
Abstract
The catecholamine, dopamine (DA), is synthesized from 3,4-dihydroxy-L-phenylalanine (L-DOPA) by aromatic L-amino acid decarboxylase (AADC). Dopamine metabolism is regulated by monoamine oxidase (MAO) and catechol-O-methyltransferase (COMT). To measure dopaminergic metabolism, we used microdialysis with radiometric detection to monitor L-[β-11C]DOPA metabolites in the extracellular space of the rat striatum. We also evaluated the effects of AADC, MAO, and COMT inhibitors on metabolite profiles. The major early species measured after administration of L-[β-11C]DOPA were [11C]3,4-dihydroxyphenylacetic acid ([11C]DOPAC) and [11C]homovanillic acid ([11C]HVA) in a 1:1 ratio, which shifted toward [11C]HVA with time. An AADC inhibitor increased the uptake of L-[β-11C]DOPA and L-3-O-methyl-[11C]DOPA and delayed the accumulation of [11C]DOPAC and [11C]HVA. The MAO and COMT inhibitors increased the production of [11C]3-methoxytyramine and [11C]DOPAC, respectively. These results reflect the L-DOPA metabolic pathway, suggesting that this method may be useful for assessing dopaminergic metabolism.
Introduction
Dopamine (DA) is a catecholamine involved in neuropsychiatric disorders such as Parkinson's disease (Hornykiewicz and Kish, 1987), schizophrenia (Lindstrom et al, 1999), Alzheimer's disease (Itoh et al, 1996), epilepsy (Smolders et al, 1997), attention deficit hyperactivity disorder (Ernst et al, 1998), as well as drug dependence (Smith et al, 2006). Dopamine is generated from 3,4-dihydroxy-L-phenylalanine (L-DOPA) by aromatic L-amino acid decarboxylase (AADC). The L-DOPA is 3-O-methylated to L-3-O-methyl-DOPA (3-OMD) through catechol-O-methyltransferase (COMT). 3,4-Dihydroxyphenylacetic acid (DOPAC) and 3-methoxytyramine (3-MT) are metabolized from DA through monoamine oxidase (MAO) and COMT, respectively, and both are converted to homovanillic acid (HVA) as the final metabolite (Ashcroft, 1969; Eisenhofer et al, 2004a) (Figure 1). These metabolic enzymes regulate DA concentration, and many treatments for Parkinson's disease target their activity (Di Stefano et al, 2009). Monitoring L-DOPA metabolism can help evaluate Parkinson's disease and treatment effects.
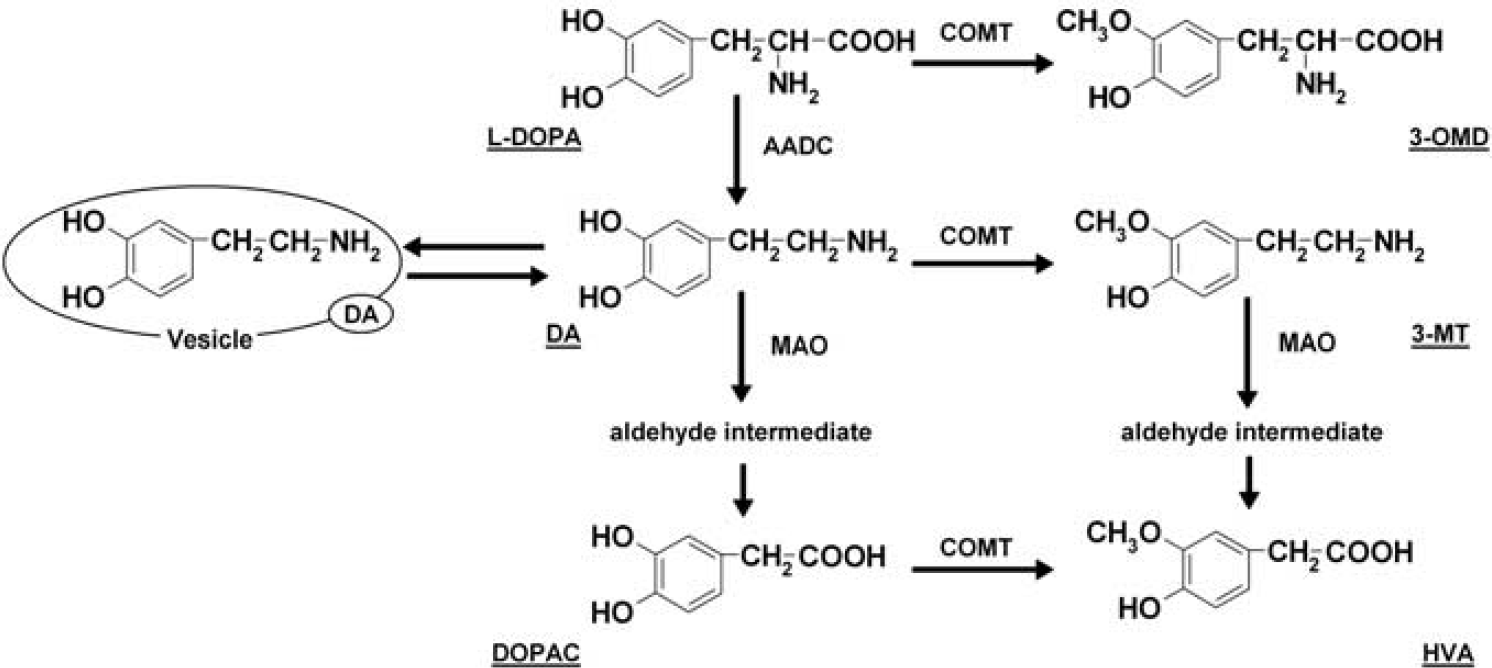
Metabolic pathway of 3,4-dihydroxy-L-phenylalanine (L-DOPA). AADC, L-amino acid decarboxylase; COMT, catechol-O-methyltransferase; DA, dopamine; DOPAC, 3,4-dihydroxyphenylacetic acid; HVA, homovanillic acid; MAO, monoamine oxidase; 3-MT, 3-methoxytyramine; 3-OMD, L-3-O-methyl-DOPA.
