
Abstract
We investigated intra-arterially administered autologous bone marrow mononuclear cells (MNCs) in rats with acute ischemic stroke. Long Evans rats (2 to 3 months or 12 months old) underwent tandem reversible common carotid artery (CCA)/middle cerebral artery (MCA) occlusion (CCAo/MCAo) for 3 h and then 24 h later underwent tibial bone marrow harvest. Ten million or 4 million cells were re-injected by an intra-carotid infusion. Control animals underwent marrow needle insertion and then saline injection into the carotid artery. Animals were assessed on a battery of neurological tests. MNCs in the ischemic brain were tracked using Q-dot nanocrystal labeling. Infarct volume and cytokines in the ischemia-affected brain were analyzed. Cell-treated animals in the younger and older groups showed improvement from 7 to 30 days after stroke compared with vehicle-treated animals. MNCs significantly reduced infarct volume compared with saline. There was a significant reduction in tumor necrosis factor-α, interleukin-1α (IL-1α), IL-β, IL-6, and a significant increase in IL-10 in injured brains harvested from the cell-treated groups compared with saline controls. Labeled MNCs were found in the peri-infarcted area at 1 h and exponentially decreased over the ensuing week after injection. Autologous bone marrow MNCs can be safely harvested from rodents after stroke, migrate to the peri-infarct area, enhance recovery, and modulate the post-ischemic inflammatory response.
Keywords
Introduction
Cellular therapy has emerged as a potential novel neurorestorative approach for the treatment of ischemic and hemorrhagic stroke. Cells originating in the bone marrow are one of many different cellular sources that have been shown to improve outcome in animal models of middle cerebral artery occlusion (MCAo). For example, bone marrow stromal cells (BMSCs), which exhibit the properties of multi-potential stem cells, reduce neurological deficits in rodent models of stroke (Chen et al, 2001). BMSCs are easily isolated from the bone marrow because they adhere to plastic, can be grown and scaled to sufficient quantities in culture, and rodents injected with BMSCs show no evidence of adverse events up to one year after injury (Shen et al, 2007). The safety of BMSCs has even been investigated in five stroke patients in Korea. In that study, these cells were harvested from the stroke patient's own bone marrow and required 30 days to grow to a pure population of sufficient quantity for autologous intravenous administration(Bang et al, 2005). A pure population of autologous BMSCs at the present time is, therefore, not a practical therapeutic option for acute or subacute stroke.
An alternative bone marrow population is the mononuclear cell (MNCs) fraction from which MSCs are purified. MNCs contain a population of mesenchymal and hematopoietic stem cells, which secrete a host of cytokines and growth factors involved in natural repair processes (Takahashi et al, 2006). They can be rapidly derived from the bone marrow of animals and patients, separated and isolated within 3h in our experience and then returned back to the animal or patient. MNCs have been shown to improve outcome in animal models of cerebral ischemia (Iihoshi et al, 2004; Kamiya et al, 2008). Preliminary studies even suggest that autologous MNCs can improve cardiac function in endstage heart failure by possibly upregulating endogenous repair responses (Perin et al, 2003), and they have been shown to improve outcome with variable results in patients with acute myocardial infarction (Kamihata et al, 2001; Strauer et al, 2002). Autologous MNCs are, therefore, a potentially attractive candidate to promote stroke recovery.
No studies have examined whether MNCs can be safely extracted from the bone marrow of animals after ischemic stroke and whether autologous intraarterial (IA) delivery at 24 h after stroke in a clinically relevant manner leads to enhanced recovery. Herein, we present an animal model of focal ischemic stroke in which MNCs were extracted from the bone marrow of the tibia of young and middle-aged Long Evans rats at 24 h after MCAo and then were autologously administered via carotid injection within 2 h after bone marrow harvest. We hypothesized that bone marrow could be safely removed from young and older rats after ischemic stroke, MNCs could then be rapidly isolated from this bone marrow, re-injected autologously, and would reduce infarct volume and neurological deficits in young and older animals. We also quantified the presence of labeled MNCs in the injured brain and present another mechanism underlying the potential benefits of MNCs in rodent stroke by assessing their effects on the post-ischemic inflammatory response.
Materials and methods
MCA Occlusion
Focal ischemia of 180-min duration in male Long Evans rats that were 350 g (3 months old) or 700 g (12 months old) was induced by tandem left middle cerebral artery (MCA) and left common carotid artery (CCA) occlusion (CCAo/MCAo), as described previously (Aronowski et al, 1997). For these procedures, animals were anesthetized with chloral hydrate (0.45 g/kg). In brief, the femoral artery was cannulated for blood pressure (BP) and blood gas recording. The temperature of the temporalis muscle was monitored/controlled using a feed-forward temperature controller. The CCA was isolated through a midline incision and tagged with a suture. Two burr holes were made in the skull: a 1 × 2.5-mm rectangular burr hole to expose the left MCA and a 1-mm-diameter burr hole to facilitate local cerebral perfusion (CP) measurement. CP in rat can be measured directly through the drill-thinned skull. A 0.005-inch diameter stainless steel wire was placed underneath to occlude the left MCA rostral to the rhinal fissure, proximal to the major bifurcation of the MCA, and distal to the lenticulostriate arteries. The left CCA was then be occluded using Heifetz aneurysm clips. Interruption of local blood flow through the MCA was inspected under the microscope and verified with a laser Doppler flowmeter (LDF) placed over the ischemic area at 2 mm posterior and 6 mm lateral to the bregma (Aronowski et al, 1997). At 24 h after stroke, local CP using LDF was measured over the penumbra area (3.8 mm posterior and 4 mm lateral from the bregma) in a subset of animals that underwent either MNC or saline injection (Aronowski et al, 1997) (
Bone Marrow Harvest
We were able to perform bone marrow harvest in rats the day after they have undergone CCAo/MCAo. The rats were anesthetized with an intraperitoneal (IP) injection of chloral hydrate, 0.4 g/kg. An incision was made through the skin to the medial aspect of the tibia. The periosteum was removed and the surgeon drilled a 1.25 × 2.5-mm burr hole extending into the medullary cavity. A 20-gauge hypodermic needle was inserted into the medullary cavity and connected to a heparinized syringe. Bone marrow (1 to 1.5 ml) was aspirated while rotating and moving the needle back and forth. The medullary cavity was flushed with saline and the content aspirated. The burr hole was sealed with bone wax and the skin closed with a nylon suture.
