
Abstract
We have shown earlier that 2-iminobiotin (2-IB) reduces hypoxia-ischemia (HI)-induced brain damage in neonatal rats, and presumed that inhibition of nitric oxide synthases (NOS) was the underlying mechanism. We now investigated the effect of 2-IB treatment in P7 rat pups to determine the role of gender and the neuroprotective mechanism. Pups were subjected to HI (occlusion of right carotid artery and 120 mins FiO2 0.08) and received subcutaneous (s.c.) 10 mg/kg 2-IB at 0, 12 and 24 h after hypoxia. After 6 weeks, neuronal damage was assessed histologically. We determined cerebral nitrite and nitrate (NOx) and nitrotyrosine, heat-shock protein 70, cytosolic cytochrome c, cleaved caspase 3, nuclear translocation of apoptosis-inducing factor (AIF) and the effect of 2-IB on NOS activity in cultured cells. 2-Iminobiotin treatment reduced long-term brain damage in female but not male rats. Unexpectedly, 2-IB treatment did not reduce cerebral NOx or nitrotyrosine levels, and did not inhibit NOS activity in vitro. The gender-dependent neuroprotective effect of 2-IB was reflected in inhibition of the HI-induced increase in cytosolic cytochrome c and cleaved caspase 3 in females only. Hypoxia–ischemia-induced activation of AIF was observed in males only and was not affected by 2-IB. Post-HI treatment with 2-IB provides gender-specific long- and short-term neuroprotection in female P7 rats via inhibition of the cytochrome c-caspase 3 neuronal death pathway. 2-Iminobiotin did not alter cerebral NOx nor inhibited NOS in intact cells. Therefore, we conclude that it is highly unlikely that the neuroprotective effect of 2-IB involves NOS inhibition.
Introduction
Hypoxia-ischemia (HI) during the perinatal period is a common cause of mortality and morbidity in the human neonate (Ferriero, 2004). We have described previously that post-HI treatment with the compound 2-iminobiotin (2-IB) is neuroprotective; treatment with 2-IB resulted in improved outcome at short term in a piglet model of cerebral HI (Peeters-Scholte et al, 2002). More recently, we showed both short- and long-term neuroprotection by post-HI treatment with 2-IB in 12-day-old (P12) rat pups, which were used as a model for the full-term human neonate (van den Tweel et al, 2005a).
We and others (Peeters-Scholte et al, 2002; van den Tweel et al, 2005a; Zhu et al, 2004) have suggested that the neuroprotective effect of 2-IB is mediated via inhibition of both the neuronal (n) and inducible (i) isoforms of nitric oxide synthase (NOS), since in vitro studies using purified iNOS and nNOS have shown that 2-IB can inhibit the activity of these enzymes (Sup et al, 1994). However, until now, it has not been proven directly that 2-IB treatment indeed leads to decreased NOS activity in vivo.
In neonatal animals, the extent of brain injury after HI depends on the duration and severity of the insult and also on the maturation of the brain (Grafe, 1994; Yager and Thornhill, 1997; Hagberg et al, 2000). It is known that the contribution of various pathways involved in HI brain damage varies with developmental stage of the brain (du Plessis and Volpe, 2002; Zhu et al, 2005). Moreover, it is becoming increasingly clear from recent human and animal adult studies that gender can be a modulator of cerebral ischemic cell death: there is evidence that both the vulnerability for HI brain damage and the cell death mechanisms after HI differ between males and females (Alkayed et al, 1998; Hurn et al, 2005). For example, it has been shown in in vitro systems that proclivity to either caspase-dependent or caspase-independent apoptotic pathways differs between neurons of female and male origin (Du et al, 2004). Most recently, it was shown in a neonatal mouse model of HI that a more pronounced translocation of apoptosis-inducing factor (AIF) occurred in males whereas caspase 3 was slightly more activated in females (Zhu et al, 2006). As a consequence, gender might influence the outcome of neuroprotective intervention strategies.
The aim of the present study was to investigate short- and long-term effects of treatment with 2-IB after HI in P7 pups, a widely used model for the moderately preterm human neonate (Romijn et al, 1991; Northington et al, 2005). We examined whether gender interacts with the neuroprotective effects of 2-IB treatment after HI. Gender-specific dimorphism in the use of caspase-dependent or caspase-independent apoptotic mechanisms in neonatal cerebral damage was determined by analyzing the activation of several key players: cytochrome c release from the mitochondria and subsequent caspase 3 activation for caspase-dependent apoptosis and nuclear translocation of AIF for the caspase-independent apoptotic pathway. The effect of treatment with 2-IB on these apoptotic pathways in vivo was assessed. Finally, we investigated the mechanism of action of 2-IB in more detail. To get more insight in the contribution of inhibition of iNOS and nNOS to the neuroprotective effect of 2-IB, we determined the effect of 2-IB on the in vivo HI-induced NOx production in the brain. In addition, we examined the effect of 2-IB on the activity of iNOS and nNOS in intact cells in vitro.
Materials and Methods
Animals
Experiments were performed in accordance with international guidelines and approved by the University Medical Center Utrecht experimental animal committee. Timed-pregnant Wistar rats (Charles River, Sulzfeld, Germany) delivered at the Central Laboratory Animal Institute (Utrecht, the Netherlands); the day of birth was considered day 0. Animals were kept on a 12:12 h light: dark cycle and were weaned at 3 weeks of age.
We used a modification of the original Vannucci and Rice model (Rice et al, 1981). Pups were anaesthetized (5 to 10 mins) with isoflurane (5.0% induction, 1.5% maintenance) in O2:N2O (1:1). Blood flow in the right common carotid artery was permanently interrupted by thermocauterization, xylocaine spray (100 mg/mL) (AstraZeneca, Zoetermeer, The Netherlands) was applied and wounds were closed. Animals were kept warm on a heated water mattress (37°C) at any time during surgery. After at least 1 and maximal 3 h of recovery, pups were exposed to 8% O2 in N2 for 120 mins. The gas mixture was humidified and preheated. During hypoxia, the animals were kept on a heated water mattress in a child incubator with a temperature of 37°C. Air temperature in the incubator was monitored and was approximately 37°C throughout the experiment. After hypoxia, the animals returned to their dams and were kept at room termperature. Shamtreated controls underwent anesthesia and incision without further surgical procedures and without hypoxia. Brains from completely untreated pups did not differ from sham-operated pups in any of the parameters tested (data not shown).
2-Iminobiotin (Sigma-Aldrich, St Louis, MO, USA) was dissolved at 1 mg/mL in 0.01 mol/L HCl and pH was adjusted to 4.2 with 0.1 mol/L NaOH. Pups received 2-IB (10 mg/kg) or vehicle (10 ml/kg) subcutaneous (s.c.) at 0, 12 and 24 h after hypoxia. The dose and treatment schedule are identical to the optimal dose used in our earlier study using P12 rats (van den Tweel et al, 2005a) that was designated 30 mg/kg/day. In the present study, we choose to describe the dose as three gifts of 10 mg/kg.
Histology
At 6 weeks after HI, rats received an overdose pentobarbital (300 mg/kg) and were perfused with 4% paraformaldehyde in phosphate-buffered saline. Brains were embedded in paraffin. Coronal sections (6 μm) were cut at approximately −3.30 mm from the bregma and stained with hematoxylin–eosin (Klinipath, Duiven, The Netherlands). Sections were scored in a blinded way in six regions of the parietal cortex and in five regions of the hippocampus (van den Tweel et al, 2005a). Scores ranged from 0 to 3: 3 = normal, < 10% of neurons damaged; 2 = 10% to 50% of neurons damaged; 1=50% to 90% of neurons damaged; 0 = almost all neurons damaged (90% to 100%), gliosis and/or cystic infarction; the maximal total score being 33.
