
Abstract
Heat shock protein 27 (HSP27) has a major role in mediating survival responses to a range of central nervous system insults, functioning as a protein chaperone, an antioxidant, and through inhibition of cell death pathways. We have used transgenic mice overexpressing HSP27 (HSP27tg) to examine the role of HSP27 in cerebral ischemia, using model of permanent middle cerebral artery occlusion (MCAO). Infarct size was evaluated using multislice
Introduction
Heat shock proteins (HSPs) form a heterogeneous family of proteins that exhibit a diverse range of protein chaperoning properties facilitating protein folding, trafficking, and removal of aberrant proteins. In addition, certain HSPs are induced during cellular stress and contribute to cell survival through antioxidant and antiapoptotic properties. These properties have been particularly well established in cell culture and using
The well-documented antiapoptotic properties of HSP27 exhibited in cell culture (Garrido et al, 1999; Bruey et al, 2000; Paul et al, 2002) have also been shown
In this study, we have used transgenic mice overexpressing HSP27 (HSP27tg) to examine the role of HSP27 in a permanent MCAO (pMCAO) model of cerebral ischemia, compared with the transient ischemia model used by Stetler et al (2008). We assessed the severity of injury using magnetic resonance imaging (MRI) to determine the overall lesion volume, estimated after 24 h of ischemia. Our results provide support for the powerful neuroprotective effects of HSP27 in cerebral ischemia in this model and further provide evidence that the presence of HSP27 enhances cell survival in regions that undergo cell death in wild-type (WT) littermates. This difference occurs despite marked induction of c-jun and ATF3 early after MCAO and shows a profound inhibitory effect of HSP27 on the c-junmediated cell death cascade.
