We investigated the systemic effect of liposomes bearing apoptotic signals on the level of inflammation and neuronal death induced by ischemia–reperfusion (IR). Using a model of retinal ischemia, we showed that treatment with phosphatidylserine (PS) and phosphatidylcholine (PC) liposomes significantly reduced the expression of proinflammatory genes, including that of Il1b, Il6, Ccl2, Ccl5, Cxcl10, and Icam1, 24 h after reperfusion. Phosphatidylserine liposome treatment was the most efficient and correlated with significantly reduced neuronal death in the retina 7 days after reperfusion. The results of our study indicate that therapeutic strategy based on mimicking a systemic increase in apoptotic signaling can significantly reduce central nervous system damage induced by IR and improve neurologic outcome.
Stroke is an acute neurologic injury, the third leading cause of death in the United States (after heart disease and cancer), and is responsible for 10% of deaths worldwide (Feigin, 2005). Ischemic stroke comprises 80% of all stroke cases (Feigin, 2005; Lipton, 1999). Energy depletion leads to necrotic cell death in the areas most severely affected by ischemia. Apoptotic neurons are concentrated in areas of milder insult (Lipton, 1999). Professional phagocytes discriminate between apoptotic and necrotic cells and respond accordingly. It has been shown that apoptotic cell death has an important role in the resolution phase of inflammation. Phagocytes that have internalized apoptotic cells inhibit the production of proinflammatory mediators and stimulate the production of antiinflammatory factors (Fadok et al, 2001; Huynh et al, 2002; Minghetti et al, 2005). Thus, the interplay between apoptotic cells and phagocytes seems to create a microenvironment that can suppress immune and inflammatory responses.
The observation of the antiinflammatory effect of apoptotic cells raises the question, by which mechanisms might this antiinflammatory effect be mediated. Phosphatidylserine (PS) is an aminophospholipid that is predominantly found in the inner leaflet of the plasma membrane of virtually every body cell. During apoptosis, PS is exposed on the outer leaflet of the cytoplasmic membrane and serves as a target for the PS receptor of professional phagocytes, including macrophages and microglia (Henson et al, 2001; Huynh et al, 2002). The latter could be responsible for antiinflammatory signaling (Hoffmann et al, 2005; Huynh et al, 2002). Presently, four different PS receptors have been identified and characterized (Bratton and Henson, 2008; Henson et al, 2001). Furthermore, PS-containing liposomes have been shown in vitro and in vivo to mimic the effects of apoptotic cells on phagocytes (Fadok et al, 2001; Hashioka et al, 2007; Minghetti et al, 2005; Nolan et al, 2004). We hypothesized that an increase in the systemic level of apoptotic signals facilitated by an intraperitoneal injection of the PS liposomes can inhibit the inflammation of the central nervous system and improve the survival rate of neurons after ischemia–reperfusion (IR) injury. We investigated the effect of the PS liposome injection on a number of surviving retinal neurons challenged with retinal IR.
Materials and methods
Animals
All experiments and postsurgical care were carried out in compliance with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals and according to the Institutional Animal Care and Use Committee (IACUC)-approved protocols. All animals that were used in our experiments were 3-month-old C57BL/6 male mice (seven animals per group).
Transient Retinal Ischemia
After anesthesia with intraperitoneal ketamine (80 mg/kg) and xylazine (16 mg/kg), the pupils were dilated with 1% tropicamide–2.5% phenylephrine hydrochloride (NutraMax Products, Gloucester, MA, USA), and corneal analgesia was achieved with 1 drop of 0.5% proparacaine HCl (Bausch & Lomb Pharmaceuticals, Tampa, FL, USA). Retinal ischemia was induced for 60 mins by introducing into the anterior chamber of the eye a 33-G needle attached to a normal (0.9% NaCl) saline-filled reservoir that was raised above the animal to increase intraocular pressure above systolic blood pressure (intraocular pressure increased to 120 mm Hg). The contralateral eye was cannulated and maintained at normal intraocular pressure to serve as a normotensive control. Complete retinal ischemia, evidenced by a whitening of the anterior segment of the eye and by blanching of the retinal arteries, was verified by a microscopic examination. After needle removal, 1% atropine and 1% vetropolycin with hydrocortisone ointment (Fougera & Atlanta, Melville, NY, USA) were applied to the conjunctival sac. Mice were killed by CO2 inhalation under anesthesia.
