateral in both vehicle and rats treated with 2-iminobiotin 30 mg/kg per day. A moderate increase was demonstrated in the 2-iminobiotin-treated rats. The inset depicts a representative Western blot for the different groups. Data are represented as mean ± s.d. *P<0.05 versus contralateral, #P<0.05 versus vehicle.
Cytochrome c
Using Western blot analysis, a faint band for cytochrome c could be observed in sham-operated animals. The level of cytochrome c was increased in the brain of vehicle-treated animals 24 hours after hypoxia-ischemia (P<0.05). Moreover, treatment with 2-iminobiotin (30 mg/kg per day) completely prevented this increase in cytochrome c (Figure 5; vehicle versus 2-iminobiotin: P<0.05).
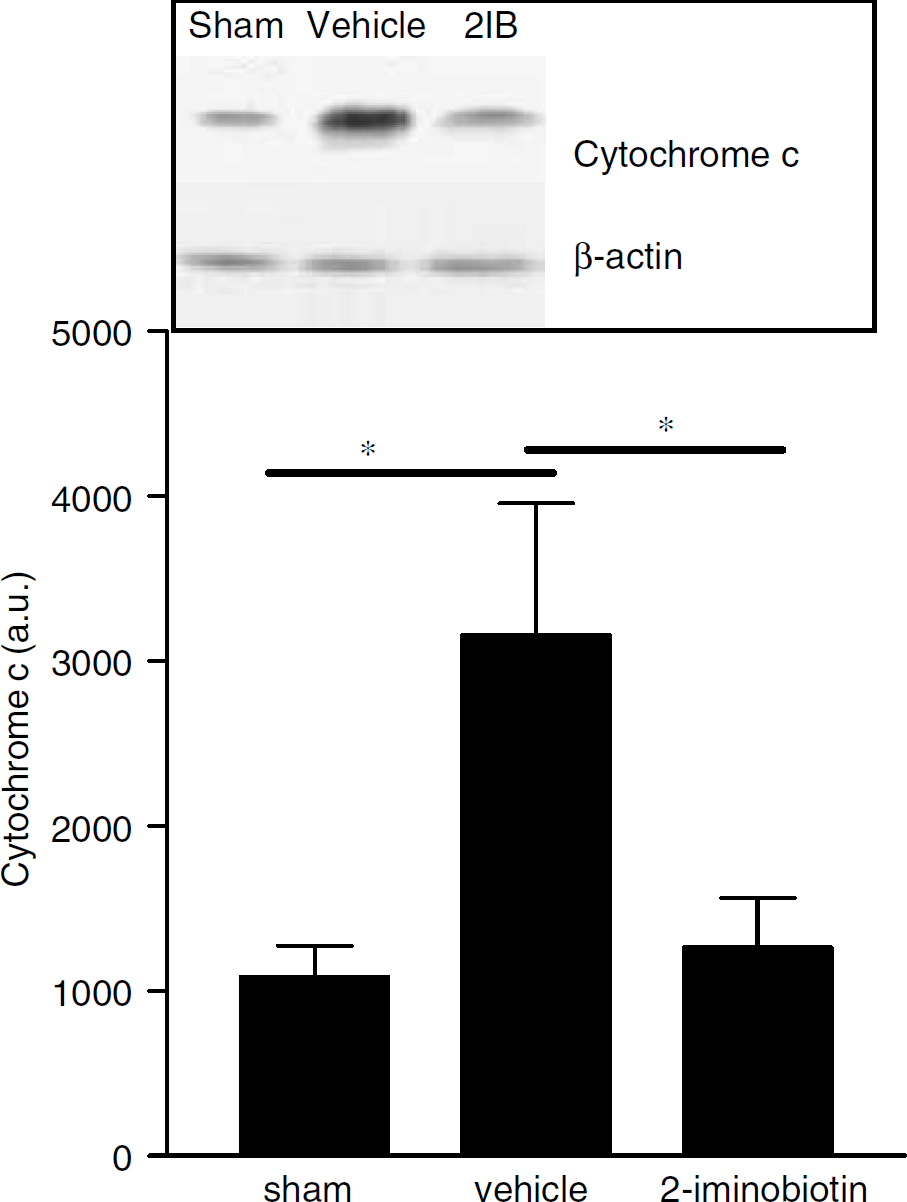
Western blot analysis was performed for cytochrome c at 24 hours after hypoxia-ischemia in ipsilateral hemisphere homogenates from sham-operated rats and rats treated with vehicle or 2-iminobiotin 30 mg/kg per day. The inset depicts a representative Western blot for the different groups. Data are represented as mean ± s.d. *P<0.05.