Positron emission tomography (PET) signals indicate the sum of radioactivity of a radioprobe and radioactive metabolites from the intracellular, extracellular, and intravascular compartments. However, PET cannot provide insight into the chemical composition or compartments of the detected species. 11C-labeled L-DOPA (L-[β-11C]DOPA) and 18F-labeled analogs of L-DOPA (6-[18F]-fluoro-DOPA) are used as biochemical probes to assess DA synthesis with PET (Ito et al, 2006; Ruottinen et al, 1997). However, the kinetics of L-[β-11C]DOPA and 6-[18F]-fluoro-DOPA are complicated by the presence of metabolites in brain and plasma, especially brain-penetrating metabolites. Accurate quantitative analysis of the kinetics of L-[β-11C]DOPA and 6-[18F]-fluoro-DOPA therefore requires compartmental modeling of several physiological processes in brain and plasma. Microdialysis coupled with liquid chromatography (MD-LC) or capillary electrophoresis can provide continuous monitoring of cerebral DA and its metabolites in extracellular fluid (ECF), and even provide structural information under appropriate conditions (Jonkers et al, 2001; Napolitano et al, 2003; Shou et al, 2006; Silva et al, 1997). Fast-scan cyclic voltammetry can also measure changes in DA concentration in freely moving animals on a subsecond time scale, but cannot differentiate molecules with similar structures by electrochemistry only (Robinson et al, 2003). Radiometric probes may be useful tools to measure the pharmacokinetics of L-DOPA and its metabolites in extracellular compartments using microdialysis, both to validate the quantitative analysis of PET studies and to evaluate the therapies for dopaminergic metabolism-related disorders.
The PET probes have limited usefulness in traditional microdialysis experiments because of the short half-life of the probes (11C: 20.4 minutes, 13N: 9.97 minutes, 15O: 2.04 minutes, 18F: 110 minutes) and extremely low radioactivity (typically <100 Bq) of target analytes in the dialysate (DeJesus et al, 2000; Lindner et al, 1995; Miwa et al, 1992; Tsukada et al, 1994). We recently developed a method combining online microdialysis with ultra-high performance liquid chromatography β+ detection (radio-LC) (Nakao et al, 2008; Okada et al, 2009) to continuously monitor extracellular radioactive metabolites of the PET probe. This method allows (1) online monitoring, (2) simultaneous measurement of endogenous and radioactive metabolites, and (3) comparison between different areas in the same animal.
Here, we established a method to assess dopaminergic metabolism in rats by radiometric monitoring of L-[β-11C]DOPA metabolites in ECF. We evaluated the effects of AADC, MAO, and COMT inhibitors on the metabolic profiles and compared the results with endogenous monitoring of the microdialysate.
Materials and methods
Chemicals and Synthesis of L-[β-11C]DOPA
The L-DOPA, DA hydrochloride, 3-OMD, 3-MT hydrochloride, DOPAC, HVA, 5-hydroxyindolacetic acid (5-HIAA), norepinephrine, benserazide, pargyline, and 3,5-dinitrocatechol were purchased from Sigma-Aldrich (St Louis, MO, USA). The L-[β-11C]DOPA was synthesized using a multi-enzymatic procedure through [11C]methyl iodine and DL-[β-11C]alanine as described earlier (Ikemoto et al, 1999; Sasaki et al, 2000). The radiochemical purity and specific radioactivity of L-[β-11C]DOPA were >98% and 43 to 103 GBq/μmol, respectively, at the time of administration.
Ethics of Animal Procedures
Animal procedures were performed according to the local regulation (Regulations for the Animal Experiments in the National Institute of Radiological Sciences (Chiba, Japan); 30 October 2008). The animal experimental protocol was approved by the Experimental Animal Committee of the National Institute of Radiological Sciences (Chiba, Japan).