Phosphatidylserine and Phosphatidylcholine Liposome Treatment
Liposome preparation is described in detail in Supplement 1. In brief, the liposomes were composed of a combination of l-[α]-phosphatidylserine (PS) and l-[α]-phosphatidylcholine (PC) at a molar ratio of 3:7 (PS:PC) (PS liposomes). For PC liposomes preparation, only l-[α]-phosphatidylcholine lipid were used. Mice were injected intraperitoneally with either liposomes (0.5 mg suspension in phosphate-buffered saline (PBS) per animal) or carrier buffer (PBS) before IR injury, at the time of surgery, and thereafter every 24 h.
Counting of NeuN-Positive Ganglion Cell Layer Neurons
Neuronal-nuclei (NeuN)-positive neurons in the ganglion cell layer (GCL), including retinal ganglion cells and displaced amacrine cells, were imaged by confocal microscopy in flat-mounted retinas. Individual retinas were sampled randomly to collect a total of 20 images located at the same eccentricity in the four retinal quadrants using × 20 objective lens. The NeuN-positive neurons were counted semi-automatically using the MetaMorph (Universal Imaging, Downingtown, PA, USA) software. Cell loss in ischemic retinas was calculated as the percentile of the mean cell density in fellow control eyes.
Statistical Analysis
Statistical analysis of real-time PCR and cell density data was carried out with one-way ANOVA (analysis of variance) followed by Tukey's test for multiple comparisons. In the case of single comparisons, Student's t-test was applied. P-values ≤0.05 were considered statistically significant.
For experiments using real-time PCR and immunohistochemistry, we applied standard techniques; brief protocols are explained in Supplement 1.
Results
Presence of PS Liposomes Resulted in Reduced Inflammation After Retinal Ischemia
We have studied the influence of PS liposomes on the inflammatory level of tissues in vivo in a retinal IR model. It has been shown that liposomes are efficiently incorporated into the brain across the blood–brain barrier. In particular, Chapat et al (1991) have used carboxyfluorescein-loaded liposomes to show that those were able to cross the