Animal Groups
All animals underwent CCAo/MCAo and then 22 h later bone marrow procedure. Animals were then randomly assigned to MNC treatment or control. Those animals assigned to the MNC group underwent harvest as described above and then received MNCs via carotid injection. Young animals received 10 million cells while older animals received 4 million cells. A lower dose of cells was chosen for older animals based on calculations of MNC yield from bone marrow harvested in older patients with congestive heart failure who are currently undergoing MNC administration in a clinical trial at our center. Four million cells was, therefore, selected to choose a dose of cells that we believe would be feasible to obtain from a bone marrow harvest in older patients with vascular disorders. Those animals assigned to the control group underwent sham bone marrow procedure consisting of needle injection and marrow flushing, but marrow was not harvested; they then received saline via carotid injection at 24 h after MCA occlusion. This control group was designed based on the idea that if cells were removed from the bone marrow, they would not be returned; therefore, instead, we elected to insert the needle into the bone marrow without harvest. In preliminary experiments, we compared animals that underwent CCA/MCAo and bone marrow harvest with animals that only underwent CCA/MCAo and found no differences in behavior on a battery of neurological tests.
Bone Marrow Cell Processing
The cells from the bone marrow aspirate were triturated, centrifuged, and washed in phosphate-buffered saline + 0.5% bovine serum albumin. Cells were then suspended in Media 199 and counted using a hemocytometer and Coulter counter. The cell suspension was added on top of 20 mL Ficoll–Paque PLUS in a 50 mL conical vial and then centrifuged. The MNCs were collected, washed with phosphate-buffered saline+ 0.5% bovine serum albumin (or media 199), and counted. Cells were suspended in sterile, cold phosphate-buffered saline at the desired concentration. The overall procedure took 2 h to complete.
Characterization of the MNC Population
We performed flow cytometry to characterize the MNC population using a previously published protocol (Harting et al, 2008).
Delivery Route
For IA delivery, under a surgical microscope the left CCA, left external carotid artery (ECA) and the left internal carotid artery (ICA) were isolated via a midline incision. An aneurysm clip ligation of the CCA and ICA was performed. The ECA was ligated with a nylon suture and nicked at a 45-degree angle. A modified PE10 catheter was inserted into the ECA and advanced so that the tip sat into the ICA. The ligature was tightened around the stump of the ECA until hemostasis was achieved. The ICA was unclipped and marrow cells or saline infused slowly over 5 mins. The catheter was then flushed with saline. The catheter was withdrawn and the ligature was tightened around the stump of the ECA. The skin incision was closed with a nylon suture.
Analysis of Long-Term Motor Dysfunction
All sensorimotor testing was performed during the light cycle. Animals were pre-tested and then tested on days 1, 7, 14, 21, and 28 post-ischemia. We used a battery of sensorimotor tests sensitive to cortical damage produced by our rodent ischemia model and include the following:
Asymmetry in the Use of Forelimbs for Postural Support (Cylinder Test):
Animals were placed into a plexiglas cylinder and their behavior observed for forelimb-use asymmetry during vertical movements along the wall of the cylinder. The final score was calculated as (non-impaired forelimb movement—impaired forelimb movement)/(non-impaired forelimb movement + impaired forelimb movement + both movement), as previously described in the rat (Schallert et al, 2000). A total of 20 movements were recorded during the 10-min test.
Asymmetry-Corner Test:
In the home cage, an animal was placed between two angled boards. When entering deep into the corner, both sides of the vibrissae are stimulated together. The animal then rears forward and upward, then turning back to face the open end. Twenty trials were performed for each rat and the percentage of left turns versus right turns was calculated. Only turns involving full rearing along either board were recorded.
Infarct Volume
Young animals (
Cytokine Analysis
A separate group of young animals (
Assessment of Transplanted MNCs
To track the transplanted cells in the brain, Q-dot nanocrystals, a red fluorescent maker (655 nm), was used. MNCs were harvested and labeled with Q-dot as described previously (Rosen et al, 2007). Transient CCAo/MCAo and cell administration were performed as described above (
TUNEL Analysis
The terminal deoxynucleotidyltransferase-mediated dUTP-biotin nick-end labeling (TUNEL) method was performed to assess cell death by using the ApoTag Fluorescein
Statistical Analysis
Data are presented as mean ± s.d. Cytokine analysis was analyzed using Student's
Results
Bone Marrow Harvest and MNC Yield
Bone marrow harvest after stroke did not cause hemodynamic compromise based on blood pressure and heart rate (data not shown), and there was no observed increase in mortality for the study period. We were able to conduct autologous MNC treatment in all animals randomized to receive cells. Needle aspiration from one tibia 22 h after stroke consistently without exception yielded over 10 million MNCs from one rat (range 11 to 30 million). The procedure did not cause limb impairment that we could observe in any of the animals that underwent a bone marrow procedure. All animals bore their own weight on the ipsilateral limb after harvest; thus, animals fully participated in our behavioral studies.