Full section images were captured with a Nikon D1 digital camera (Nikon, Tokyo, Japan). Measurements were performed by using image processing tools in Adobe Photoshop 6.0 (Adobe Systems Inc., San Jose, CA, USA). Contra- and ipsilateral hemisphere or hippocampus was outlined manually, and the ratio of ipsi- to contralateral areas was calculated. All measurements were performed by one investigator (CN) and the intra-observer variance was less than 5%.
Preparation of Samples for Western Blot Analysis
Pups were decapitated at 24 or 48 h post-HI, cerebellum was removed and left and right hemispheres were frozen in liquid nitrogen. Hemispheres were pulverized using a liquid nitrogen-cooled mortar and pestle, divided in three fractions and stored at −80°C, allowing us to use the brain of one animal for multiple analyses.
Pulverized hemisphere fractions were processed by homogenization in buffer containing 70 mmol/L sucrose, 210 mmol/L mannitol, 5 mmol/L HEPES (N-2-hydroxyethylpiperazine-N'-2-ehanesulfonic acide), 1 mmol/L EDTA (ethylenediaminetetraacetic acid) and protease inhibitors (van den Tweel et al, 2005a) using a Potter homogenizer (Heidolph, Schwabach, Germany). Homogenates were incubated on ice for 30 mins, followed by 10 mins centrifugation at 800g at 4°C, leading to a nuclear pellet (P1). Supernatants (S1) were collected and centrifuged at 10,000g for 10 mins at 4°C to obtain mitochondrion-free supernatant (S2) and a mitochondrial pellet (P2).
Nuclear proteins were obtained by homogenizing nuclear pellets (P1) in buffer containing 10 mmol/L HEPES (pH 7.9), 10 mmol/L KCl, 1.5 mmol/L MgCl2, 0.1 mmol/L EDTA, 0.1 mmol/L EGTA (ethyleneglycol tetraacetate) and 1 mmol/L dithiothreitol (DTT) supplemented with protease inhibitors using a Potter-homogenizer. After 15 mins incubation on ice, NP-40 was added (final concentration of 3.125%), the mixture was vortexed and centrifuged for 1 min at 10,000g at 4°C. The nuclear pellet was resuspended in ice-cold hypertonic nuclear extraction buffer containing 20 mmol/L HEPES (pH 7.9), 420 mmol/L NaCl, 1.5 mmol/L MgCl2, 1 mmol/L EDTA, 1 mmol/L EGTA, 1 mmol/L DTT and protease inhibitors, incubated on ice for 30 min and centrifuged for 5 mins at 10,000g at 4°C. The supernatant containing the nuclear proteins was collected and kept at −80°C until use.
Mitochondrial protein fraction was obtained by sonicating mitochondrial pellets (P2) in ice-cold buffer containing 50 mmol/L Tris, 5 mmol/L EDTA, 150 mmol/L NaCl and protease inhibitors. Homogenates were incubated on ice for 30 mins, centrifuged at 10,000 g for 15 min at 4°C and supernatants containing mitochondrial proteins were collected.
Western Blotting
Protein concentration was determined using a protein assay (BioRad, Hercules, CA, USA) with BSA as standard. Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Hybond-C, Amersham Biosciences, Roosendaal, The Netherlands). Equal protein loading was verified by Ponceau-S staining. Membranes were stained with mouse-anti-nitrotyrosine (Biomol, Plymouth, PA, USA), mouse-anti-HSP70 (Stressgen Biotech, Victoria, Canada), mouse-anti-cytochrome c (Pharmingen, San Diego, CA, USA), rabbit-anti-cleaved caspase 3 (Cell Signaling, Danvers, MA, USA) or rabbit-anti-AIF (Sigma-Aldrich) followed by incubation with goat-α-mouse-HRP (Jackson Immunoresearch, Cambridgeshire, UK) or donkey-α-rabbit-HRP (Amersham Biosciences). Specific bands were visualized by chemiluminescence detection (ECL, Amersham Biosciences) with X-ray film exposure. Films were scanned with a GS-700 Imaging Densitometer and analyzed with Quantity One Software (both BioRad).
Nitrite and Nitrate (NOx) Measurements
NOx in brain homogenates (S2 fraction) and neuronal cell culture supernatants were measured with fluorescent method using 2,3-diaminonaphthalene (DAN) (Sigma-Aldrich) adapted from Misko et al (1993). Since the presence of proteins interferes with fluorescence, samples were deproteinized by adding 30% ZnSO4 (1:20). Protein-free supernatants were incubated at room temperature (RT) with 0.2 mmol/L reduced nicotinamide adenine dinucleotide and 100 mU/mL Nitrate Reductase (Sigma-Aldrich) for 2 h. Next, 10 μL of DAN (50 mg/mL in 0.62 N HCl) was added to 100 μL sample and incubated at RT for 30 min. The reaction was stopped and fluorescence was maximalized by adding 20 μL 2.8 N NaOH. Formation of fluorescent product was measured in 96 wells FluoroNunc plates (Nunc) using a spectrofluori-plate reader with excitation at 355 nm and emission at 460 nm. Samples of sodium nitrite standard in corresponding buffer were measured to obtain a reference curve with a detection limit of 125 nmol/L nitrite.
Nitrite formation in macrophage culture supernatants was measured using Griess reagent (Promega, Madison, WI, USA), according to the manufacturer's instructions. Samples of sodium nitrite standard in RPMI 1640 medium were measured to obtain a reference curve.
In Vitro Effect of 2-Iminobiotin on Nitric Oxide Synthases Activity
SK-N-SH neuroblastoma cells (ATCC, Manassas, VA, USA) were cultured in 12-wells plates in a humidified (5% CO2) incubator at 37°C in Dulbecco's modified Eagle's minimum essential medium (DMEM)/Ham-F10 (1:1) supplemented with 10% FCS, 2 mmol/L L-Glutamine, 100U/mL penicillin and 100 mg/mL streptomycin (all culture media were from Gibco, Carlsbad, CA, USA). When confluency was reached, cells were washed twice with Gey's balanced salt solution (GBSS) supplemented with 25 mmol/L HEPES, 30 mmol/L glucose, 240 mg/L L-arginine and antibiotics. Cells were preincubated with different concentrations of 2-IB or L-NMMA in supplemented GBSS for 30 mins followed by depolarization with 50 mmol/L KCl for 30 mins. Hereafter, supernatants were taken for analysis of NOx with DAN fluorescent method.
Adult female rats were terminated by CO2 inhalation. Peritoneal lavage was performed by injecting 20 ml ice-cold RPMI 1640 medium (Gibco) into the peritoneal cavity, followed by gentle massage of the abdomen and drainage. Peritoneal cells (mainly macrophages) were washed twice in ice-cold RPMI 1640 supplemented with 5% fetal calf serum, antibiotics (100 mg/mL streptomycin, 100U/mL penicillin), 2 mmol/L glutamine and 0.1% β-mercaptoethanol. Cells were cultured (0.5 × 106 cells/mL) for 24 h at 37°C (humidified, 5% CO2) in 24-wells plates in the presence of 10 ng/mL lipopolysaccharide (LPS) (Sigma-Aldrich) and different concentrations of 2-IB or aminoguanidine (Sigma-Aldrich). After 24 h, supernatants were collected and used for nitrite analysis with Griess reagent.
Statistical Analysis
Mortality rates were analyzed with Fisher's exact test. Hemisphere area-, Western Blot- and nitrite or NOx data are presented as mean and s.e.m. Paired t-tests were used to analyze differences between contra- and ipsilateral hemispheres. Two-way ANOVA with Bonferroni post-tests was used to analyze treatment and gender effects. Histological scores are presented as individual data with median and were analyzed using Kruskal-Wallis tests. Mann-Whitney U tests were used for further analysis of 2-IB treatment and gender effects.