blood–brain barrier of the postischemic tissue. We injected experimental mice intraperitoneally with PS liposomes twice: 24 h before IR and at the time of surgery. Control animals were injected with PC liposomes at an equimolar concentration or PBS. We induced unilateral retinal ischemia in mice by increasing the intraocular pressure above normal systolic levels.
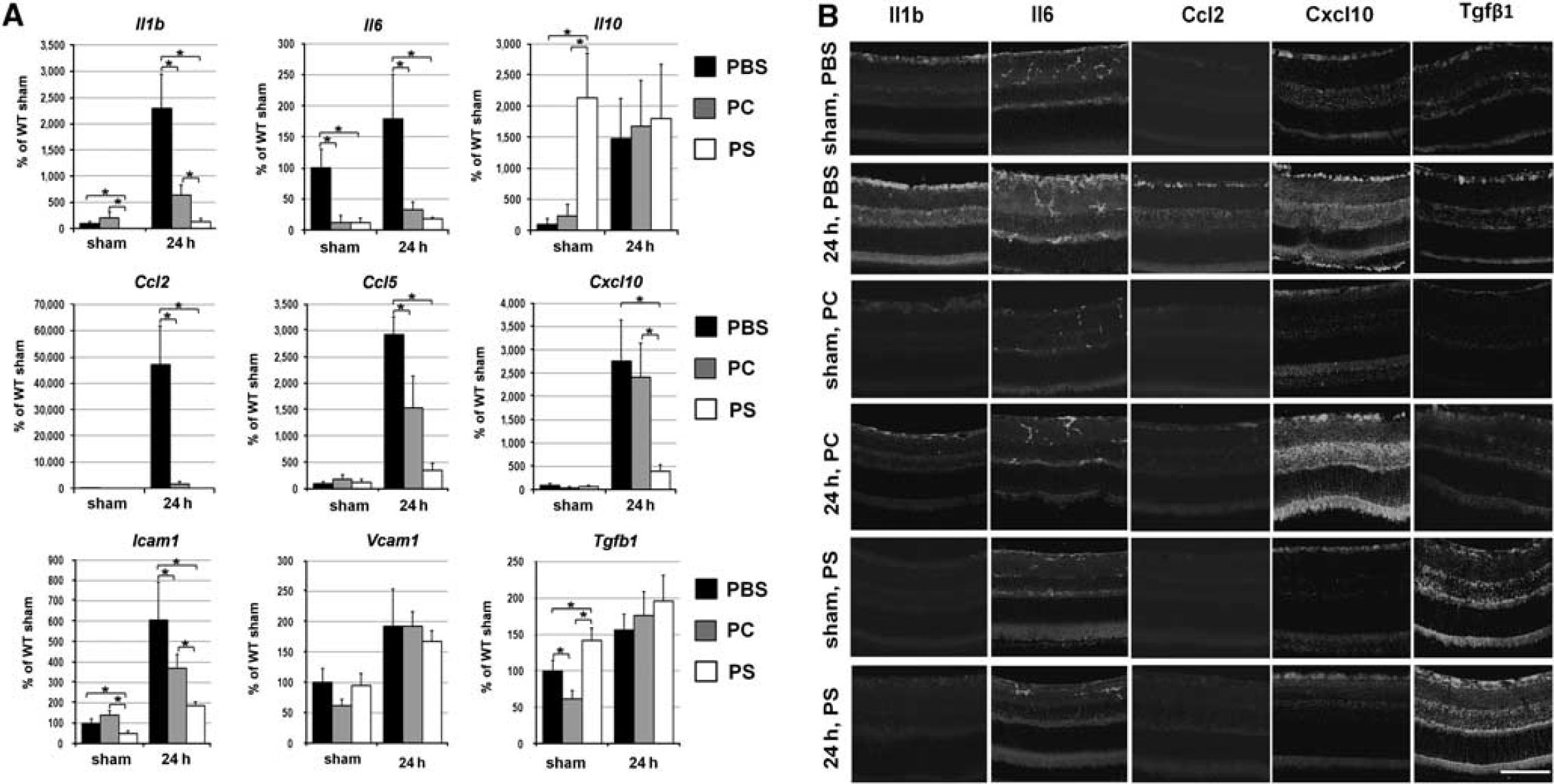
We assessed gene expression 24 h after reperfusion using real time-PCR, because most changes in gene expression for proinflammatory factors typically occur shortly after ischemic injury. The transcriptional upregulation of cytokines (Il1β and Il6), chemokines (Ccl2, Ccl5, and Cxcl10), and cell adhesion molecules (Icam1) was evident in experimental eyes of animals that were treated with PS, PC liposomes, or PBS (Figure 1A). However, in the mice treated with PS liposomes, the expression of these genes was significantly reduced relative to PBS-treated animals (Figure 1A). Significantly, in the mice treated with PS liposomes, the levels of three genes, namely Il1β, Cxcl10, and Icam1, were significantly (P<0.05) reduced relative to PC-liposome-treated animals (4.8-, 6.1-, and 2.0-fold change, respectively, Figure 1A). The gene expression profiles for Il1β, Il6, Ccl2, Cxcl10, Tgfβ1, and Icam1 were consistent with the corresponding protein accumulation levels detected by immunohistochemistry 24 h after reperfusion (Figure 1B and Supplement 2). The gene expression changes for Tgfβ1 and Il10 genes, after retinal ischemia were not statistically different between animals treated with PS liposomes and control (PC-liposome- and PBS-treated) animals. Intriguingly, the PS-liposome-mediated increase in apoptotic signals in the contralateral control (sham-operated) eye resulted in a statistically significant reduction of inflammation. Thus, the transcriptional activation of the antiinflammatory molecules (Tgfβ1 and Il10), as well as the suppression of the cytokines (Il1b and Il6) and the cell adhesion molecule (Icam1) were evident in PS-liposome-treated, but not in PC-liposome-treated sham-operated eyes (Supplement 3).