Phenotypic Characterization
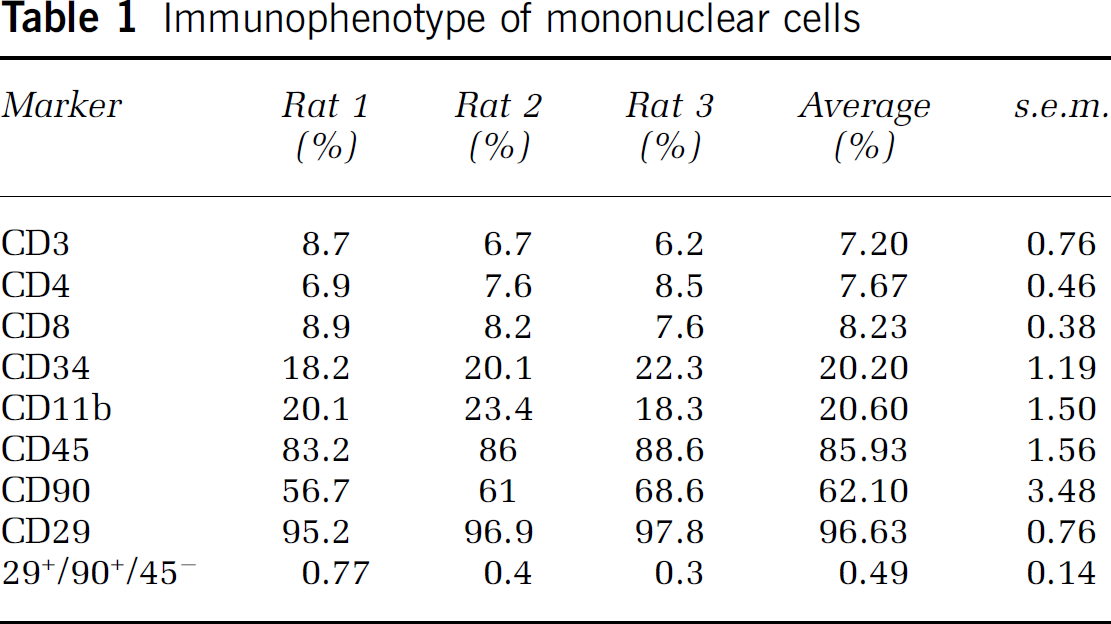
The MNC fraction of bone marrow is a mixture of different cellular populations. We provide immunophenotypic characterization of MNCs in Table 1, which gives the breakdown of cell-surface markers of mononuclear cells derived from the bone marrow of young Long Evans rats. The mesenchymal stem cell (MSC) population in the MNC fraction was approximated by assessing the percentage of cells that were CD29+/90+ /CD45−.
Immunophenotype of mononuclear cells
Intra-arterial Delivery Enhances Recovery in Young Rats
Long Evans rats underwent left CCA/MCA occlusion for 3 h, which consistently led to right-hemibody deficits. MNCs (10 million cells) or vehicle were injected at 24 h after CCAo/MCAo into the ipsilateral carotid artery. All animals survived to the study endpoint and underwent serial neurological testing throughout the study period. Neurological testing demonstrated that both groups of animals were impaired before injection of cells or saline. In the