In Vivo Microdialysis
Male Sprague-Dawley rats (250 to 350g; Japan SLC, Shizuoka, Japan) were housed under a 12-hour light cycle (07:00 to 19:00 hours) and allowed free access to food and water. Rats received an injection of sodium pentobarbital (50 mg/kg, i.p.) and, if necessary, an additional dose was injected. Lidocaine (2%, 0.6 mg) containing epinephrine (1:80,000) was used as a local anesthetic and rats were placed in a stereotaxic frame. A guide cannula (AG-8, Eicom, Kyoto, Japan) was implanted into the right striatum (A: 0.2 mm, L: 3.0 mm, D: 3.5 mm from the bregma) (Paxinos and Watson, 2004) through a hole drilled in the skull and fixed with two anchor screws and dental cement. At least 2 days after surgery, a microdialysis probe with a membrane length of 3 mm and a cutoff at 50,000 (AI-8-3, Eicom) was carefully inserted through the guide cannula. A cannula was inserted into the tail vein under diethyl ether anesthesia. Rats were moved into an acrylic box and allowed to recover from anesthesia. The probe was perfused at a flow rate of 4 μL/min with Ringer solution (Na+: 147 mmol/L, K+: 4 mmol/L, Ca2+: 2.3 mmol/L, Cl−: 155.5 mmol/L). Before experiments, in vitro recoveries of the microdialysis probe for L-DOPA and its metabolites were determined: all probes had recoveries of 9.8% to 13.7% at a flow rate of 4 μL/min. After a stabilization period (about 2 to 3 hours), rats received an intraperitoneal injection of an AADC inhibitor (benserazide, 25 mg/kg), MAO inhibitor (pargyline, 70 mg/kg), or COMT inhibitor (3,5-dinitrocatechol, 30 mg/kg). Saline (0.9%, i.p.) was administered as a control. Thirty minutes after drug treatment, 158 to 353 MBq (1.99 to 7.52 nmol) of L-[β-11C]DOPA was injected intravenously through a catheter. Brain dialysate was collected every 10 minutes and analyzed. Femoral artery dialysis was also conducted with a blood vessel microdialysis probe (membrane length, 10 mm; cutoff, 50,000; in vitro recoveries, 17.2% to 23.6%; TP-100-10, Eicom) in the group treated with benserazide. Plasma dialysate was collected every 1.5 minutes and analyzed.
Liquid Chromatography Conditions for the Metabolite Study of L-[β-11C]DOPA
We recently developed an online microdialysis radio-LC method for metabolite analysis of PET probes (Nakao et al, 2008). This system consists of an X-LC pump (3185PU, Jasco, Tokyo, Japan), a six-port online injection valve (HV-2080-01, Jasco) equipped with a 50-μL sample loop, a column oven (3167CO, Jasco), and a β+ detector (Oyokouken, Tokyo, Japan) housed in a shield with 50-mm thick lead and an electrochemical detector (ECD-300, Eicom). The applied potential between a glassy carbon working electrode and a reference Ag/AgCl electrode was set at 0.75 V. The mobile phase consisted of 100 μmol/L sodium phosphate buffer (pH 2.05), 0.4 μmol/L sodium dodecyl sulfate, and 0.2 μmol/L Na2EDTA containing 20% acetonitrile. Chromatographic separations were performed on an ACQUITY UPLC BEH C18 column (2.1 mm I.D. × 3.0 mm + 2.1 mm I.D. × 5.0 mm, 1.7 μm; Waters, Milford, MA, USA) for the striatum and on an ACQUITY UPLC BEH C18 column (2.1 mm I.D. × 5.0 mm, 1.7 μm; Waters) for plasma. The column temperature was set at 40 °C. The flow rates of the mobile phase for the striatum and plasma were set at 0.32 and 0.4 mL/min, respectively. Under these conditions, all analytes of interest were eluted within 5 minutes for the striatum and 1.5 minutes for plasma. The integrated counting time of the β+ detector was 3 seconds, and the flow cell volume for striatum and plasma were 25 and 10 μL, respectively. Striatal dialysate fractions obtained 0 to 10, 10 to 20, 20 to 30, 30 to 40, and 40 to 50 minutes after administration of L-[β-11C]DOPA were analyzed automatically.
Calculations
Data are expressed as mean ± standard error of the mean. The horizontal axis indicates the median time of each fraction. Radioactivity was corrected for physical decay of 11C to the time of administration of L-[β-11C]DOPA. In vitro probe recoveries for dopaminergic metabolites do not entirely correlate with in vivo ones, and we could not obtain these in vivo recoveries, so relative concentrations are presented. 11C radioactivity was expressed as the percentage of injected dose per dialysate fraction (percentage injected dose/40 μL). The statistical significance of the effects of each inhibitor for the time–radioactivity curves was determined by repeated measures analysis of variance. Area under the curve for 0 to 45 minutes (AUC0 to 45) was calculated for each time–radioactivity curve, and AUC0 to 45 was analyzed by two-tailed paired t-tests. Changes in endogenous metabolite concentrations after treatments compared with baseline levels were analyzed by one-way analysis of variance with Fisher's protected least significant difference test.