Treatment by PS-liposomes suppresses induction of pro-inflammatory markers after IR: (A) Differential expression of the cytokines, the chemokines, and the cell adhesion molecules in the retinas of PS, PC liposomes, and PBS-treated animals after IR. The gene expression was assessed using real-time PCR in sham-operated controls and experimental retinas after IR. For each gene, results are expressed as a percentage of the corresponding value in sham-operated eye of PBS-treated animals±s.e.m. after normalization to β-actin. Statistical analysis was carried out with one-way ANOVA followed by Tukey's test (∗P<0.05, n=7). (B) Immunohistochemistry for the Il1b, Il6, and Ccl2 proteins accumulation in the postischemic retinas of PS, PC liposomes, and PBS-treated mice are consistent with increased levels of the transcripts at the level of corresponding proteins. Scale bar: 100 μm.
Presence of PS Liposomes Reduced the Severity of Ischemic Damage to the Retina
Retinal ischemia causes GCL neuron loss, which, in similarity to ischemic stroke, is biphasic. Primary degeneration occurs within 24 h after reperfusion, whereas secondary degeneration progresses over several days (Crotty et al, 2008; Hoffmann et al, 2005; Minghetti et al, 2005; Nolan et al, 2004). To detect the cumulative survival effect after both phases of degeneration, we assessed neuronal density in the GCL 1 week after reperfusion. The mice were injected intraperitoneally with liposomes or PBS 24 h before IR, at the time of surgery, and thereafter every 24 h. We evaluated neuronal cell death by counting the number of neurons in the GCL in flat-mounted retinas. Neurons were labeled with the neuronal marker, NeuN, and the quantity of cells was determined.
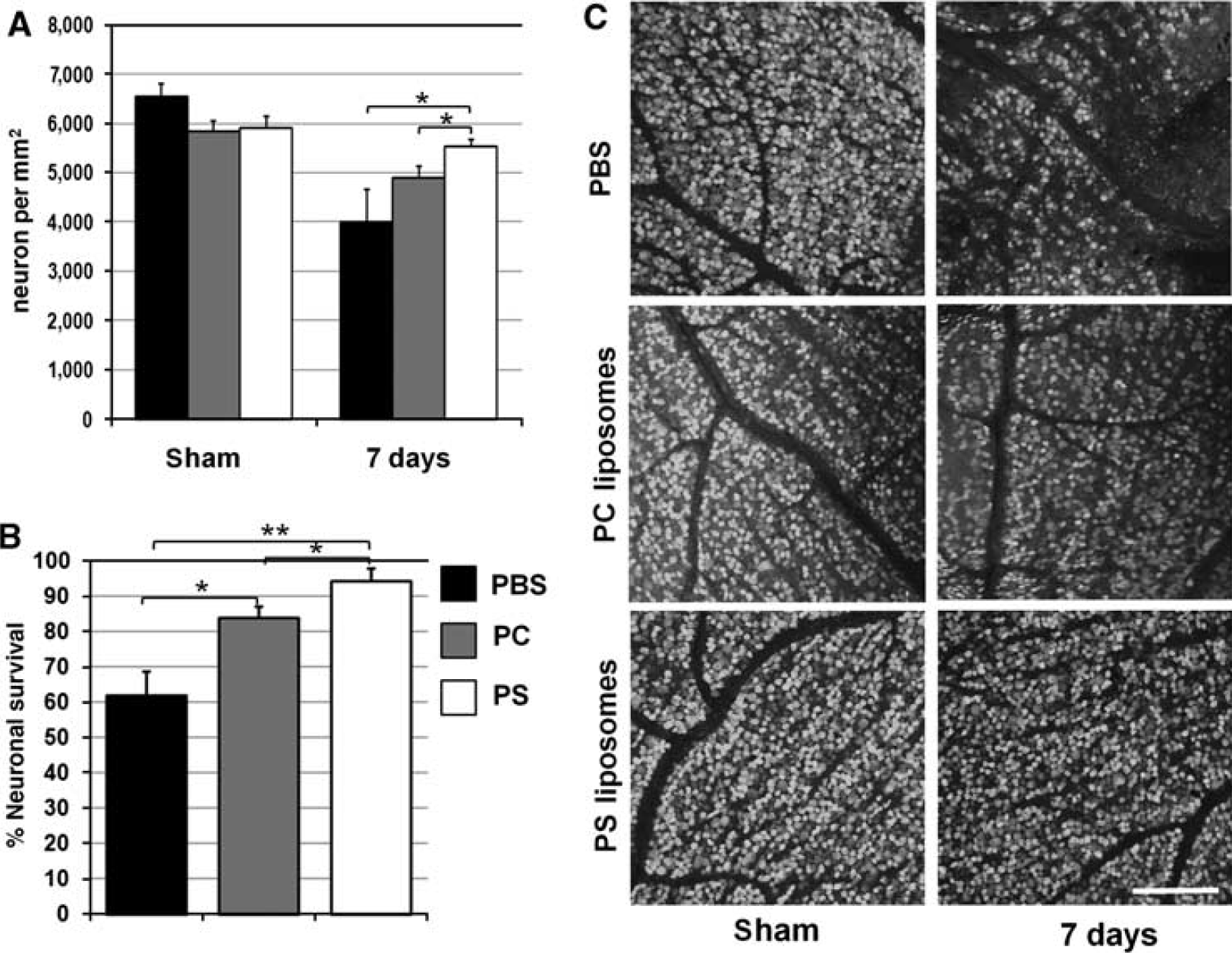
There was no statistically significant difference in the density of neurons in the sham-operated eyes treated by PS, PC liposomes, and PBS (Figure 2A). In contrast, the IR-induced loss of retinal neurons was apparent in experimental eyes of both liposome- and PBS-treated retinas 7 days after reperfusion (Figure 2). A comparison of neuronal loss among PS, PC liposomes, and PBS-treated mice, has shown significant differences in neuronal resistance to the IR injury. The mean density of GCL neurons in the ischemic retina was significantly higher in mice injected with PS liposomes compared with those injected with PC liposomes (P<0.05) and PBS (P<0.05) (Figure 2A). Retinas in PBS-treated mice had significantly lower number of surviving NeuN-positive neurons as a percentage of the sham-operated eye (62±6%) in GCL as compared with retinas from animals treated with either PC liposomes (84±3%, P <0.01) or PS liposomes (94±4%, P<0.001) (Figure 2B).