vehicle-treated group, deficits persisted in both the cylinder (0.48 ± 0.11) and corner tests (90% ± 5% turns to the left) at 30 days. The cell-treated young animals began to show improvement by 3 to 7 days after stroke and their deficits resolved on both tests by 30 days (cylinder, 0.02 ± 0.15 and corner, 50% ± 6% (
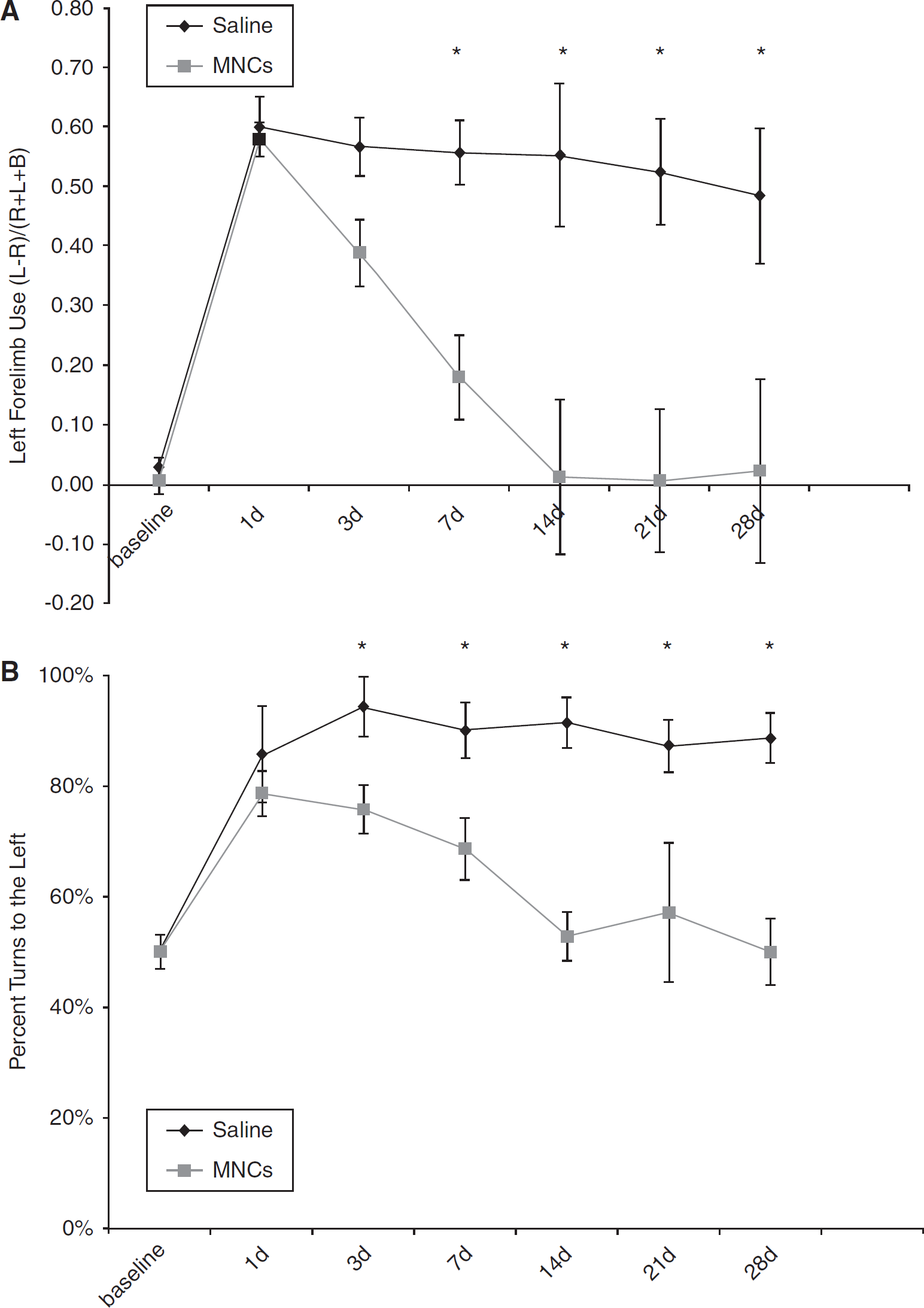
Young Long Evans rats treated with MNCs have improved recovery after cortical stroke. Animals underwent CCAo/MCAo, and 24h later either received an IA injection of autologous MNCs or saline followed by neurological testing for 30 days. (
Intra-arterial Delivery Enhances Recovery in Older Rats
Retired breeder Long Evans rats underwent left CCA/MCA occlusion for 3 h, which consistently caused right-hemibody deficits. MNCs (4 million cells) or saline vehicle were injected at 24 h after focal ischemia. Among the older animals, cell-treated rats (
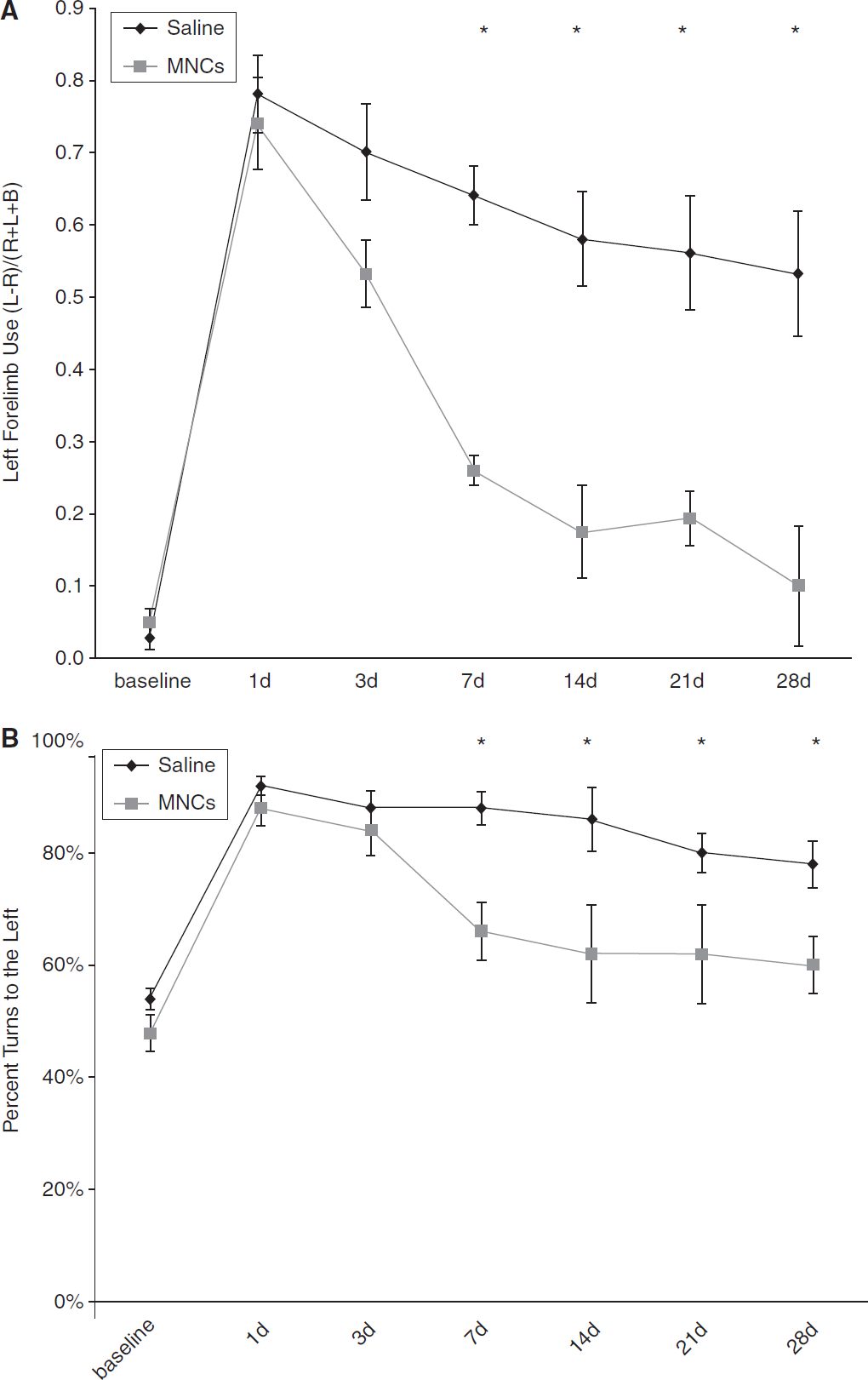