Materials and methods
Animals
All animal care and procedures were in accordance with the UK Animals (Scientific Procedures) Act 1986. Transgenic mice overexpressing human HSP27 (HSP27o/e) (under the control of a chicken β-actin promoter and a cytomegalovirus enhancer element) on a C57BL10/CBA background were used. The line was maintained by continuously back-crossing with C57BL10/CBA mice (Akbar et al, 2003).
Middle Cerebral Artery Occlusion Model
Adult male (weighing 24 to 28 g; 10 to 13 weeks old) HSP27tg mice (
Magnetic Resonance Imaging
Coronal images were obtained with the central slice ∼0.5 mm from bregma at 2 and 24 h after the onset of stroke. Magnetic resonance imaging measurements were performed on a 2.35-T horizontal bore magnet (Oxford Instruments, Oxford, UK) interfaced to a SMIS console (Surrey Medical Imaging Systems, Guildford, UK). The radio frequency pulses were transmitted with a volume coil of 6.5 cm length. The signal was received with a separate 1-cm-diameter surface coil.
After MCAO, the animals were transferred to the magnet and imaged using the following protocol:
Multislice
Image Processing
Perfusion maps were obtained by subtracting the labeled image from the control image. The
The lesion area was determined manually by an observer blinded to the experiment, using the multislice anatomical
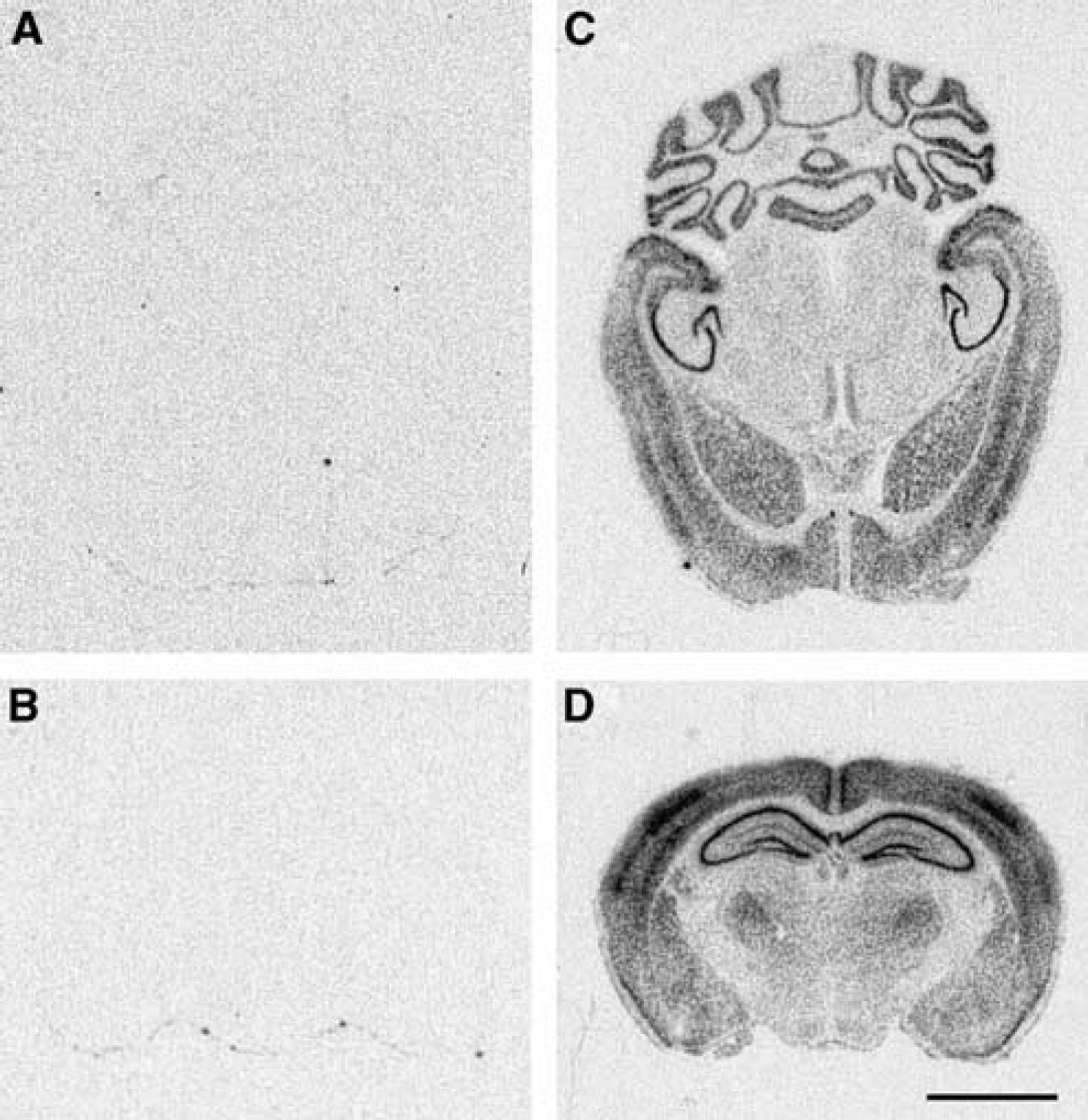
Representative
Cerebral blood flow images were analyzed on a region-of-interest basis. Ten regions of interest were placed in the medial cortex, the dorsolateral cortex, the ventrolateral cortex, the lateral caudate putamen, and the medial caudate putamen of each hemisphere. The CBF of the affected hemisphere was expressed relative to the contralateral side.