Results
Mortality
Total mortality in our experiments was 25.2% (29/115). Mortality rates differed significantly between genders; 37.3% (22/59) in male pups and 12.5% (7/56) in female pups (P < 0.01). The timing of mortality did not differ between male and female pups (male 18/22 during hypoxia, 4/22 during follow-up; female 5/7 during hypoxia, 2/7 during follow-up). There was no effect of treatment (vehicle or 2-IB) on mortality during follow-up. Mortality during follow-up occurred within the first week after HI, except for one rat (female) that died at day 24.
Preliminary evidence suggests that the animals that died during hypoxia did not have major brain damage. Moreover, in the surviving animals, the development of neuronal damage is a relatively slow process, which takes hours to days (van den Tweel et al, 2006). Therefore, we propose that cardiac or respiratory failure rather than brain damage is the primary cause of death in these animals.
Long-Term Effects of 2-Iminobiotin Treatment In Vivo
Area measurements of whole hemisphere and hippocampus were performed as depicted in Figure 1A. Hypoxia-ischemia induced a reduction in the area of the ipsilateral hemisphere or hippocampus, which was significantly attenuated in female but not male rats receiving 2-IB treatment compared with vehicle-treated rats (P < 0.01; Figures 1B and 1C). The reduction in the area of the ipsilateral hemisphere or hippocampus after HI was not different between vehicle-treated male and female rats. Additionally, histological examinations were performed in six regions of parietal cortex and five regions of hippocampus as depicted in Figure 2A. The neuroprotective treatment effect of 2-IB treatment was observed in female rat pups only (P < 0.01; Figure 2B), and treatment was not effective in male littermates. The neuroprotective effect of 2-IB was observed in both cortex (P < 0.01 versus vehicle) and hippocampus (P < 0.05 versus vehicle) of female rat pups (Figures 2C and 2D). In vehicletreated animals, no difference in severity of brain injury after HI between male and female rat pups was observed.