Twelve-month-old, Long Evans rats treated with MNCs show improved recovery after cortical stroke. Retired breeders underwent CCAo/MCAo, and 24h later either received an IA injection of autologous MNCs or saline. Saline-treated rats show persistent severe deficits lasting for at least 28 days as determined using cylinder (
Physiological Monitoring During Cell Infusion
We monitored retired breeder animals (
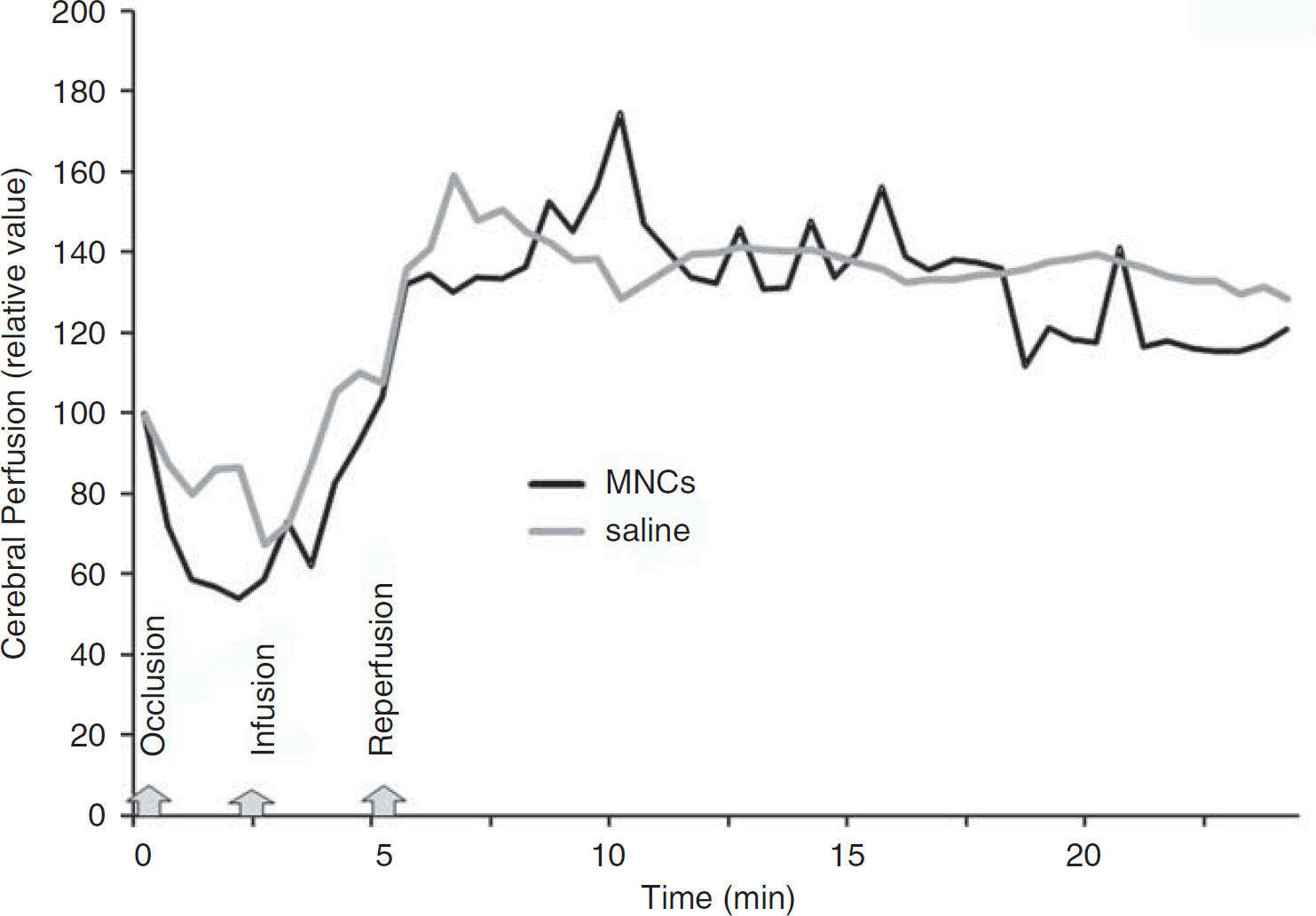
Cerebral perfusion during IA infusion of MNCs or saline. At 24h after acute stroke, the external carotid and common artery were ligated, which corresponded with a decline in cerebral perfusion as usual in this model (labeled occlusion). Then a catheter was inserted to the ICA to infuse saline or MNCs in a final volume of 1 ml (labeled infusion). In response to the infusion of MNCs or saline, the cerebral perfusion increased in all animals tested beyond baseline. The CCA was unclamped (labeled reperfusion) and then there was a return of cerebral perfusion to mildly elevated levels in both saline- and MNC-treated groups. Data represent the mean of four animals per group.