In Situ Hybridization
For
Specific antisense oligonucleotides were synthesized (Sigma-Genosys, Cambridge, UK) for use in the
Statistics
The lesion volume per slice was compared using a degree of freedom adjusted repeated-measures analysis of variance (ANOVA). For each analysis, the appropriate within-subject variables and between-subject factors were adopted. Differences were considered significant when the
Results
Lesion volume
The HSP27 transgenic mouse line used in this study has been characterized extensively in earlier studies (Akbar et al, 2003) and shows no effect of the genetic modification on development or survival. The transgene is widely expressed across multiple tissues and is particularly abundant in the cerebral cortex, the hippocampus, and the striatum (Figure 1). To determine whether HSP27 overexpression confers protection against ischemic injury, HSP27tg (
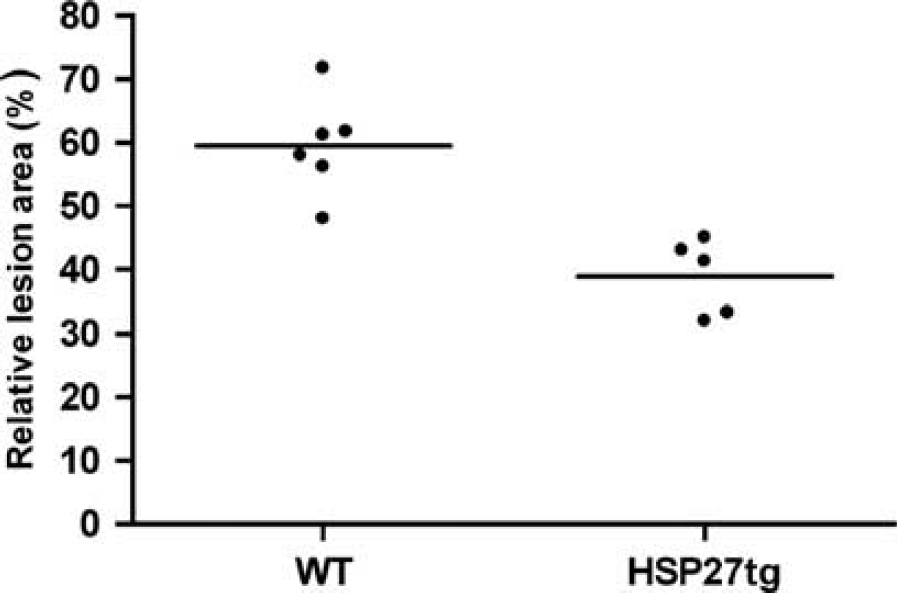
Effect of heat shock protein 27 (HSP27) overexpression on the lesion area after permanent middle cerebral artery occlusion. Relative lesion area for transgenic mice overexpressing HSP27 (HSP27tg) and control mice. The lesion area was calculated relative to the contralateral hemispherical area (horizontal bar indicates the group mean). Mixed model analysis of variance (ANOVA) showed a significant difference in lesion area between the HSP27tg mice and the WT mice.
Successful occlusion of the MCA results in a severe reduction of CBF in a large part of the ipsilateral hemisphere. However, there were no significant differences in CBF reduction within the lesion area between WT and HSP27tg at 2 or 24 h after stroke (Supplementary Figure 2) as previously found with HSP70tg mice (van der Weerd et al, 2005).