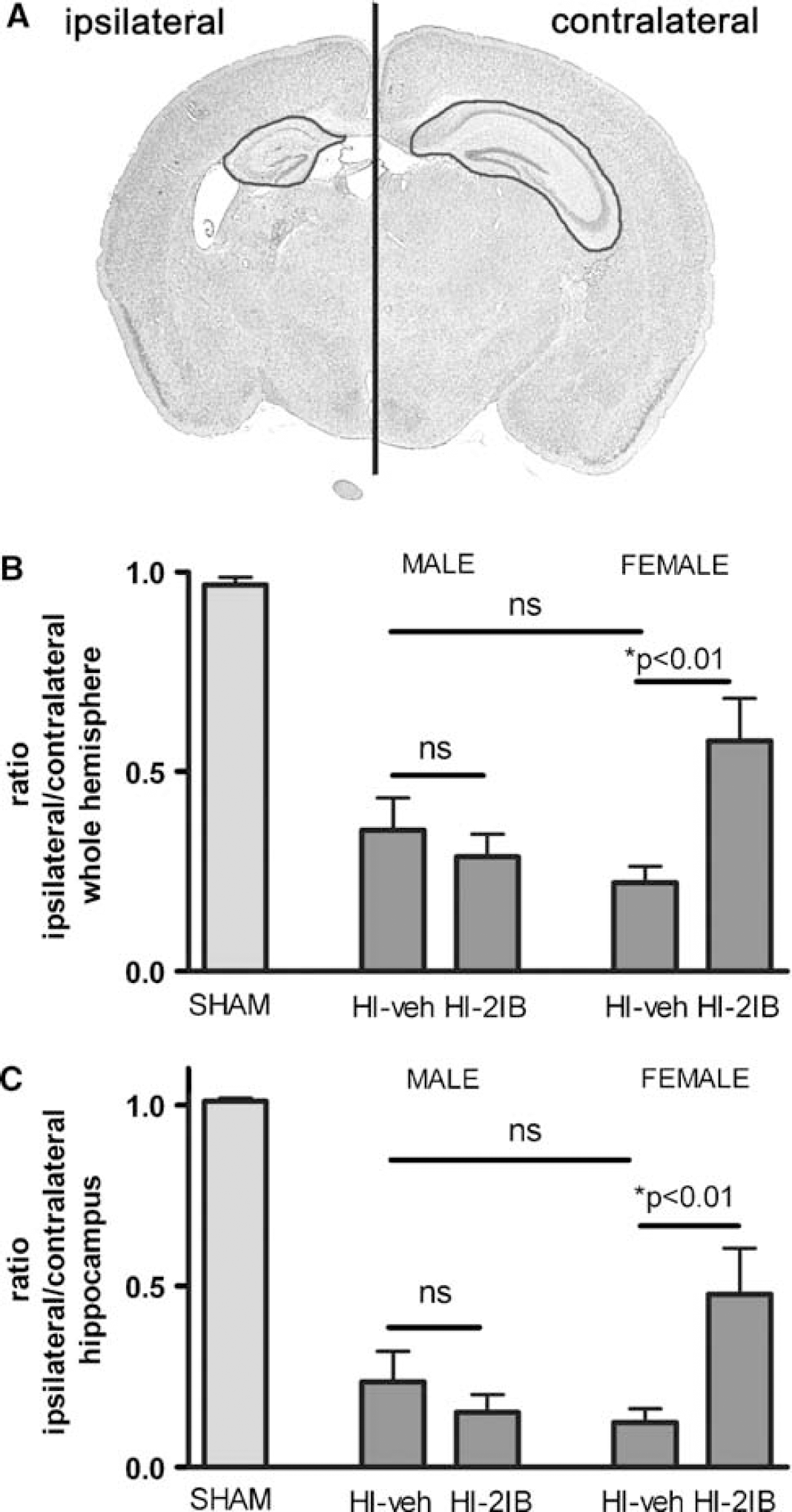
Long-term effects of 2-iminobiotin treatment on brain area loss in P7 rats. (
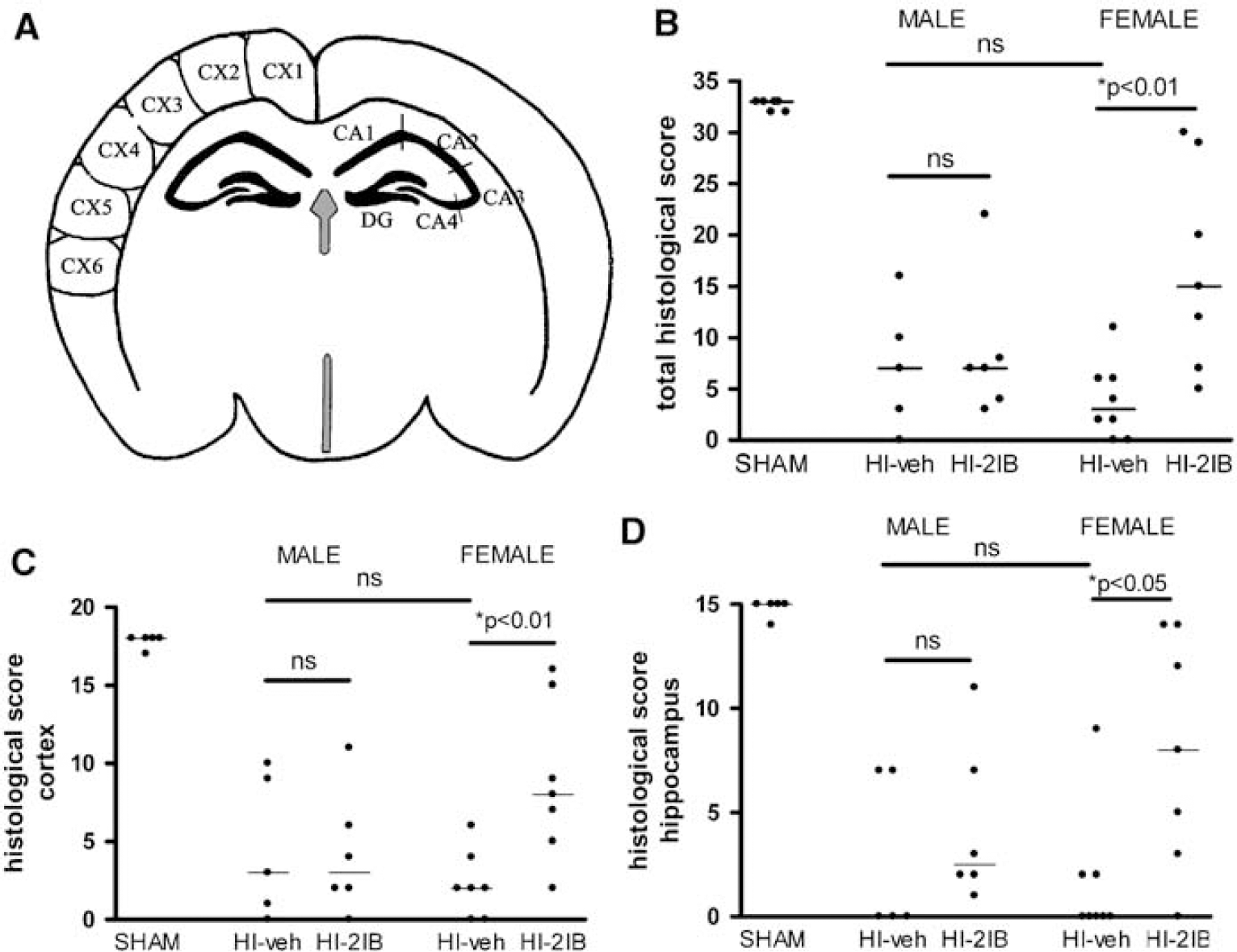
Long-term effects of 2-iminobiotin treatment on histological scores in P7 rats. Brain sections were stained with hematoxylin-eosin. (
Involvement of Inhibition of Nitric Oxide Production in the Effect of 2-Iminobiotin
In vivo: nitrite plus nitrate (NOx) and nitrotyrosine: Based on the described inhibitory effect of 2-IB on purified iNOS and nNOS in vitro, we have previously suggested that the neuroprotective effect of 2-IB was dependent on inhibition of nNOS and iNOS activity. To determine whether the neuroprotective effects of 2-IB indeed involve NOS inhibition, we now determined NOx level in brain homogenates of P7 rat pups at 24 h after the HI insult as an indicator of NO production. NOx levels were significantly increased after HI in the ipsilateral hemispheres in both female and male animals and no gender differences were observed. More importantly, 2-IB treatment did not have a significant effect on total NOx level in either gender (Figure 3A).
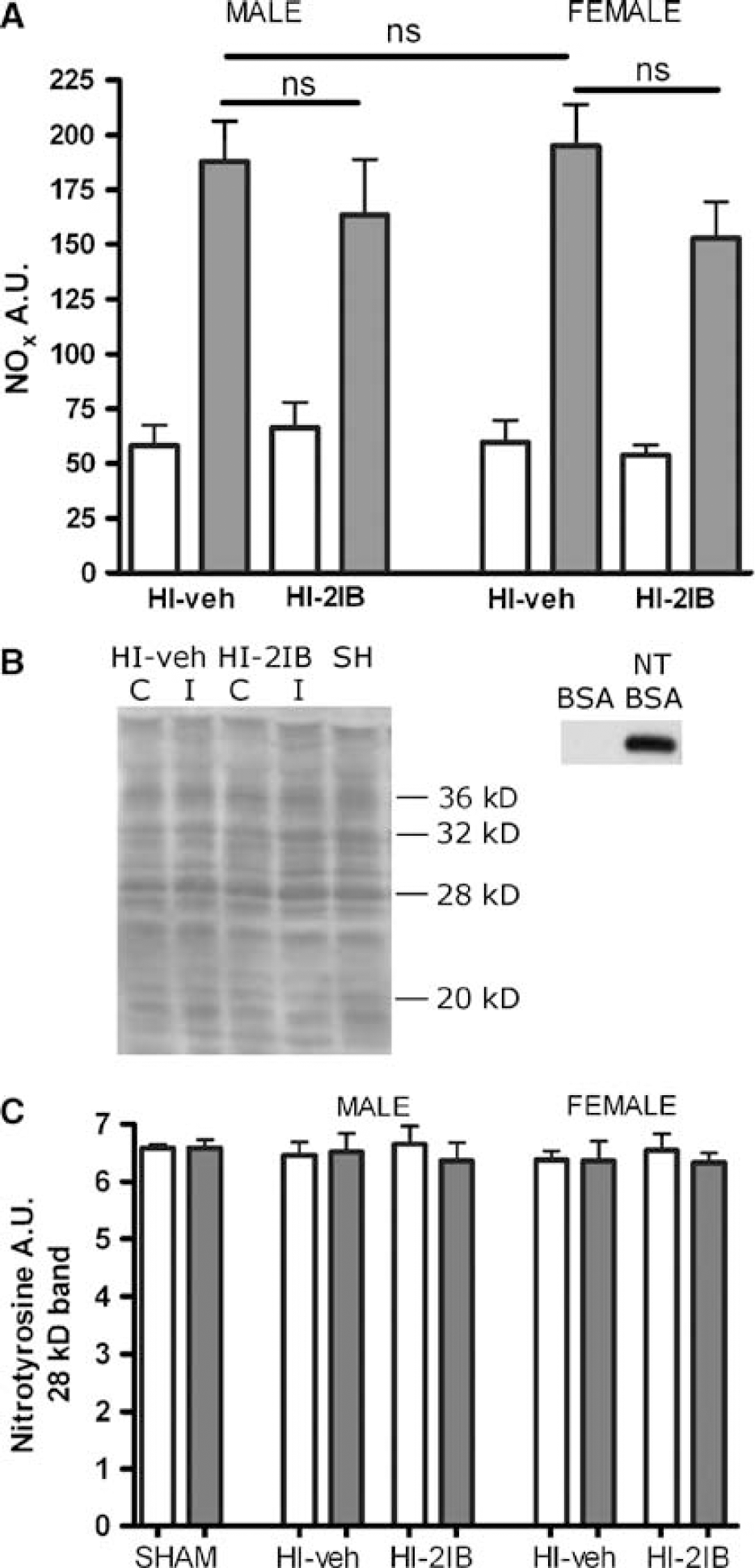
Effect of 2-iminobiotin on NOx and nitrotyrosine levels in the brain 24h post-HI in P7 rats. Animals were exposed to HI followed by 2-iminobiotin treatment. (
As an additional measure of the effect of 2-IB on NOS activity in the brain, we determined cerebral nitrotyrosine (NT) levels. In line with our earlier observations in the P12 rat (van den Tweel et al, 2005a, b ), we did not observe a HI-induced change in nitration of tyrosine residues in cerebral proteins after analyzing several nitrotyrosine bands at different molecular weights (Figures 3B and 3C). Moreover, no effect of 2-IB on brain NT levels or any effects of gender were observed (Figures 3B and 3C).