Treatment by PS liposomes results in neuroprotective effects in the GCL of retinas after IR: (A) mean densities (neuron per mm2) and (B) percentage of surviving of GCL neurons in the IR retinas of PS, PC liposomes, and PBS-treated animals 7 days after IR. (C) Representative confocal images of NeuN-labeled GCLs (green) in flat-mounted retinas acquired at 1 mm from the optic nerve in sham-operated controls and ischemic retinas 7 days after reperfusion. Statistical analysis was carried out with one-way ANOVA followed by Tukey's test (∗P<0.05, ∗∗P<0.01, n=7). Scale bar: 100 μm.
Discussion
It has been shown previously in vitro that phagocytes in the presence of PS liposomes decrease the synthesis of proinflammatory factors, such as tumor necrosis factor-α, interleukin-1β, interleukin-6, CCL2, and nitric oxide and increase the production of antiinflammatory factors, such as TGF-β1 (Crottyet al, 2008; Hoffmann et al, 2005; Minghetti et al, 2005; Nolan et al, 2004). Our data indicate that the inflammatory response observed in the retina 24 h after reperfusion was suppressed in animals treated by PS liposomes. It is noted that injection of PC liposomes also reduced the level of inflammatory gene expression in the retina, but to a lesser extent compared with PS liposomes. PC liposomes have been shown to reduce the production of proinflammatory and toxic molecules in phagocytes (Hashioka et al, 2007). However, the expression of the proinflammatory genes, namely Il1β, Cxcl10, and Icam1, was significantly reduced in the mice treated with PS liposomes compared with PC-liposome-treated animals, which correlated with significantly increased neuroprotection observed after PS liposome treatment. Given that these genes have an important role in ischemic pathology (Fadok et al, 2001; Hashioka et al, 2007; Nolan et al, 2004), the increase in protection could be attributed to the reduced activity of these genes. Significantly, in agreement with a previous report (Huynh et al, 2002), our data showed that PS liposome injection facilitated systemic antiinflammatory changes, which PC liposomes did not; the expression of antiinflammatory genes such as Tgfβ1 and Il10 was upregulated, whereas the expression of proinflammatory genes was reduced in the control (sham-operated) retinas. The lack of a similar effect in control eyes after PC liposome injection allows us to speculate that the systemic antiinflammatory effect is caused by an increase in apoptotic signals facilitated by PS liposomes.