MNCs in the Brain After IA Delivery
We found labeled MNCs in the per-infarct regions as early as 1h after IA injection. Figure 4 shows representative photomicrographs of brain sections demonstrating fluorescence-labeled MNCs in the peri-ischemic areas of the brain 1h to 7 days after IA administration. There was an exponential decrease in the number of labeled MNCs in the peri-infarct area (Figure 4E). No labeled cells were observed in the contralateral hemisphere or cerebellum (data not shown). Immunohistochemistry with TUNEL showed that some Q-dot-labeled MNCs were TUNEL positive (Figure 4F to 4H) at 3 h after injection.
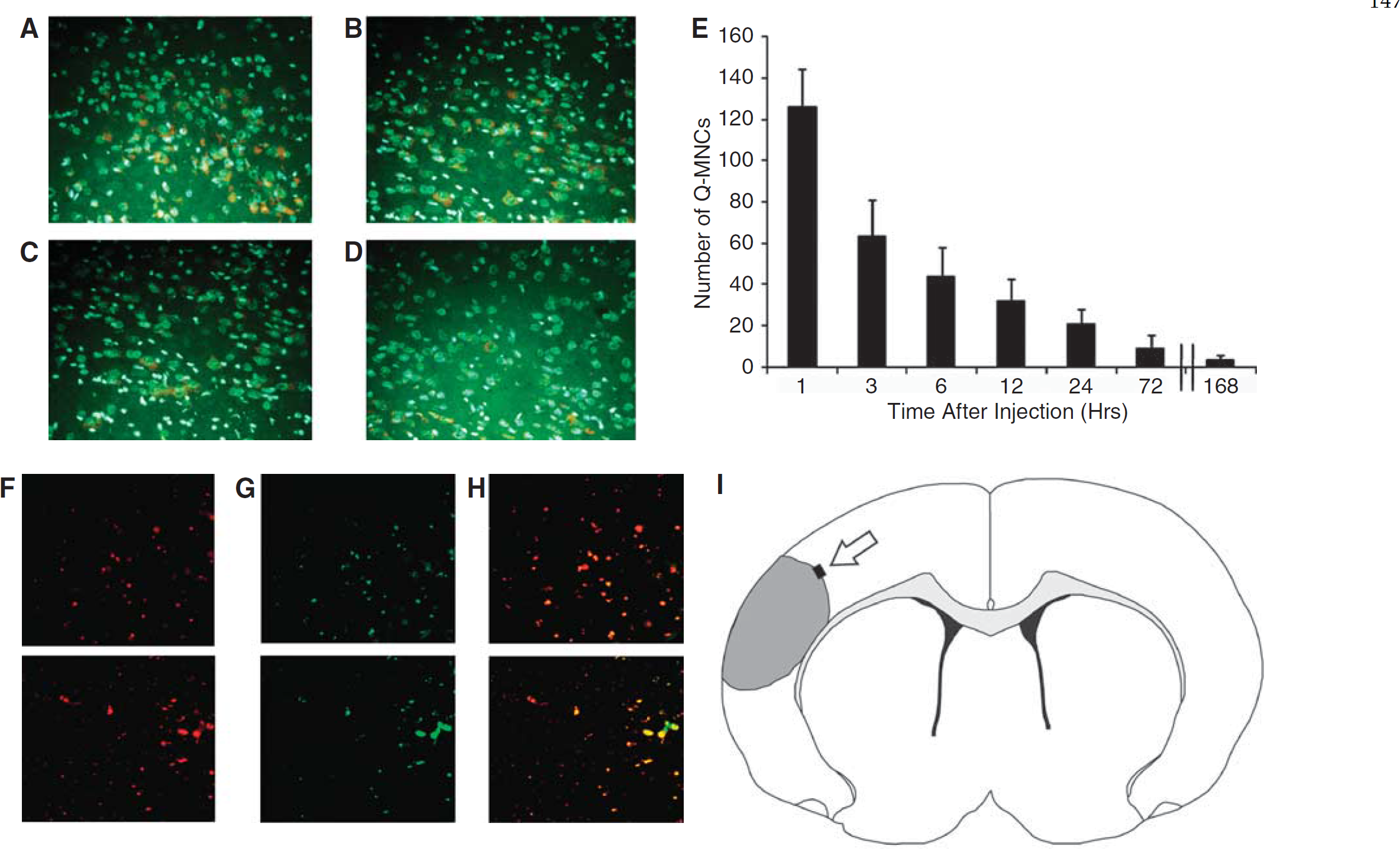
Twenty-four hours after CCAo/MCAo, Long Evans rats received an intra-carotid injection of MNCs. The MNCs were derived from the bone marrow and labeled with Q tracker nanocrystals before infusion. Fluorescence microscopic images illustrate time-dependent elimination of MNCs in per-infarcted rat brain. One hour (
Infarct Volume
Infarction size was corrected for edema. The rats treated with MNCs showed significant reduction in infarction size when compared with saline control animals (