In vitro: inhibition of neuronal and inducible Nitric Oxide Synthases: In contrast to our expectations, the data from the in vivo studies mentioned above suggest that 2-IB does not protect females from neuronal injury via inhibition of NO production. To further address this issue, we also determined the capacity of 2-IB to inhibit nNOS activity in intact cells in vitro. As a model, we used a neuronal cell line of female origin (SK-N-SH cells). Cells were preincubated with increasing amounts of 2-IB or the well-described NOS inhibitor L-NMMA. Nitrix oxide production with NOx formation as a read-out was determined after depolarization with KCl. Figure 4A shows that 2-IB has a small inhibitory effect on NOx formation, that did not reach statistical significance even at the highest concentration used (maximal 33.8% inhibition with 1 mmol/L; P > 0.05). Equal volumes of vehicle solution (0.01 mol/L HCl) were tested and did not affect NOx formation (data not shown). In contrast, inhibition with L-NMMA showed already significant (77.7%) inhibition of NOx formation with the lowest concentration tested (50 mmol/L; P < 0.001) and complete inhibition with L-NMMA concentration between 250 and 500 mmol/L (Figure 4A).
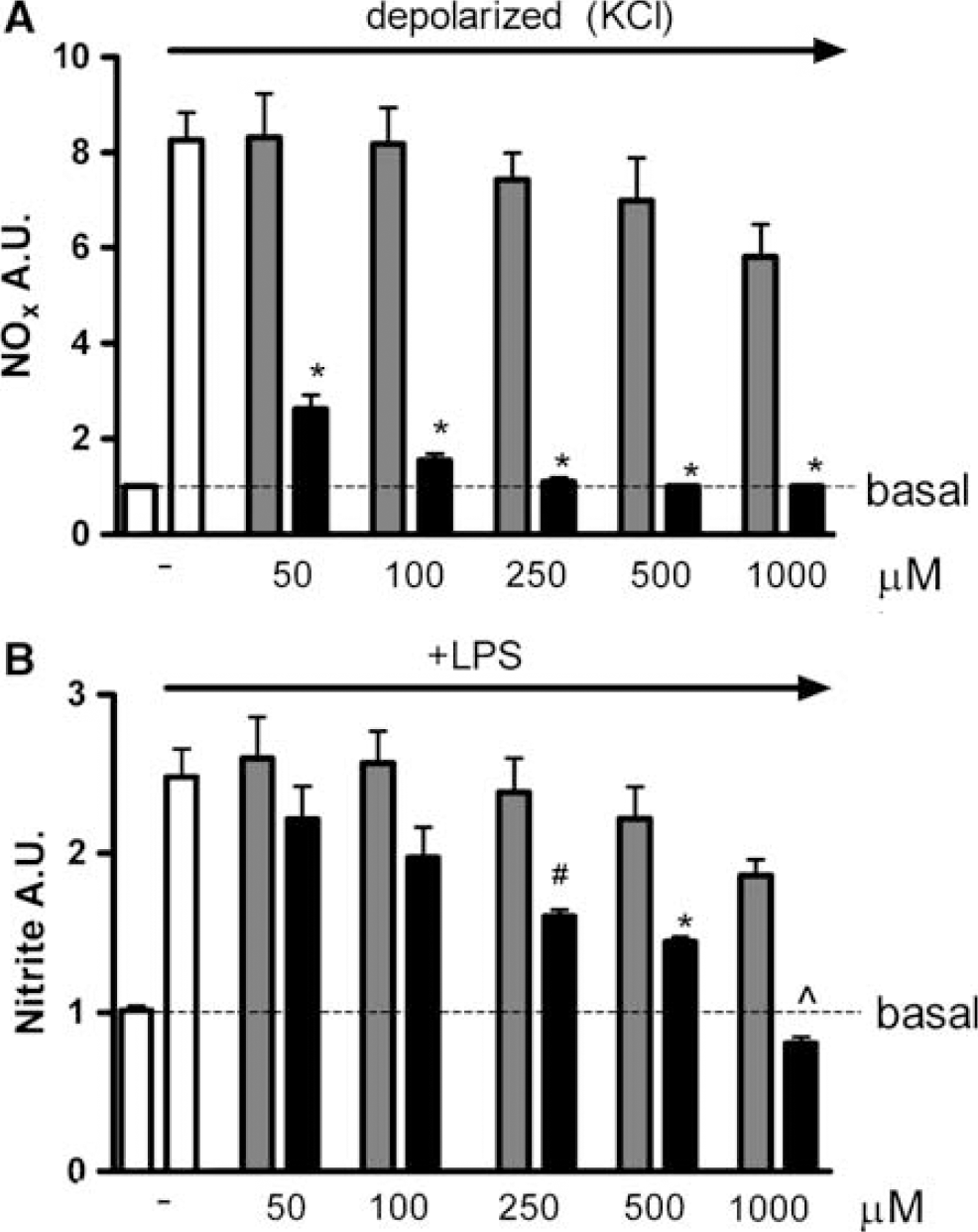
Effect of 2-iminobiotin, L-NMMA and aminoguanidine on NO production by cultured cells. (
Next, we compared the capacity of 2-IB to inhibit iNOS activity in LPS-stimulated macrophages with the iNOS-inhibitor aminoguanidine (AG). Incubation with 50 to 1000 mmol/L of 2-IB did not lead to a significant inhibition of nitrite formation (Figure 4B). In contrast, inhibition of NO production with AG was already statistically significant at 250 μmol/L (50.7%, P < 0.05) and NO production was prevented completely using 1 mmol/L AG (Figure 4B, P < 0.01). The same experiments were repeated in IMDM medium, which contains three times less arginine (NOS substrate) than RPMI medium. As expected, inhibition of nitrite formation was facilitated for all concentrations AG used (data not shown). In contrast, the lower concentration of arginine did not influence the effect of 2-IB on nitrite formation (data not shown).
Short-Term Effects of 2-Iminobiotin Treatment In Vivo: HSP70 as a Cell Stress Marker
The results from the analysis of the effect of 2-IB on NOx formation in vivo and in vitro suggest that the neuroprotective effect of 2-IB is not mediated via inhibition of NOS activity. To get more insight in the mechanism involved in the neuroprotective effect of 2-IB in female rats, we evaluated the HI-induced increase in HSP70 levels after the HI-insult as an early marker of brain damage. In preliminary experiments, we analyzed HSP70 expression at 24, 48 and 72 h after HI and observed a maximal increase at 48 h after HI (data not shown). Therefore, we examined HSP70 expression at 48 h. Hypoxia- ischemia induced a similar increase in HSP70 in female and male pups. However, 2-IB treatment only reduced HSP70 expression in female pups (P < 0.01; Figure 5). In both hemispheres of sham-operated pups and in the contralateral hemisphere of HI male and female rat pups, only a very low expression of HSP70 was detected.