Results
Measurement of L-[β-11C]DOPA and Its Radioactive Metabolites in Extracellular Fluid
Figures 2A and 2B show typical radiochemical and electrochemical chromatograms, respectively, obtained with a striatal dialysate collected 30 to 40 minutes after intravenous administration of L-[β-11C]DOPA in the control group. The peaks of [11C]DOPAC, [11C]HVA, L-[β-11C]DOPA, and [11C]3-OMD, which eluted at 1.1, 1.3, 1.5, and 2.3 minutes, respectively, were identified on the basis of the retention time of their standards. An unidentified 11C-labeled metabolite ([11C]unknown, retention time: 0.9 minutes) was detected in all samples, but [11C]DA (retention time: 2.7 minutes) could not be detected in any of the samples. The time course of the extracellular radioactivity of L-DOPA and its metabolites from rats treated with saline is shown in Figure 3A, and the time course of endogenous concentration of L-DOPA and its metabolites from rats treated with saline is shown in Figure 4A. Saline treatment did not affect endogenous L-DOPA metabolite profiles (DOPAC, HVA, L-DOPA, 3-OMD, DA, and 3-MT). In the first fraction (0 to 10 minutes after the administration of L-[β-11C]DOPA), 43.7 ± 0.8% of total radioactivity was derived from [11C]DOPAC and 41.9 ± 6.7% from [11C]HVA. The remainder of the radioactivity was from [11C]unknown, L-[β-11C]DOPA, [11C]3-OMD, and [11C]3-MT. Subsequently, the radioactivity due to [11C]HVA, [11C]unknown, and [11C]3-OMD gradually increased whereas that due to [11C]DOPAC decreased.
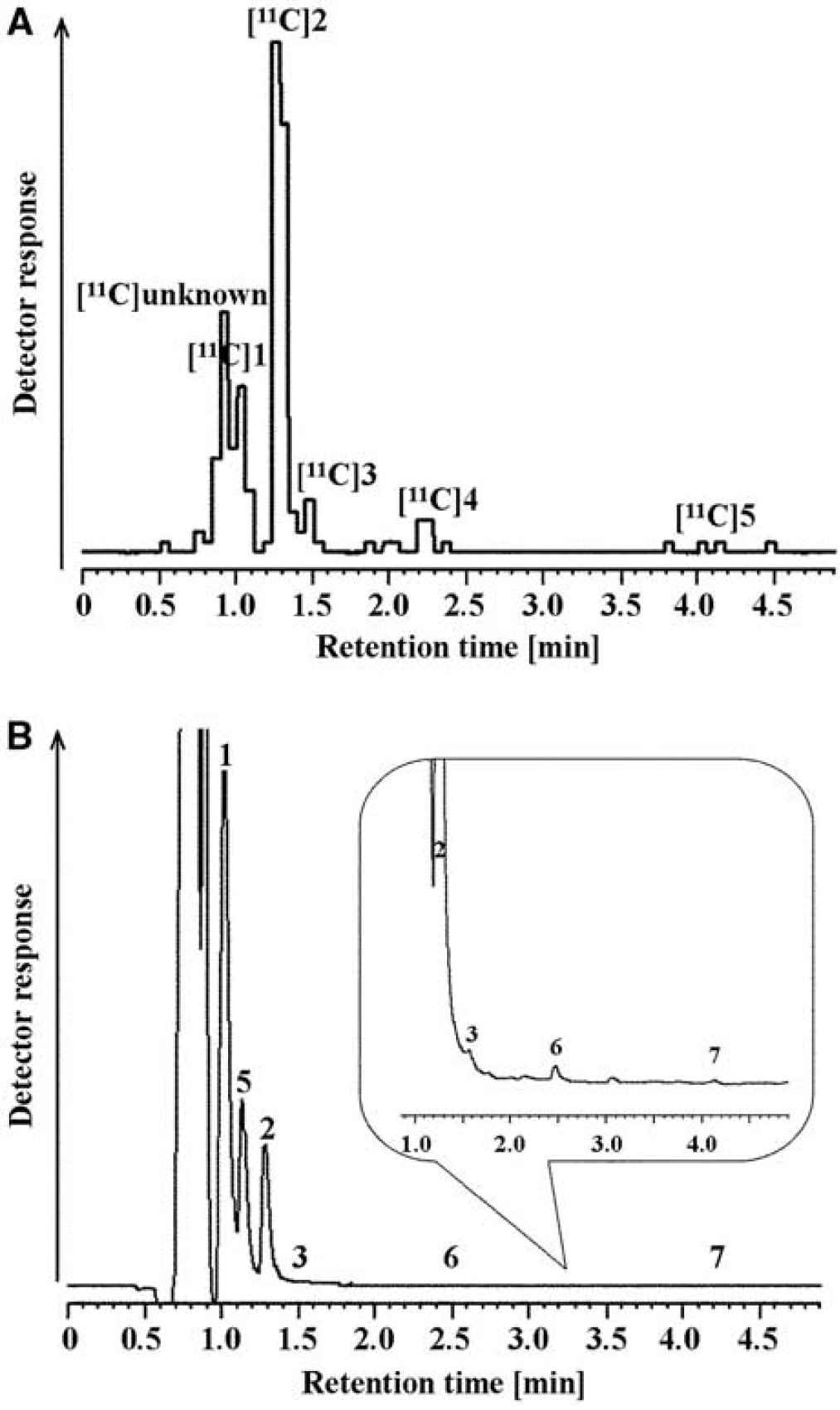
Typical (
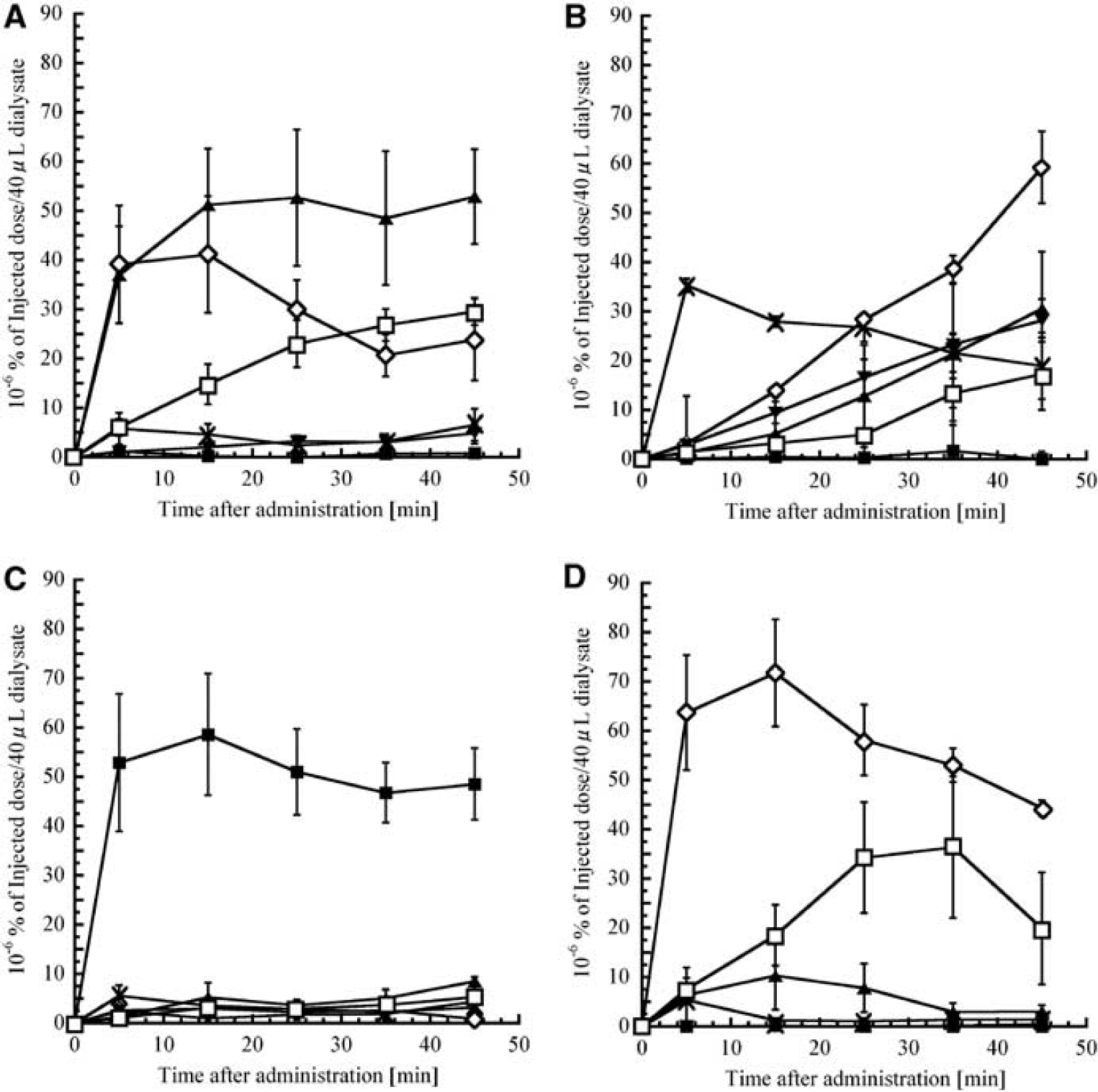
Time-radioactivity curves of L-DOPA and its metabolites obtained from striatal dialysate. Rats were treated with (
Time courses of endogenous L-DOPA and related metabolites obtained from striatal dialysate. Rats were treated with (
Effect of L-Amino Acid Decarboxylase Inhibition with Benserazide
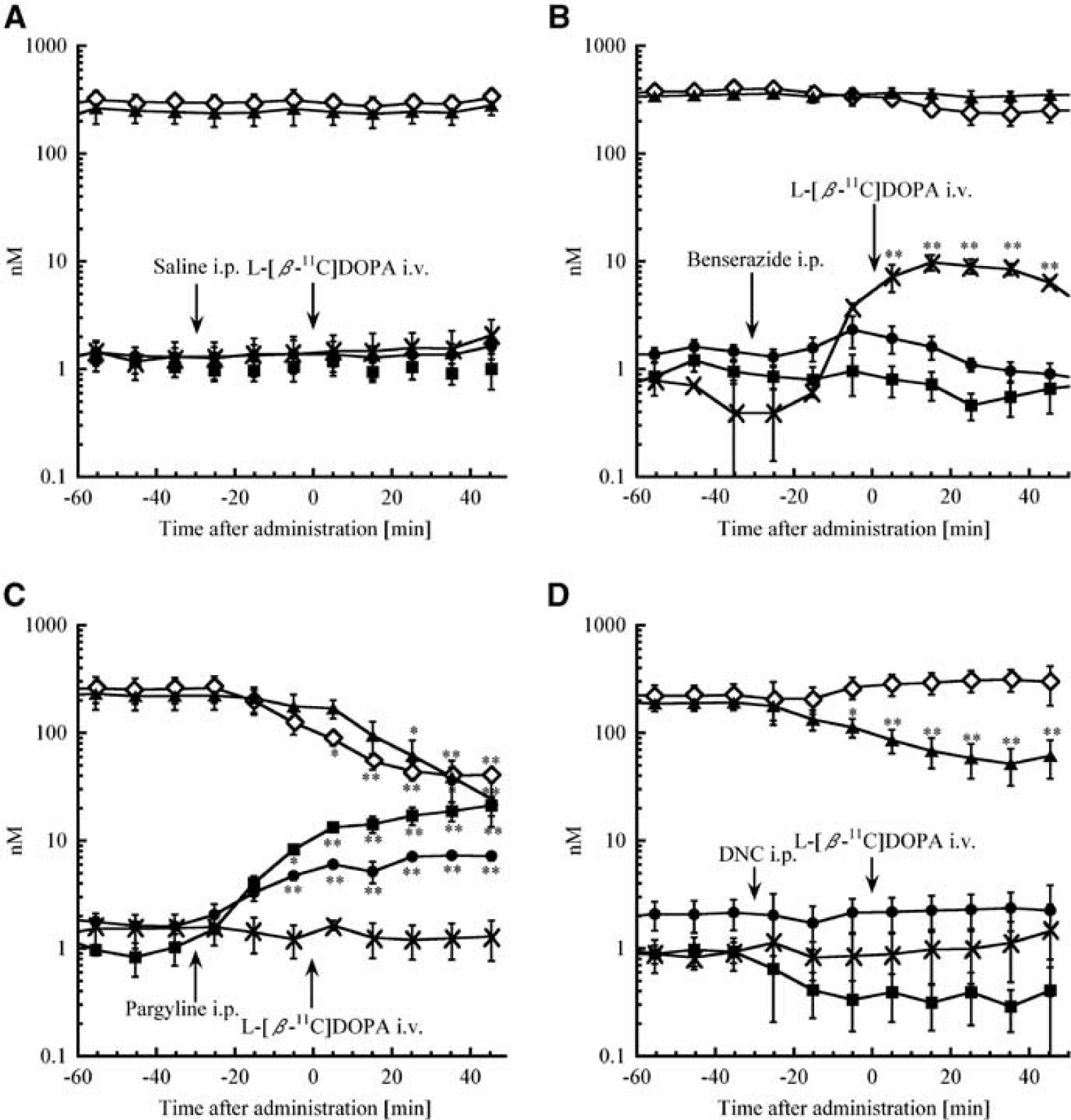
Benserazide increased L-[β-11C]DOPA and [11C]3-OMD levels compared with saline treatment (Figure 3B), and [11C]DOPAC, [11C]HVA, and [11C]unknown showed a delayed increase. The AUC0 to 45 of L-[β-11C] DOPA and [11C]3-OMD significantly increased to 540 ± 280% and 510 ± 150%, respectively (P < 0.01 and P < 0.05, respectively), and that of [11C]HVA and [11C]unknown significantly decreased to 23 ± 3.9% and 34 ± 3.7%, respectively (P < 0.05 and P < 0.01, respectively). Benserazide also increased endogenous L-DOPA up to 9.82 ± 1.42 nmol/L (Figure 4B).