DISCUSSION
In a previous study, we described short-term neuroprotective effects of 2-iminobiotin in newborn piglets after cerebral hypoxia-ischemia (Peeters-Scholte et al, 2002). The present study shows both short-term and long-term neuroprotective effects of 2-iminobiotin, a selective inhibitor of nNOS and iNOS (Sup et al, 1994), in 12-day-old rats after cerebral hypoxia-ischemia. In a recent study, we have demonstrated a similar long-term neuroprotection using simultaneous inhibition of nNOS with 7-nitroindazole and iNOS with aminoguanidine (van den Tweel et al, 2002). These findings confirm the important role of nNOS and iNOS in neurotoxicity after hypoxia-ischemia, as has been demonstrated in numerous animal models (Dorrepaal et al, 1997; Hamada et al, 1994; Ikeno et al, 2000; Ishida et al, 2001; Suzuki et al, 2002; Takizawa et al, 1999; Tsuji et al, 2000). Reactive oxygen species are excessively formed during, but even more so after, hypoxia-ischemia, which can exceed the antioxidant capacity of the neonatal brain and therefore lead to brain injury (Chan, 2001; Fellman and Raivio, 1997). Nitric oxide and superoxide can react in vivo to form peroxynitrite, a highly reactive compound that is capable of nitrating tyrosine residues of proteins leading to disruption of cell membranes, inhibition of mitochondrial respiration, and DNA damage (Koppenol et al, 1992; Numagami et al, 1997; Reiter et al, 2000).
The cumulative neuropathology score of the hippocampus and cortex indicated that 2-iminobiotin at doses of 10 and 30 mg/kg per day provides long-term neuroprotection. However, measurement of the areas of the left and right hemisphere indicated that 2-iminobiotin is only neuroprotective at a dose of 30 mg/kg per day. In previous studies, neuropathological scores, area measurements, or weight measurements of the ipsilateral and contralateral hemispheres were used to quantify the extent of neuroprotection in the Vannucci-Rice rat model (Bagenholm et al, 1996; Hagberg et al, 1997; Taylor et al, 2002; van den Tweel et al, 2002; Vannucci et al, 1998; Yuan et al, 2001). In the present study, two different methods of quantification of the cerebral injury resulted in slightly different results. Using the neuropathology score, changes in the white matter are not measured. The method that compares the area of brain tissue of ipsilateral and contralateral hemispheres not only quantifies tissue loss of gray matter structures (cortex and hippocampus) but also loss of white matter. With both methods, the dosage of 30 mg/kg per day of 2-iminobiotin showed definite neuroprotection. Therefore, further studies were based on this dose regimen.
Neuroprotective effects of 2-iminobiotin were lost at a dosage of 60 mg/kg per day. It is possible that the selectivity of 2-iminobiotin toward nNOS and iNOS is lost using higher doses, and that eNOS is also inhibited. Endothelial NOS has important vasoactive properties, that is, maintaining cerebral blood flow by causing vasodilatation, which may prove important after cerebral hypoxia-ischemia. Indeed, inhibition of eNOS deteriorates hippocampal dysfunction after transient forebrain ischemia in rats (Jiang et al, 2002). In addition, adult knockout mice lacking eNOS develop larger infarcts after MCA occlusion probably through vasoconstriction (Huang et al, 1996). In contrast, Ferriero et al (1996) suggested that hippocampal damage in nNOS –/– mice was due to increased eNOS activity that occurs selectively in the hippocampus after hypoxia-ischemia. In these animals, the cortex is protected from damage. It may well be possible that basal eNOS activity is required to maintain hippocampal integrity, but that the increase in eNOS activity contributes to damage in the hippocampus. Interestingly, similar to the pattern of damage in the nNOS-/– mice, we show here that the neuroprotective effect of 10 and 30 mg/kg per day of 2-iminobiotin was more pronounced in the cortex than in the hippocampus. The possible role of eNOS in this phenomenon remains to be determined.