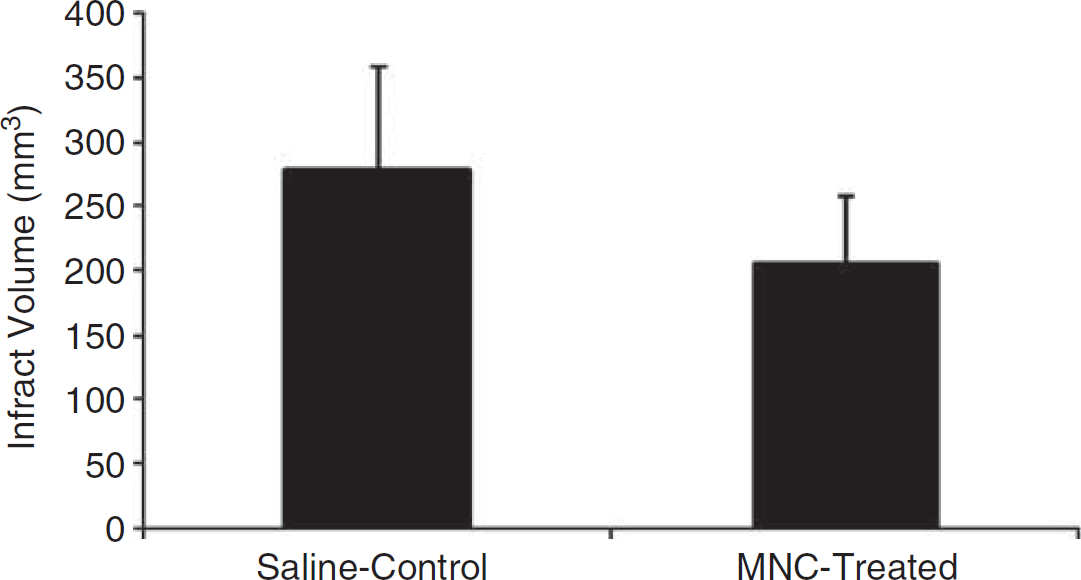
A bar graph exhibiting significantly (
Cytokine Analysis
At 48h after stroke in young animals, tissue harvested from the ischemic hemisphere had significantly lower levels of IL-1α, IL-1β, IL-2, tumor necrosis factor-α, and IL-6 in cell-treated animals compared with saline-treated animals (
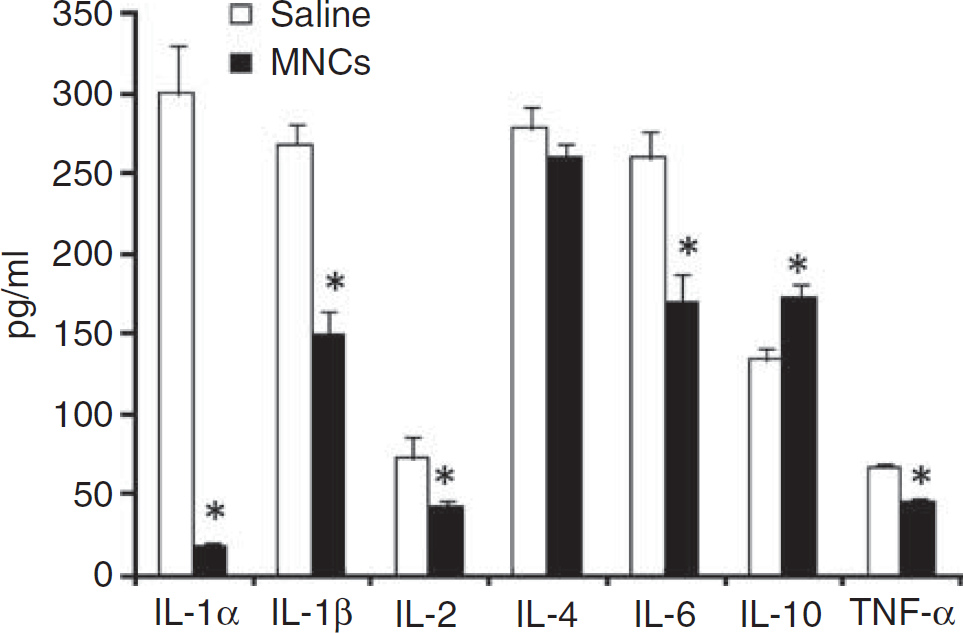
Ischemic brain tissue after stroke treated with MNCs or saline was homogenized, centrifuged, and then the supernatant was analyzed using a multiplex bead-based enzyme-linked immunosorbant assay for the presence of selective cytokines. As shown, levels of several proinflammatory cytokines were significantly lower (
Discussion
Acute IA infusion of autologous MNCs after stroke reduced neurological deficits in young and older rats. MNCs were found in the peri-infarct area within 1h after injection, but decreased within hours and were nearly undetectable by 7 days afterwards. MNCs modulated post-ischemic inflammatory cytokines within the brain and reduced infarct volume.