Effect of Monoamine Oxidase Inhibition with Pargyline
With MAO inhibition, about 80% of total radioactivity (50 × 10−6% of injected dose/40 μL) was derived from [11C]3-MT throughout the experiment (P < 0.01), and the accumulation of [11C]DOPAC, [11C]HVA, and [11C]unknown was much lower than controls (each P < 0.05) (Figure 3C). The AUC0 to 45 values were also different (each P < 0.05), and endogenous measurements showed these tendencies: DOPAC decreased followed by reduced HVA but increases in 3-MT (Figure 4C). Although pargyline increased endogenous DA levels fivefold, radioactive [11C]DA was not detected.
Effect of Catechol-O-Methyltransferase Inhibition with 3,5-Dinitrocatechol
3,5-Dinitrocatechol increased [11C]DOPAC (P < 0.05) and decreased [11C]HVA (P <0.05) levels, with no [11C]3-OMD detected (P < 0.05) (Figure 3D). The AUC0 to 45 values were also different (each P < 0.05), except for [11C]unknown. 3,5-Dinitrocatechol decreased endogenous HVA from 197 ± 10 to 81 ± 9.0 nmol/L and 3-MT from 0.91 ± 0.14 to 0.35 ± 0.06 nmol/L, but did not change endogenous DOPAC (Figure 4D).