Next, we focused on the short-term effects of 2-iminobiotin after hypoxia-ischemia and reperfusion, using the optimal dose of 30 mg/kg per day. The presence of HSP70 in brain tissue has been suggested to be an early marker of injury (Ferriero et al, 1990; van den Tweel et al, 2002). Therefore, to investigate whether reduced HSP70 expression can be used as an early indicator of neuroprotection, we determined the effects of 2-iminobiotin administration on HSP70 expression at 24 hours after hypoxia-ischemia. According to a pilot study in our institute, this is the time point of maximal HSP70 expression (data not shown). The results show that HSP70 was elevated significantly in the ipsilateral side of the brain, but not in the contralateral, uninjured side of the brain 24 hours after hypoxia-ischemia. After treatment with 2-iminobiotin, the levels of HSP70 were significantly lower than in the vehicle-treated animals. These data suggest that inhibition of nNOS and iNOS is an important pathway involved in brain injury after hypoxia-ischemia. However, the incomplete neuroprotection indicates that alternate pathways may also be important and need to be addressed for complete neuroprotection.
It has been demonstrated that nitrotyrosine formation in proteins and the change in function of these proteins is an important pathway of neurodestruction by nitric oxide production, and ensuing peroxynitrite production (Koppenol et al, 1992; Rodrigo et al, 2001). To examine potential changes in nitrotyrosine levels after NOS inhibition with 2-iminobiotin, nitrotyrosine levels were measured 24 hours after hypoxia-ischemia. This time point was chosen as others have demonstrated a significant increase in nitrotyrosine in brain tissue 24 hours after hypoxia-ischemia (Ikeno et al, 2000). Western blot analysis of nitrotyrosine at 24 hours did not show differences between ipsilateral and contralateral hemispheres, nor were nitrotyrosine levels different from sham-operated animals. In addition, no changes of nitrotyrosine bands were found at the time points 3, 6, and 12 hours after hypoxia-ischemia in vehicle-treated animals (data not shown). Conflicting reports of nitrotyrosine levels in brain tissue have been published. Ikeno et al (2000) have demonstrated increased levels of nitrotyrosine in the ipsilateral hemisphere compared with the contralateral hemisphere and compared with baseline in 7-day-old rats peaking at 24 to 48 hours. In contrast, Fernandez et al (2003), using P0 rats, and Alonso et al (2002), using adult rats, showed no overall change in nitrotyrosine levels with Western blot analysis, but a change in the distribution of nitrotyrosine from the nucleus to the cytoplasm with immunohistochemistry. In a pilot study we could not reproduce these findings (data not shown). The differences between the study of Ikeno et al and our findings may find their origin in the postnatal age of the animals studied, since expression of iNOS is dependent on gestational age, being highest between postnatal days 0 and 5 (Alonso et al, 2002; Fernandez et al, 2003).
Since the neuroprotective effects of 2-iminobiotin in the present study were not accompanied by a decrease in nitrotyrosine, we suggest that the beneficial effects of NOS inhibition were achieved by a reduction in NO acting on the mitochondria. Recently, it has been published that NO and peroxynitrate could have direct effects on mitochondria (i.e., without production of nitrotyrosine), resulting in release of cytochrome c followed by activation of caspases and apoptosis, and that this pathway can be operative in neurons (Brown and Bal-Price, 2003; Cooper, 2002; Raoul et al, 2002). Moreover, it has been shown before that neonatal hypoxic-ischemic brain damage is associated with increased release of cytochrome c (Zhu et al, 2003). Our results confirm this observation by showing that cytochrome c levels are increased 24 hours after hypoxia-ischemia. More importantly, we show that after treatment with 2-iminobiotin the levels of cytochrome c were significantly lower than in the vehicle-treated animals. These data support the hypothesis that 2-iminobiotin protects against hypoxic-ischemic brain damage by preventing ***NO-or peroxynitrate-induced mitochondrial damage.
In conclusion, 2-iminobiotin at a dose of 30 mg/kg per day reduced both short-term neuronal injury as demonstrated by HSP70, and long-term neuronal damage as demonstrated with histology. The absence of changes in nitrotyrosine levels in brain tissue after hypoxia-ischemia or 2-iminobiotin treatment indicates that direct effects of NO on mitochondria may be more important for neuronal injury than the formation of nitrotyrosine in proteins. This hypothesis is supported by our observation that cytochrome c levels are decreased after 2-iminobiotin treatment.
Footnotes
Acknowledgements
The authors thank Maaike Yedema, Edina Tjepkema, and Maria de Boer for their excellent technical assistance.