Glial Fibrillary Acidic Protein expression
The spatial and temporal expression profile of three sensitive markers of tissue injury, GFAP, c-jun, ATF3, and two endogenous HSPs, mHSP25 and HSP70, in WT and HSP27tg mice after pMCAO was assessed using
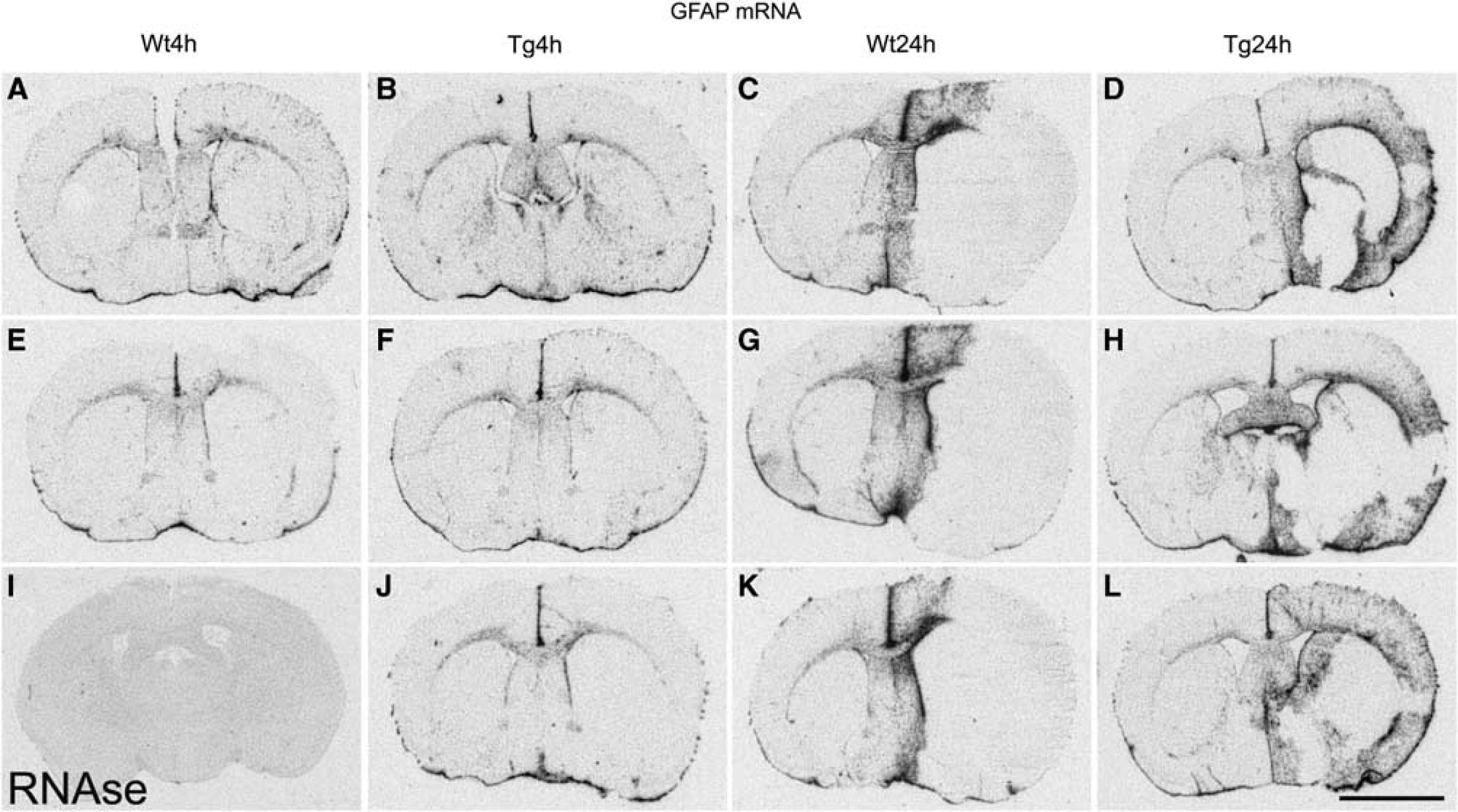
The effect of middle cerebral artery occlusion (MCAO) on glial fibrillary acidic protein (GFAP) expression. c-jun mRNA detected by
c-jun Expression
In sham-operated control mice, there were low-to-moderate levels of constitutive expression of c-jun mRNA throughout the cortex. In contrast, at 4 h after MCAO, a profound induction of c-jun was observed in most animals in both WT and HSP27tg mice (Figures 4A, 4B, and 4F). c-jun mRNA expression in these animals was evident throughout the ischemic hemisphere, the outer and deep cortical layers showing the highest levels compared with the intermediate layers. Variable expression was seen in the caudate and putamen in the ischemic hemisphere. However, the most intense level of c-jun labeling was seen in the cortical and striatal regions of HSP27tg mice. These regions coincided with the areas of tissue preservation in HSP27tg mice at 24 h (Figures 4B and 4F). This contrasts with the extensive infarcts that develop in the same regions in control animals. Intense labeling was also seen in the pyriform cortex of all mice in the ipsilateral cortex at this time point with moderate expression in the pyriform cortex of the contralateral side (Figures 4A, 4B, 4E, 4F, and 4J). There was a low level expression of c-jun mRNA in the contralateral cortex in both WT and HSP27tg mice.