Discussion
We could obtain the biochemical kinetics of both 11C-labeled and endogenous L-DOPA and related metabolites simultaneously for 50 minutes after administration of L-[β-11C]DOPA, with a 10-minute resolution. Chromatographic separation of these compounds was completed within 5 minutes, which is significantly faster than earlier methods (Adachi et al, 2005; Jung et al, 2006; Napolitano et al, 2003; Niazi Shahabi et al, 2003; Silva et al, 1997). We could therefore perform studies on two rats in parallel using one chromatograph system combined with two six-port online injection valves, with a 5-minute time lag for each analysis. The predominant 11C-radioactivity was in the form of [11C]DOPAC and [11C]HVA in a 1:1 ratio soon after the administration of L-[β-11C]DOPA. Later, the ratio shifted toward [11C]HVA (Figure 3A). These results agree with the metabolite pathway and brain clearance of L-DOPA (Figure 1), which could not be clearly obtained from endogenous monitoring (Figure 4A).
We could not detect the [11C]DA fraction in any dialysate, potentially indicating that (1) [11C]DA is present mainly in synaptic vesicles; (2) released extracellular [11C]DA is immediately removed by metabolism, re-uptake, and binding; and (3) newly synthesized DA ([11C]DA) is not immediately released into the extracellular space. In particular, recaptured DA and leaked DA from vesicles into the cytoplasm is subsequently metabolized by MAO within neurons (Eisenhofer et al, 2004b), hence we could detect 11C-labeled DA metabolites even though we could not detect [11C]DA. The extracellular concentrations of acidic metabolites such as DOPAC and HVA are much higher than the parent transmitter, DA (Adachi et al, 2005; Napolitano et al, 2003); monitoring of these radioactive metabolites in ECF could therefore provide useful information about dopaminergic metabolism.
Inhibitors of AADC, MAO, and COMT are used for the treatment of Parkinson's disease (Bonifacio et al, 2007; Di Stefano et al, 2009; Fernandez and Chen, 2007; Gordin et al, 2004). Monitoring of dopaminergic metabolism during drug treatment is therefore useful to assess therapy. We evaluated the effects of AADC, MAO, and COMT inhibitors. Peripheral L-DOPA is immediately decarboxylated into DA, which cannot cross the blood–brain barrier (unlike L-DOPA). Peripheral AADC inhibitors such as carbidopa and benserazide are therefore co-administered with L-DOPA (levodopa) in the treatment of Parkinson's disease to increase the brain uptake of L-DOPA (Di Stefano et al, 2009). Benserazide increased extracellular L-DOPA (Figure 4B) as well as L-[β-11C]DOPA (Figure 3B). Although benserazide is a strong peripheral AADC inhibitor, it may also decrease central AADC activity and is detected in cerebral ECF (Jonkers et al, 2001; Silva et al, 1997). We also detected a benserazide peak in the striatum dialysate, which suggests that central AADC activity may be affected, and explains why L-[β-11C]DOPA levels increased and the delayed accumulation of [11C]DOPAC and [11C]HVA. In the presence of a peripheral AADC inhibitor, COMT (which is widely distributed) becomes the major metabolizing enzyme for L-DOPA, thereby increasing 3-OMD levels. Although extracellular endogenous 3-OMD was too low to detect (<1 nmol/L in our analytical conditions) (Napolitano et al, 2003), [11C]3-OMD could be detected. Plasma [11C]3-OMD gradually increased up to 20 minutes after intravenous L-[β-11C]DOPA until total 11C-radioactivity was derived from [11C]3-OMD and L-[β-11C]DOPA in a 1:1 ratio (Figure 5). Radioactive 3-OMD can be detected in plasma (Leger et al, 1998; Tsukada et al, 1994) and increases gradually (as observed here). The [11C]3-OMD levels in striatal ECF are low after intrastriatal administration of L-[β-11C]DOPA (data not shown), suggesting that [11C]3-OMD in striatal dialysate was generated mostly in peripheral tissue. The 3-OMD can readily pass through the blood–brain barrier through a large neutral amino acid system (Cumming and Gjedde, 1998). The [11C]3-OMD and L-[β-11C]DOPA are transported through the same large neutral amino acid system and 3-OMD has a longer half-life than L-DOPA, so the influx of L-[β-11C]DOPA is reduced by the presence of plasma [11C]3-OMD (Cumming and Gjedde, 1998; Ishikawa et al, 1996; Leger et al, 1998). In addition, a high concentration of 3-OMD in plasma and cerebrospinal fluid is detected in Parkinson's disease with chronic levodopa therapy, which causes dyskinesia (Benetello et al, 1997; Hardie et al, 1986; Tohgi et al, 1991) and oxidative stress (Lee et al, 2008). Therefore, reducing accumulation of 3-OMD with COMT inhibitors is effective in the treatment with levodopa/ AADC inhibitor, and also in PET studies (Ishikawa et al, 1996). Treatment with the MAO inhibitor, pargyline, increased levels of endogenous 3-MT and DA (Figure 4C), with most of the 11C-radioactivity being derived from [11C]3-MT (Figure 3C). In this case, [11C]DOPAC decreased significantly, thereby reducing [11C]HVA, changes that were also observed for endogenous compounds. The decrease in DOPAC with the accumulation of DA could elevate 3-MT levels in ECF. Although pargyline increased endogenous DA levels, [11C]DA was still not detectable. Extracellular [11C]DA could be immediately metabolized by COMT, resulting in increased [11C]3-MT, and limited to under and/or near the detection limit of [11C]DA (<1Bq).