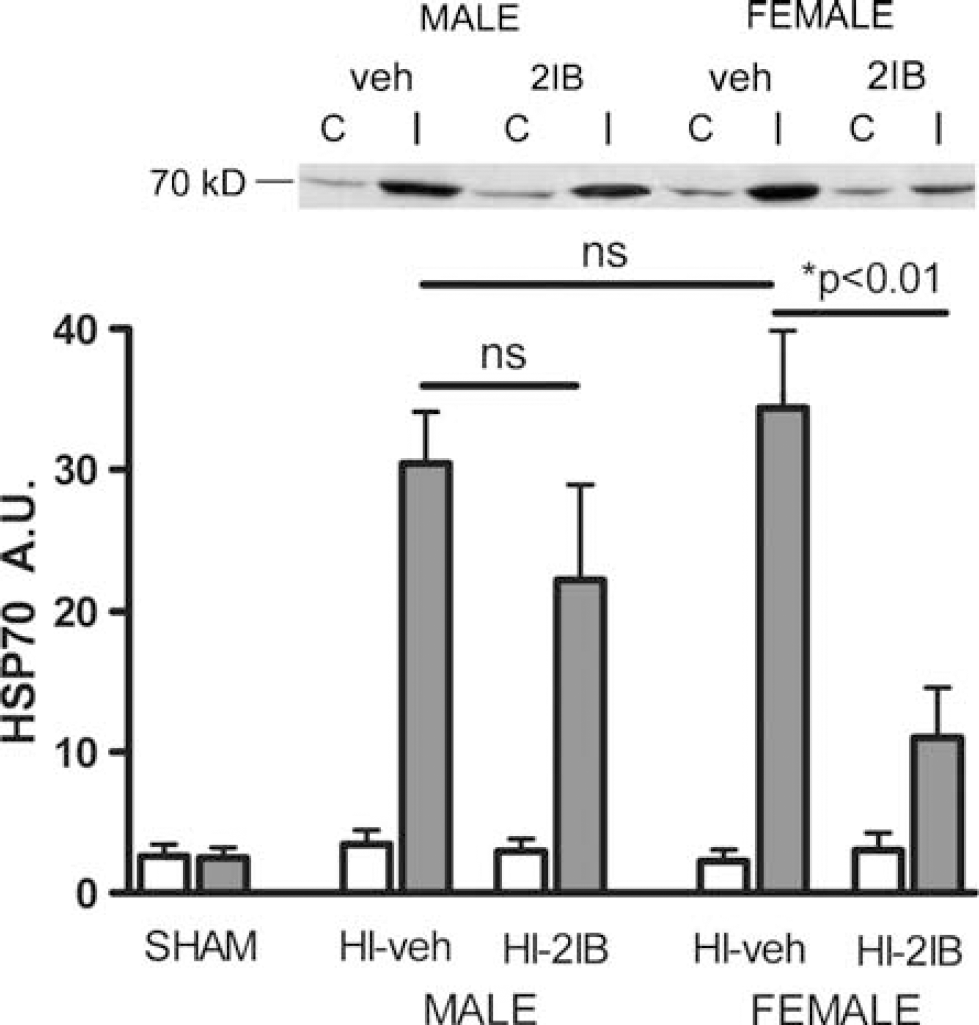
Effect of 2-iminobiotin on HSP70 expression at 48 h after hypoxia-ischemia in P7 rats. Animals were subjected to HI and ipsi- (gray bars) and contralateral (white bars) hemispheres were processed for Western blot analysis. Sham-operated (n = 5); males treated with vehicle (n = 7) or 2-iminobiotin (n = 7) and females treated with vehicle (n = 10) or 2-iminobiotin (n = 10). Inset shows representative blot. Data are represented as mean ± s.e.m.
Short-Term Effects of 2-Iminobiotin Treatment In Vivo: Caspase-Dependent and Caspase-Independent Cell Death Pathways
2-Iminobiotin treatment is neuroprotective in female and not in male rats and the protective effect can already be detected early after HI. Moreover, it has been shown in vitro and in vivo that female neurons predominantly use the cytochrome c-caspase 3 pathway of apoptosis whereas male neurons preferentially use the caspase-independent AIF pathway. Therefore, we next examined whether the gender-specific neuroprotective effect of 2-IB is associated with specific effects on the cytochrome c-caspase 3 pathway.
First, cytochrome c release from the mitochondria to the cytosol was determined at 24 h after HI in male and female P7 pups. The HI-induced increases in ipsilateral cytosolic cytochrome c levels were similar in vehicle-treated males and females. However, only in female pups, the increase in cytosolic cytochrome c level in the ipsilateral hemisphere after HI was significantly reduced after 2-IB treatment (P < 0.001; Figure 6A).
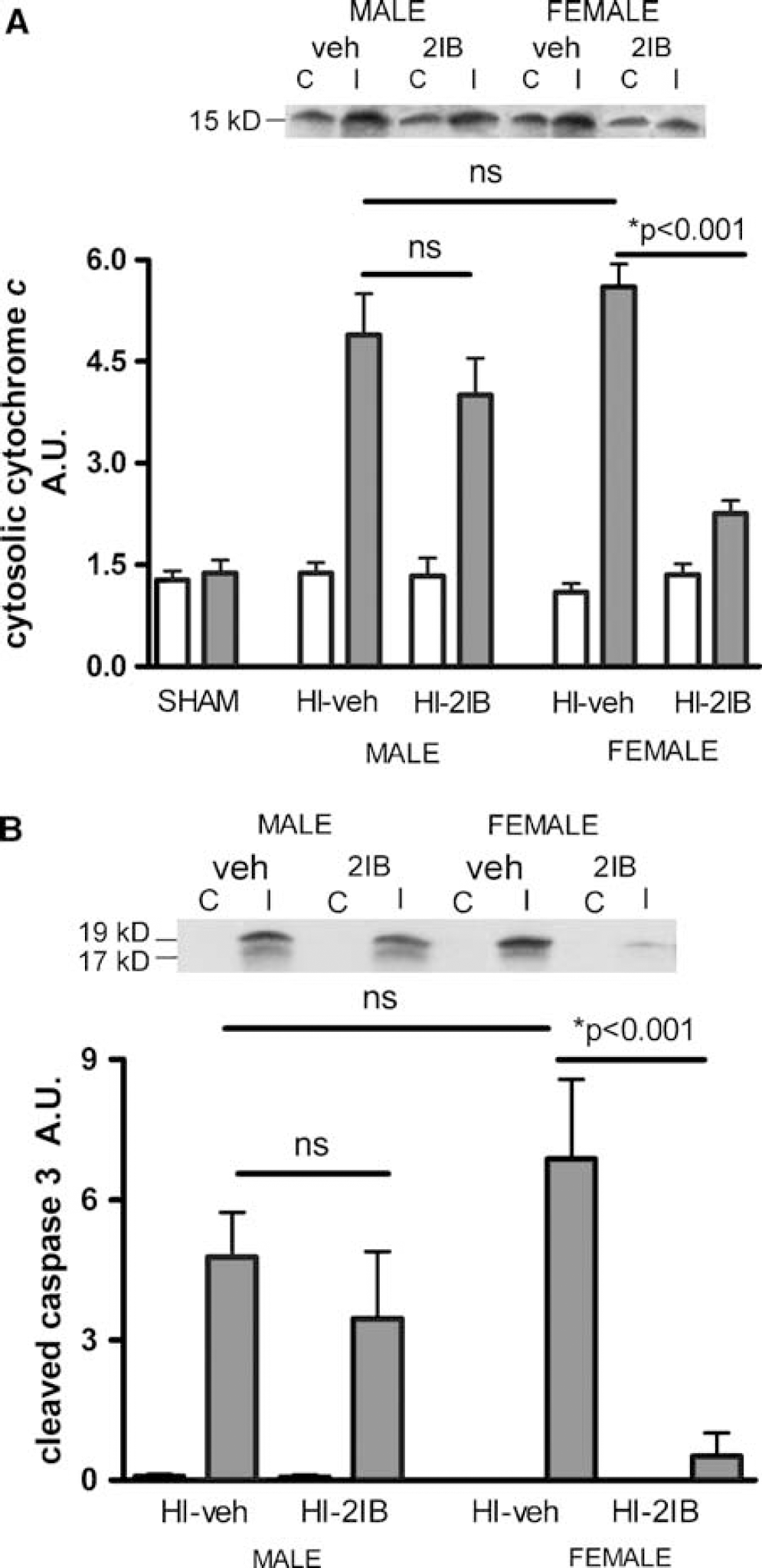
Effect of 2-iminobiotin on cytosolic cytochrome c and active (cleaved) caspase 3 at 24 h after hypoxia-ischemia in P7 rats. Animals were exposed to HI followed by 2-IB treatment and mitochondria free cytosolic preparations were subjected to Western blot analysis. White bars show levels in contralateral hemispheres and gray bars in ipsilateral hemispheres. (
Cleavage of inactive caspase 3 to form active (cleaved) caspase 3 is a downstream event of cytochrome c release and a crucial step in apoptotic cell death. To confirm the cytochrome c-caspase 3 pathway, we measured the expression of active (cleaved) caspase 3 at 24 h after HI. Levels of cleaved caspase 3 were not significantly different in the ipsilateral hemispheres of vehicle treated-male and female animals at 24 h after HI. 2-Iminobiotin treatment decreased cleaved caspase 3 levels in female animals only (P < 0.001; Figure 6B). Cleaved caspase 3 levels in both hemispheres of shamoperated animals or contralateral hemispheres of HI animals were undetectable.