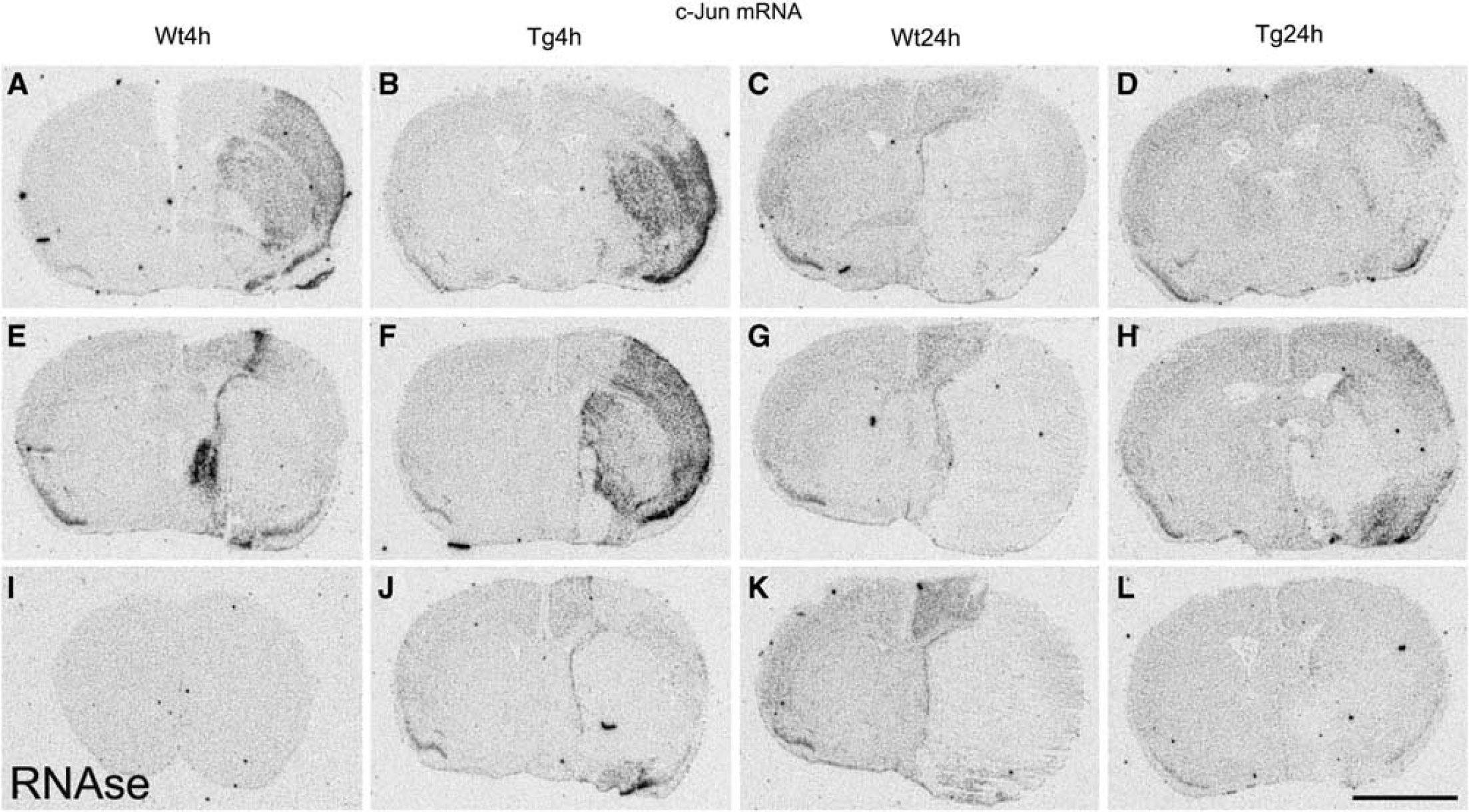
The effect of middle cerebral artery occlusion (MCAO) on c-jun expression. c-jun mRNA detected by
At 24 h, the expression levels of c-jun mRNA were generally reduced. In WT mice, there was moderate expression throughout the ventral, lateral, and dorsal cortex in the contralateral hemisphere, whereas strong labeling in the pyriform cortex was maintained. In the ipsilateral hemisphere, c-jun mRNA was confined to the penumbral tissue with no signal detected in the infarct area. In HSP27tg mice at 24 h after MCAO, expression of c-jun mRNA in the contralateral hemisphere was similar to that described for WT mice. However, expression in the ipsilateral hemisphere of HSP27tg mice showed a rather different pattern with intense to moderate labeling seen in the larger area of surviving cortical and striatal tissue (penumbral tissue/peri-infarct area) and more intense labeling in the pyriform cortex compared with WT mice (Figures 4D, 4H, and 4L).