The effect of liposome treatment was assessed by a comparison of neuronal survival rates in the retinal GCL in liposome-treated and control animals. Injection of PS liposomes versus PBS significantly increased neuronal survival after retinal ischemia. Although less efficient than PS liposome injection, the neuroprotective effect of the PC liposome treatment was also shown (P<0.05). The antiinflammatory effect of PC liposomes is in agreement with the results obtained in vitro in an earlier study (Hashioka et al, 2007). The protective role of PS liposomes has been shown previously in vivo only in the rat hippocampus lipopolysaccharide (LPS) injection model (Nolan et al, 2004). The data obtained from this study indicate that this protection is distinct from the antiinflammatory effect triggered by PC liposomes or native cell membranes (also PC-containing), that provided phagocytes with an antiinflammatory stimulus (Fadok et al, 2001; Hashioka et al, 2007). To our knowledge, this study represents the first report on protection from the IR-induced pathology in the retina induced by PS liposomes. The results of our study suggest that increased apoptotic signals introduced by PS-containing liposomes can promote a neuroprotective environment, reduce tissue damage, and may be considered as a potential therapy for IR injury.
Footnotes
The authors declare no conflict of interest.
Notes
Supplementary Information accompanies the paper on the Journal of Cerebral Blood Flow & Metabolism website ()
References
1.
BrattonDLHensonPM (2008) Apoptotic cell recognition: will the real phosphatidylserine receptor(s) please stand up?Curr Biol18:R76–9
2.
ChapatSFreyVClaperonNBouchaudCPuisieuxFCouvreurPRossignolPDelattreJ (1991) Efficiency of liposomal ATP in cerebral ischemia: bioavailability features. Brain Res Bull26:339–42
3.
CrottySFitzgeraldPTuohyEHarrisDMFisherAMandelABoltonAESullivanAMNolanY (2008) Neuroprotective effects of novel phosphatidylglycerolbased phospholipids in the 6-hydroxydopamine model of Parkinson's disease. Eur J Neurosci27:294–300
4.
FadokVABrattonDLGuthrieLHensonPM (2001) Differential effects of apoptotic versus lysed cells on macrophage production of cytokines: role of proteases. J Immunol166:6847–54
5.
FeiginVL (2005) Stroke epidemiology in the developing world. Lancet365:2160–1
6.
HashiokaSHanYHFujiiSKatoTMonjiAUtsumiHSawadaMNakanishiHKanbaS (2007) Phosphatidylserine and phosphatidylcholine-containing liposomes inhibit amyloid beta and interferon-gamma-induced microglial activation. Free Radic Biol Med42:945–54
7.
HensonPMBrattonDLFadokVA (2001) The phosphatidylserine receptor: a crucial molecular switch?Nat Rev Mol Cell Biol2:627–33
8.
HoffmannPRKenchJAVondracekAKrukEDalekeDLJordanMMarrackPHensonPMFadokVA (2005) Interaction between phosphatidylserine and the phosphatidylserine receptor inhibits immune responses in vivo. J Immunol174:1393–404
9.
HuynhMLFadokVAHensonPM (2002) Phosphatidylserine-dependent ingestion of apoptotic cells promotes TGF-beta1 secretion and the resolution of inflammation. J Clin Invest109:41–50
10.
LiptonP (1999) Ischemic cell death in brain neurons. Physiol Rev79:1431–568
11.
MinghettiLAjmone-CatMADe BerardinisMADe SimoneR (2005) Microglial activation in chronic neurodegenerative diseases: roles of apoptotic neurons and chronic stimulation. Brain Res Brain Res Rev48:251–6
12.
NolanYMartinDCampbellVALynchMA (2004) Evidence of a protective effect of phosphatidylserine-containing liposomes on lipopolysaccharide-induced impairment of long-term potentiation in the rat hippocampus. J Neuroimmunol151:12–23
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.