The caspase-independent pathway of apoptosis is associated with mitochondrial to nuclear translocation of AIF. The level of AIF in mitochondrial, cytosolic and nuclear fractions of the ipsi- and contralateral hemisphere was determined at 24 h post-HI. We detected a single band of AIF at 57 kDa, indicating that the AIF we detect is of mitochondrial origin as newly synthesized AIF has a molecular weight of 67 kDa (Saelens et al, 2004). We observed significant HI-induced translocation of AIF from the mitochondria to the nucleus in the ipsilateral hemisphere in males but not in females: HI significantly reduced ipsilateral mitochondrial AIF (P < 0.05 versus contralateral; Figure 7A) and increased ipsilateral nuclear AIF (P < 0.001 versus contralateral; Figure 7B) in males but not in females. 2-Iminobiotin treatment after HI had no effect on HI-induced AIF translocation in males. Moreover, the minor translocation of AIF in females was not prevented by 2-IB treatment (Figures 7A and 7B). Apoptosis-inducing factor expression in cytosolic fractions was observed in males only (Figure 7C).
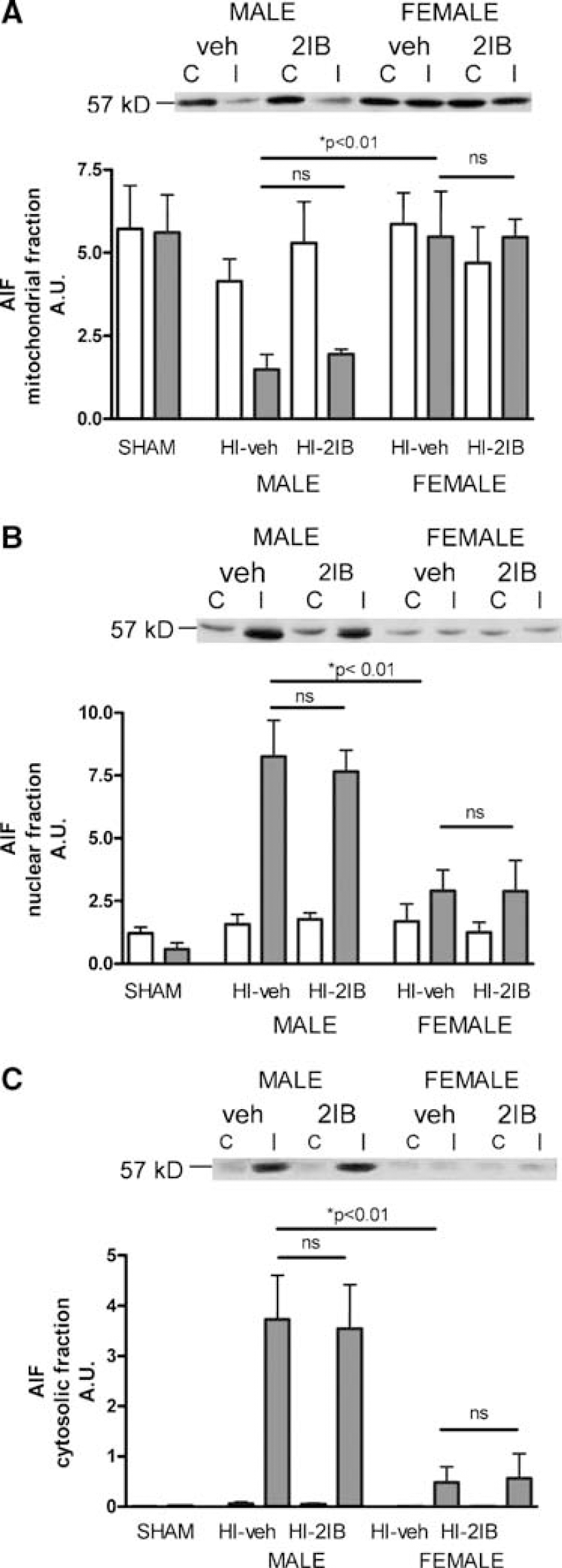
Effect of 2-iminobiotin on translocation of AIF at 24 h post-hypoxia-ischemia in P7 rats. Animals were exposed to HI followed by 2-IB treatment and mitochondrial (
Discussion
Recently, we have described short- and long-term neuroprotection by post-HI treatment with 2-IB in P12 rat pups as a model for the full-term human neonate (van den Tweel et al, 2005a). Here, we used P7 rat pups as a model for the human neonate born preterm with brain development comparable to human gestational age of 30 to 34 weeks of pregnancy (Romijn et al, 1991; Northington et al, 2005). Aim of the present study was twofold: to address the role of gender in determining the neuroprotective effect of 2-IB after HI in the neonatal rat and to further analyze the mechanism involved in the neuroprotective effect of 2-IB. We show here for the first time that 2-IB treatment provides long-term neuroprotection in P7 female rat pups after HI and not in male rat pups.
The finding that the neuroprotective effect of 2-IB treatment after HI is gender-dependent, prompted us to further investigate the mechanisms of action of 2-IB. It has been suggested that increased NO production by the inducible and neuronal form of NOS is an important mechanism in neonatal HI-induced brain damage. We have explored the neuroprotective capacity of 2-IB in neonatal rats and in piglets based on a study by Sup et al (1994), who described that 2-IB is capable of inhibiting the enzymatic activity of purified nNOS and iNOS in vitro. Therefore, we and others have previously suggested that the in vivo neuroprotective effect of 2-IB is mediated by inhibition of the activity of cerebral nNOS and iNOS (van den Tweel et al, 2005a; Peeters-Scholte et al, 2002; Zhu et al, 2004). However, the data presented here urged us to revise our hypothesis on the mechanism underlying the neuroprotective effect of 2-IB.
First of all, we show that HI induces an increase in NOx in the ipsilateral hemisphere in both males and females, implying that increased NO production does occur in neonatal HI-induced brain damage. However, we also show that 2-IB does not inhibit this HI-induced increase in NOx in female or male rats. In line with these observations, 2-IB did not alter the level of NT in the brain. Second, 2-IB is at best a poor inhibitor of NOS activity in intact (female) neurons or macrophages in vitro. Only at the maximum dose tested (1 mmol/L), we observed a small, but not statistically significant inhibitory effect of 2-IB on NO production in neuronal cells and in macrophages. In contrast, the specific iNOS inhibitor aminoguanidine results in a significant inhibition of NO production in macrophages at a much lower dose (0.25 mmol/L). In this respect, it is important to note that the neuroprotective dose of 2-IB, which is given in vivo, is very low in comparison to the dose of aminoguanidine that inhibits iNOS activity in vivo (2-IB: 0.12 mmol/kg versus AG: 3.25 mmol/kg). The results may seem contradictory to the recent finding that 2-IB inhibited nitration of proteins after HI (Zhu et al, 2004). However, protein nitration depends on peroxynitrite formation, which results from both production of superoxide radicals and NO and may therefore not accurately reflect production of nitric oxide.