ATF3 expression
The expression of ATF3 mRNA in forebrain sections from control nonoperated/naive or sham-operated animals showed no significant hybridization signal (data not shown). A similar pattern of ATF3 mRNA expression was seen throughout the nonischemic (contralateral) hemisphere of both WT and HSP27tg mice killed 4 h after pMCAO (Figures 5B and 5E). In contrast, a marked induction of ATF3 mRNAwas seen in the caudate/putamen in the ischemic (ipsilateral) hemisphere in sections from WT and HSP27tg mice (Figures 5B and 5E). Moderate labeling was seen in the overlying cortex, predominantly in the lateral and ventral cortex. These regions define the core of the infarct area. A strong signal was also apparent in the pyriform cortex. It is noteworthy that the animals with the highest levels of c-jun expression were also accompanied by the highest levels of ATF3 and HSP70 expression (Figures 5A, 5B, and 5C and, respectively). At 24 h after pMCAO, ATF3 mRNA expression levels had declined to background levels in both WT and HSP27tg mice (data not shown).
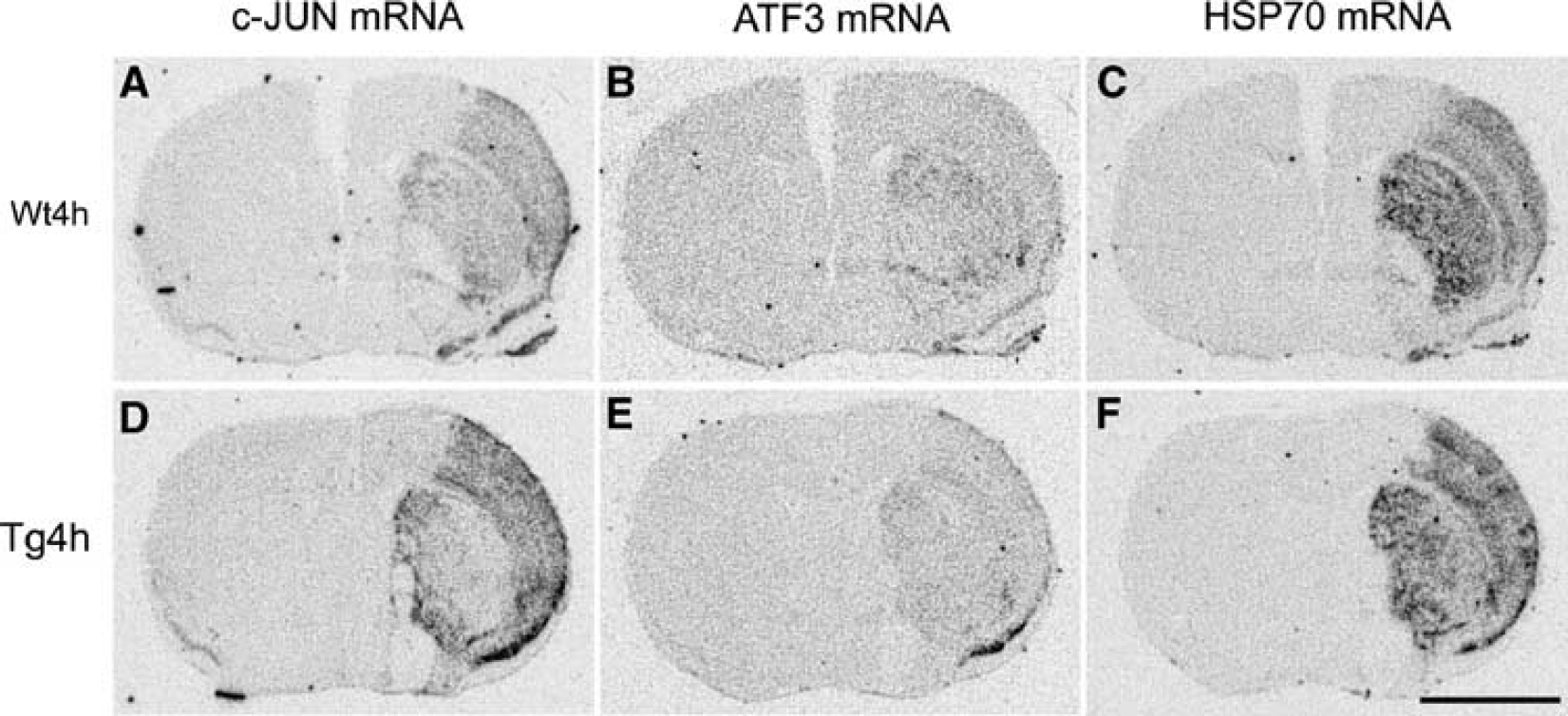
The effect of middle cerebral artery occlusion (MCAO) on expression ATF3 compared with c-jun and HSP70 mRNA.