In this clinically relevant rodent model, we were able to extract over 10 million MNCs from the tibia of rats after they had a stroke without causing mortality or limb impairment. Injection of MNCs within hours of harvest did not alter cardiovascular hemodynamics (data not shown) or cerebral perfusion (Figure 3). MNC-treated animals showed significant improvement in neurological deficits on two different behavioral tests. Impairment was reduced by 7 days after injection in the young animals and recovery was sustained over a 30-day period. Among older animals administered a lower number of cells, recovery was also enhanced in the cell-treated animals. Normally, 12-month-old, Long Evans rats are close to a period of time in their lifespan when they begin to develop comorbidities such as cardio-myopathy; mortality begins to increase around 14 months of age, according to our experience. Therefore, our results may suggest the possibility that both younger and older animals with stroke derived benefit from autologous MNC therapy.
Previous studies have investigated the therapeutic effects of MNCs in focal cerebral ischemia (Baker et al, 2007; Iihoshi et al, 2004; Kamiya et al, 2008), but differ from the present report in several ways. First, we show that older animals with stroke also derive benefit from autologous MNCs. Second, in the prior studies, MNCs were extracted several days before the stroke and re-injected 6h after MCA occlusion (Baker et al, 2007) or extracted 1h before stroke and re-injected at reperfusion (Kamiya et al, 2008). To our knowledge, this is the first clinically relevant study to show the feasibility of harvesting MNCs from the bone marrow at 22h after ischemic stroke, rapidly isolating them, and then immediately re-injecting the cells with an IA delivery at 24 h after stroke. We chose 24h in contrast to these prior studies because this is a more clinically practical time point to harvest bone marrow from patients when they are out of the window for thrombolytic or neuroprotective therapies. We also chose saline as a control given that some clinical studies testing mononuclear cells have used saline in their control group (Yao et al, 2008); however, we have also tested the effects of autologous dead MNCs, which do not reduce neurological deficits in our stroke model (data not shown).
Despite the concern for edema formation, brain inflammation, and emboli, our study supports the feasibility and safety of performing IA cell delivery in the acute setting of ischemic stroke. A recent report, however, found that intra-carotid delivery of MSCs in some animals led to cerebral blood flow reduction and mortality (Walczak et al, 2008). There are several differences that distinguish our work from this study. First, MSCs grown in culture for days to weeks likely adhere to each other and, therefore, can predispose to emboli. Injection of these cells could, therefore, lead to microembolization within the cerebrovasculature. MNCs, conversely, are not grown in culture, do not adhere to each other, and they did not lead to a decrease in cerebral blood flow in the animals that were studied (Figure 3). Second, MSCs have an average cell size of at least 12 to 18 μm whereas MNCs have an average cell size of 7 μm (Fischer et al, 2008). Third, the prior study used an endovascular suture to occlude the arterial lumen, thereby potentially injuring the arterial wall, causing the cells to adhere to a damaged endothelial surface and obstruct the lumen. Our ischemia model involves a focal occlusion of the MCA with a wire through a small craniotomy; however, any manipulation of the vasculature can induce vessel damage.
We chose an IA approach because it is less invasive than an intracranial approach and is clinically relevant to acute ischemic stroke with the intention to focus cell delivery to the area of pathology. Intravenous delivery has been shown to deposit bone marrow cells in the liver, spleen, and lungs (Chen et al, 2001). In addition, our previous work has shown that more purified cells such as MSCs, when administered intravenously, can be trapped in the lung (Fischer et al, 2008). We, therefore, bypassed the first pass filter of the lung with an intra-carotid injection. This delivery method led to immediate deposition of cells in the peri-infarct area, but labeled cells exponentially decreased to minimal levels in the peri-infarct region over the ensuing week after injection. This is the first report to provide a kinetic analysis of labeled MNCs in the injured brain over time, and we did see evidence that labeled MNCs were TUNEL positive at 3h after injection, indicating that MNCs are dying after they migrate to the area of injury. One explanation may be that the injured brain poses a toxic environment to MNCs.
The mechanisms underlying cell-therapy-induced recovery are unclear. Prior studies suggest that MNCs decrease infarct volume (Baker et al, 2007; Iihoshi et al, 2004; Kamiya et al, 2008) and increase angiogenesis and vessel density in the peri-infarct areas of the brain (Baker et al, 2007). Other studies suggest that some MNCs show neural markers in and around the ischemic lesion after intravenous injection in the rodent stroke model (Iihoshi et al, 2004). The early recovery seen in our study argues against angiogenesis or neurogenesis as the predominant mechanisms underlying the efficacy of MNCs in reducing neurological deficits. We found a significant reduction in the levels of several proinflammatory cytokines in the ischemic brains harvested from rodents treated with MNCs as compared with saline controls. In addition, IL-10 level was significantly increased in cell-treated animals as compared with saline controls. Immunomodulation may, therefore, be another important mechanism underlying the effects of intra-arterially administered MNCs in the post-ischemic brain, similar to what has been reported for other types of cellular therapeutics (Vendrame et al, 2005). However, we also found that MNCs reduce infarct volume when administered at 24 h after stroke. It is, therefore, possible that the reduction in inflammatory cytokines may be secondary to reduced tissue injury. The few cells observed in the peri-infarcted areas by the time improvement in neurological deficits was seen suggest that these cells may release factors to exert cytoprotective and immunomodulatory effects and then die off rapidly in the peri-infarcted regions. Other studies have also shown that tissue protection is still possible to achieve even when cellular treatments are administered within the first 1 to 2 days after stroke. It has been shown that there is ongoing injury in the peri-infarct areas through such mechanisms as apoptosis, which contribute to infarct maturation over the ensuing days after stroke (Vendrame et al, 2004). Further work is, therefore, needed to determine a therapeutic time window and dose response of MNCs in ischemic stroke.
Footnotes
The authors declare no conflict of interest.