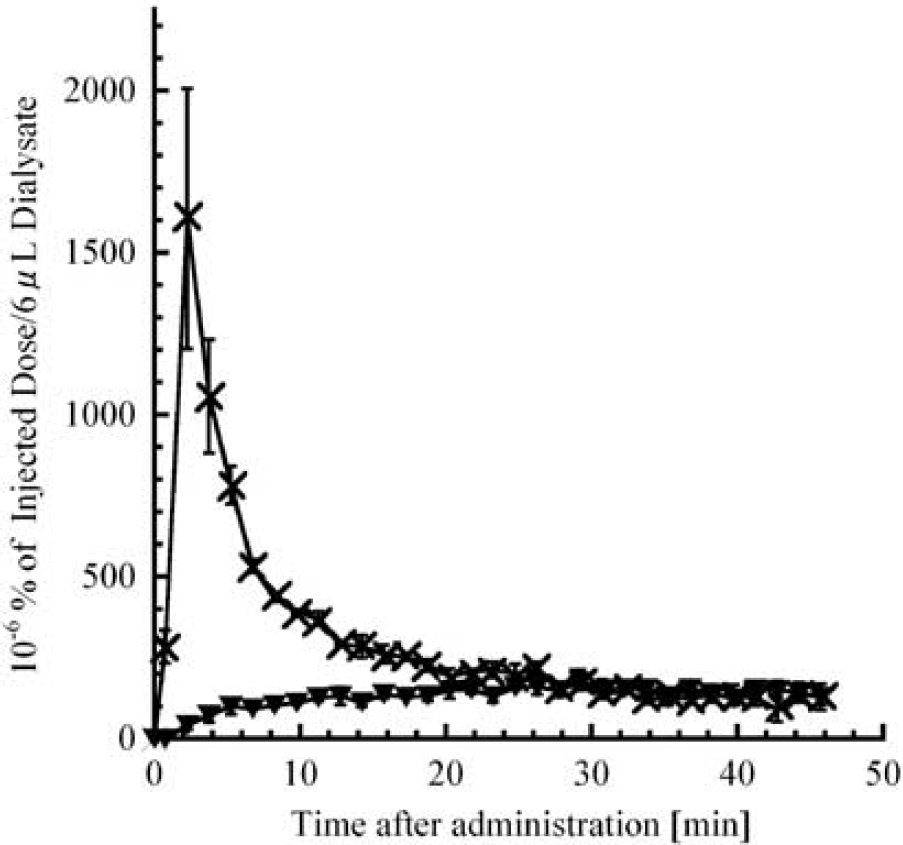
Time–radioactivity curves of L-DOPA and its metabolites obtained from femoral arterial dialysate. Rats were treated with benserazide 30 minutes before L-[β-11C]DOPA injection. The horizontal axis indicates the time after administration of L-[β-11C]DOPA (minutes). x, L-[β-11C]DOPA; ▾, [11C]3-OMD. Values are mean ± s.e.m. from three rats. L-DOPA; 3,4-dihydroxy-L-phenylalanine; 3-OMD, L-3-O-methyl-DOPA.
The COMT inhibitor, 3,5-dinitrocatechol, increased [11C]DOPAC and eliminated [11C]3-OMD (Figure 3D) but did not change L-[β-11C]DOPA, because it is preferentially metabolized by AADC, indicating that COMT inhibitors are effective only in combination with levodopa/AADC inhibitor. Nonradioactive DOPAC levels did not increase (Figure 4D), suggesting that radiometric detection is more sensitive. We also detected an unknown 11C-labeled metabolite eluted at 0.9 minutes (‘[11C]unknown’ in Figure 2A).
We showed that the kinetics of L-[β-11C]DOPA and its metabolites from rat striatal dialysate reflect the complicated metabolic processes of L-DOPA. Furthermore, although extracellular [11C]DA could not be detected due to its extremely low radioactivity, this method may be a useful tool for validating the quantitative analysis of L-[β-11C]DOPA in a PET study. Our method could improve the understanding of the information obtained from an L-[β-11C]DOPA PET study or provide an index of dopaminergic metabolism-related neuropsychiatric disorders and evaluation of drug therapies for the treatment of the disorder.
Footnotes
Acknowledgements
The authors are grateful to Mr M Takei and Mr N Nengaki for their technical support in the synthesis of L-[β-11C]DOPA.
The authors declare no conflict of interest.