Expression of HSP70 and HSP25
There was no signal detected for HSP70 mRNA in sections from sham-operated mice (data not shown). HSP70 mRNA expression generally showed a similar labeling pattern to that observed for c-jun. At 4 h after MCAO onset, there was robust expression of HSP70 mRNA in the ipsilateral hemisphere in both WT and HSP27tg mice (Supplementary Figures 3A, 3B, 3E, 3F, and 3J). Intense labeling was evident in the pyriform cortex, in the outer and inner layers of the cortex, with moderate signal detected in the intermediate cortical layers. Robust expression was also evident in the striatum. The distribution of HSP70 induction at 4 h showed considerable overlap with the expression of c-jun mRNA in near adjacent sections from the same animals (Figure 5).
At 24 h after pMCAO, the expression of HSP70 mRNA was much reduced. In WT animals, HSP70 mRNA was present in midline brain structures adjacent of the infarct region, although expression was granular/patchy in appearance (Supplementary Figures 3C, 3G, and 3K). In HSP27tg mice at this time point, strong expression of HSP70 mRNA was present in the outer and deep layers of the cortex in the ipsilateral hemisphere (Supplementary Figures 3D, 3H, and 3L). There was no hybridization signal detected in the contralateral hemisphere in both WT and HSP27tg mice at both time points after induction of focal cerebral ischemia.
There was moderate diffuse expression of endogenous inducible mHSP25 mRNA in the striatum and cortex of WT mice at 4 h after ischemia, which was reduced in HSP27tg mice (Supplementary Figure 4). At 24 h, there was a greater intensity of expression in midline peri-infarct areas and in the dorsal cortex in WT mice. No signal was detected in the infarct region. In HSP27tg mice at 24 h, there was robust expression of mHSP25 mRNA in the penumbral tissue with clear labeling in the deep and superficial layers of the cortex and low to moderate labeling in the intermediate cortical layers. There was no mHSP25 mRNA detected in the contralateral hemisphere. Overall, the expression of mHSP25 mRNA at 24 h in cells adjacent to infarcted tissue highlighted the extent of salvaged tissue in the HSP27tg animals.
Discussion
The MRI results described in this study clearly show that HSP27 over-expression reduces infarct size. This is also substantially supported by expression studies. The large infarct seen in WT animals affects the whole MCA territory in ipsilateral cortical and striatal regions. In contrast, in HSP27tg mice, although the striatum is severely affected, the majority of the cerebral cortex in the MCA is preserved at 24 h after pMCAO. Increased GFAP expression is seen in tissue adjacent to the infarct at 24 h in WT mice, being localized to regions largely outside the MCA territory. In HSP27tg mice, GFAP is present in rescued tissue ipsilateral to MCAO and little upregulation was detected outside the MCA territory.