In addition, it should also be emphasized that the neuroprotective effect of the well-described nNOS and iNOS inhibitors 7-nitroindazole and aminoguanidine that we have described previously in P12 rats was not gender specific (van den Tweel et al, 2002), showing that inhibition of NOS in the neonatal rat can be neuroprotective but is not gender specific. The gender-dependent neuroprotective effect of 2-IB was not restricted to the P7 rat. Re-analysis of our data in the P12 pups also shows that the neuroprotective effect of 2-IB occurred in the females only (data not shown). Similarly, we observed gender-dependent neuroprotection by 2-IB treatment after HI in P3 rats (Nijboer et al, unpublished data). Previous studies in adult models of ischemic brain injury have shown that treatment with the iNOS inhibitor aminoguanidine reduced infarct size in male, but not in female animals (Park et al, 2006). Similarly, it has been shown that nNOS−/− males or males treated with the nNOS inhibitor 7-nitroindazole are protected from ischemia-induced brain damage whereas female animals are not (McCullough et al, 2005). Conversely, those studies showed that inhibition of nNOS or nNOS deficiency increases infarction in females and iNOS deficiency did not alter damage in females. Thus, in adult animals, it appears that inhibition of iNOS or nNOS will protect males rather than females. The gender-specific effects of NOS inhibition in adult models of stroke has been attributed to estrogen-dependent effects (McCullough et al, 2005).
The question that should be addressed in the present study is: what is the mechanism of action of 2-IB that leads to neuroprotection in female but not male neonatal rats? Gender dimorphism in the effect of 2-IB was already observed when examining the early marker of brain damage HSP70 at 48 h after the insult. It is important to note that the gender-dependent effects of treatment were not associated with gender differences in the severity of cerebral injury after HI. In human adults, male sex is a risk factor for clinical stroke and this is linked to the protective effects of female reproductive hormones (Hurn and Brass, 2003; Hurn et al, 2005). Similarly, in animal stroke models, adult females are relatively protected against cerebral ischemia compared with males (Alkayed et al, 1998), and female reproductive hormones are responsible for the protective effects (Toung et al, 1998; McCullough and Hurn, 2003). Although in our experiments male pups had a significantly higher mortality rate during hypoxia, the surviving untreated males and females did not differ in NOx production, HSP70 expression or longterm damage (the latter in agreement with Hagberg et al, 2004; Zhu et al, 2006). In the neonatal rat, it is not likely that female sex hormones play a major role at P7 as endogenous estrogen production is not initiated before P12 and effects of endogenous estrogens will not emerge until 21 days of life (MacLusky et al, 1979). The influence of maternal estrogens in both females and males and endogenous estrogens in females will be low because of the presence of alpha-fetoprotein (AFP) in the neonatal circulation, which prevents circulating estradiol from entering (neuronal) cells (Wallen and Baum, 2002; Bakker et al, 2006). Finally, the level of estrogen is most likely to be higher in the male brain, where the enzyme aromatase (CYP19) catalyzes the local synthesis of estrogens from androgens, a process that is involved in brain masculinization (Ivanova and Beyer, 2000; Wallen and Baum, 2002). Interestingly, evidence from studies in adult animals suggests that estrogens (via binding to estrogen receptors) can modulate the activity of the antiapoptotic molecules Bcl-2 and Bcl-XL (Amantea et al, 2005). Unfortunately, we are not aware of any data in the literature describing gender differences in the activity of these antiapoptotic pathways. It could be possible however, that 2-IB via an unknown mechanism mimics the effects of estrogens that are not present in sufficient levels in the female brain at this age. If this is the case, one explanation for the lack of effect of 2-IB in the neonatal male brain could be that the presence of locally synthesized estrogens prevents an additional effect of 2-IB via this pathway.
Interestingly, using in vitro cultures of neuronal cells, innate gender differences in cell death pathways have been described (Du et al, 2004). In the in vitro situation, female (XX) neurons predominantly use the cytochrome c-caspase 3-dependent pathway of apoptosis after exposure to a number of cell death inducing agents. In contrast, male (XY) neurons show a proclivity for the use of the caspase-independent pathway of apoptosis that is characterized by PARP-1 activation and AIF translocation from mitochondria to the nucleus. Our present data show that also in vivo there is a gender difference in the use of the AIF pathway in the neonatal rat: in males, exposure to HI induced AIF translocation from the mitochondria to the nucleus, whereas in females we did not observe significant changes in the localization of AIF after HI. These data suggest that the PARP-1-AIF pathway associated with caspase-independent apoptosis is much more important in males than in females. This conclusion is supported by a study of Hagberg et al (2004) showing that PARP-1 deletion protects neonatal male, but not female mice from HI-induced cerebral injury.
Importantly, our data show that 2-IB treatment did not have any effect on AIF translocation in males and that the minor change in AIF translocation observed in females was not affected by 2-IB treatment. In accordance with our data, Zhu et al (2004) described that treatment with 2-IB did not alter the extent of AIF activation after HI, although the authors did not mention any gender specificity.
After HI, we observed an increase in cytosolic cytochrome c and cleaved caspase 3 in the ipsilateral hemisphere of both male and female rat pups after HI. Moreover, the increase in cytosolic cytochrome c and activation of caspase 3 was similar in males and females. Nevertheless, 2-IB treatment only reduces cytochrome c release to the cytosol and subsequent activation of caspase 3 in females and not in males. Correspondingly Zhu et al (2004) described a decrease in activation of caspase 3 after post-HI treatment with 2-IB; however, gender was not considered. The intriguing question is now why the increase in cytosolic cytochrome c is affected by 2-IB in females, but not in males. One possibility is that in both the male and female brain, exposure to HI induces translocation of cytochrome c and activation of caspase 3. 2-IB inhibits a pathway upstream of cytochrome c translocation to the cytosol in females that is not inhibited in males for unknown reasons, for example, because the upstream pathways differ between males and females, and therefore the cytochrome c release and caspase 3 activity in males are not affected by 2-IB.
A second explanation for the gender difference in the effect of 2-IB on cytosolic cytochrome c and caspase 3 activation therefore could be that in females, exposure to HI results in translocation of cytochrome c to the cytosol through the Bax/Bak pore and this process is inhibited by 2-IB. In males, HI primarily results in activation of the AIF/PARP pathway, which could induce further damage to the mitochondria, for example, mitochondrial outer membrane rupture (MOMP), resulting in secondary leakage of cytochrome c to the cytosol and subsequent activation of caspase 3 (Wang et al, 2004; Yu et al, 2002). If the AIF pathway is not inhibited by 2-IB, the MOMP model would explain why in the males 2-IB cannot prevent cytochrome c translocation to the cytosol.
In summary, 2-IB is a potent neuroprotective drug in female rats only. The neuroprotective activity of 2-IB does not involve inhibition of NO formation. Neonatal HI induces AIF translocation in males, whereas both males and female rats show increased cytosolic cytochrome c and cleaved caspase 3 levels. However, only in female rats, 2-IB does prevent the increase in cytosolic cytochrome c and activation of caspase 3.
Although the exact target of 2-IB in females remains to be determined, we propose that 2-IB is neuroprotective in female rats via inhibition of initiation of cell death pathways at a level upstream of the activation of cytochrome c-caspase 3 dependent apoptosis.
Footnotes
Acknowledgements
The premixed gas (8% oxygen) used in this study was kindly provided by HoekLoos Medical BV, Eindhoven, the Netherlands. The authors would like to thank Marlies Gouw, Alette Faber and Sander Bakker for their excellent technical assistance.