To gain some insight into the mechanisms contributing to cell death and its attenuation by HSP27, we analyzed the expression of c-jun, ATF3, and HSP70. c-jun is upregulated within minutes of a cellular stress (Honkaniemi et al, 1997) and in the form of homodimers or heterdimers regulates transcription through the AP-1 binding site. Induction of c-jun contributes both to cell death through the JNK cascade and also survival responses to central nervous system insults mediated through the Akt pathway. In neutrophils, HSP27 also provides an essential role in regulating the antiapoptotic effects of Akt (Rane et al, 2003). Widespread expression of c-jun occurs after focal cerebral ischemia (Ohba et al, 2003, Honkaniemi et al, 1997). ATF3 is also induced after cerebral ischemia and nerve injury, and has been shown to colocalize in surviving neurons (Ohba et al, 2003, Takeda et al, 2000, Tsujino et al, 2000). ATF3 is known to protect vulnerable CA3 neurons from kainate toxicity (Francis et al, 2004), and furthermore, ATF3 has been shown to prevent JNK-induced neuronal death in PC12 cells by promoting HSP27 expression and Akt activation (Nakagomi et al, 2003). In the pMCAO model, the ischemic insult caused a rapid induction of c-jun and ATF3, which was mainly localized to the cortical and striatal regions served by the MCA at 4 h. At this stage, severe loss of cellularity was evident across both regions in WT animals, but was mainly restricted to striatal regions in HSP27tg mice.
Importantly, the c-jun induction seen at 4 h tended to be localized to regions that were salvageable. The 4-h time point coincides with high/maximal levels of JNK and c-jun phosphorylation occurring in MCA (Stetler et al, 2008). This is particularly marked in the HSP27 animals where cortical regions served by the MCA showed profound induction; at the later time point of 24 h, these regions showed greater cellularity. Interestingly, the distribution of HSP70 induction at 4 h showed considerable overlap overall and within animals with the expression of c-jun mRNA indicating a highly coordinated activation of the two pathways of transcriptional activation, mediated through heat shock factor-1 (HSF-1) in the case of HSPs and MAP kinases and their targets, such as cyclic AMP-responsive element binding protein (CREB), in the case of c-jun.
The induction of ATF3 was of much smaller magnitude than that of c-jun and was not detectable at 24 h. The distribution of c-jun mRNA was similar in WT and HSP27 tg mice but was absent in areas that had already undergone infarction, which was more extensive in WT animals. By 24 h, c-jun induction ipsilateral to MCAO was restricted to a midline region, which is not served by the MCA and coincides with the zone of GFAP induction. Interestingly, c-jun was induced throughout the contralateral hemisphere. In contrast, in HSP27 tg mice, c-jun was mainly present at low levels in the ipsilateral cortex. c-jun and other immediate early genes are known to show a widespread upregulation, which is attributed to spreading depression caused by glutamate release at the site of injury (Kiessling and Gass 1994).
The induction of HSP25 showed a marked difference between control and transgenic animals, being particularly evident at 24 h and mainly present in peri-infarct areas, which in control animals are seen outside the MCA territory in midline regions and in transgenic animals lie within the MCA territory associated with the much attenuated infarct. HSP27 is thought to be regulated by c-jun/ATF3 heterodimers, which have been shown to increase HSP25/27 transcription by binding to an atypical CRE motif in the promoter region of HSP27 (Nakagomi et al, 2003). This mechanism may be responsible for the induction of mHSP25 in striatal regions of WT animals at 4 h, at which time both c-jun and ATF3 are elevated. However, as ATF3 induction is transient, this regulatory mechanism is unlikely to explain the delayed more widespread increase in mHSP25 mRNA.
This paper shows that HSP27 mediates substantial neuroprotection in a model of pMCAO. In HSP27tg animals, infarct development is attenuated even in tissue producing abundant levels of c-jun, the substrate for JNK, which is critical for mediating neuronal cell death in cerebral ischemia (Gao et al, 2005). Furthermore, this effect is consistent with the ability of HSP27 to suppress JNK/MKK4 activity and subsequent cell death (Stetler et al, 2008). In summary, the neuroprotective properties of HSP27 shown in this and other studies have important therapeutic implications for the treatment of stroke.