
Abstract
The choroid plexus, being part of the blood-brain barriers and responsible for the production of cerebrospinal fluid, is ideally positioned to transmit signals into and out of the brain. This study, using microarray analysis, shows that the mouse choroid plexus displays an acute-phase response after an inflammatory stimulus induced in the periphery by lipopolysaccharide (LPS). Remarkably, the response is specific to a restricted number of genes (out of a total of 24,000 genes analyzed, 252 are up-regulated and 173 are down-regulated) and transient, as it returns to basal conditions within 72 h. The up-regulated genes cluster into families implicated in immune-mediated cascades and in extracellular matrix remodeling, whereas those down-regulated participate in maintenance of the barrier function. Importantly, several acute-phase proteins, whose blood concentrations rise in response to inflammation, may contribute to the effects observed
Keywords
Introduction
The precise mechanisms through which peripheral inflammatory stimuli trigger brain inflammation are poorly understood. Interest on the subject is increasing in light of the importance of inflammation in the central nervous system (CNS) in multiple sclerosis (MS), and the more recent implications of CNS inflammation in neurodegenerative diseases such as Parkinson and Alzheimer diseases.
Although most studies addressing communication between the periphery and the CNS focus on the blood-brain barrier (BBB) formed by endothelial cells of the brain capillaries, considerably less attention has been paid to the involvement of the blood-cerebrospinal fluid (CSF) barrier (BCSFB), formed by the choroid plexus (CP) epithelial cells. The CP, located within the cerebral ventricles, is composed of a vascularized stroma surrounded by a tight layer of epithelial cells that restrict cellular and molecular traffic between the blood and the CSF (Redzic and Segal, 2004). The CP is best known for its role in the production of the CSF that fills the brain ventricles and the subarachnoid space, and it is ideally located to transmit signals into and out of the brain (Emerich et al, 2005).
To date, studies published on the CP response to peripheral inflammatory stimuli have mostly focused on single proteins or groups of proteins. Among these are immune mediators such as interleukin IL-1β and TNF (Nadeau and Rivest, 1999; Quan et al, 1999), enzymes such as prostaglandin D2 synthase (Marques et al, 2007) and bacteriostatic proteins such as lipocalin 2 (LCN2) (Marques et al, 2008). Accessory molecules important for leukocyte adhesion such as
Materials and methods
Animals and Lipopolysaccharide Injection
All experiments were carried out using 8 to 9 weeks old C57BL/6 male mice and 8 to 9 weeks old Wistar rats (Charles River, Barcelona, Spain), in accordance with the European Community Council Directive 86/09/EEC guidelines for the care and handling of laboratory animals. Animals were maintained under 12 h light/dark cycle at 22.5°C and 55% humidity and fed with regular rodent's chow and tap water
The CSF was collected from the cisterna magna and pooled from several mice. An aliquot of each pool was used to verify the absence of blood contamination and the remainder immediately frozen until use.
Rats were similarly anesthetized and killed 3 and 6h after LPS injection. Blood was collected and used to stimulate primary cultures of rat CP epithelial cells.
Microarray Experimental Design and Data Analysis
Total RNA was isolated with Trizol (Invitrogen, Carlsbad, CA, USA) following manufacturer's instructions. After quality assessment using the Agilent Bioanalyzer (Agilent Technologies, CA, USA), 100 ng of total RNA from three pooled controls and two pooled samples from each time point were amplified and labeled with Illumina TotalPrep RNA Amplification Kit (Illumina Inc, San Diego, CA, USA). The labeled cRNA was then hybridized using the recommended protocol in a total of two Illumina Whole-genome Mouseref-8 expression Beadchips (Illumina Inc). This mouse beadchip contains eight arrays, each comprising a total of 24,000 well-annotated RefSeq transcripts.
After scanning, raw data from BeadStudio software (Illumina Inc) was read into R/Bioconductor and normalized using quantile normalization. A linear model was applied to the normalized data using Limma package in R/Bioconductor (Gentleman et al, 2004). A contrast analysis was applied and differentially expressed genes were selected using a Bayesian approach with a false discovery rate of 5%.
The differentially expressed genes were categorized using Gene Ontology from Biomart (http://www.biomart.org/) or Ingenuity tools (Redwood City, CA, USA). Enrichment analysis was performed using the DAVID (http://david.niaid.nih.gov/david/ease.htm) and the Ingenuity softwares.
Gene Expression Measurements by Quantitative Real-Time-Polymerase Chain Reaction
As described earlier, 500 ng of total RNA isolated were amplified using a Superscript RNA Amplification System (Invitrogen) according to the manufacturer's instructions. After amplification, RNA was reverse transcribed into first strand cDNA using random hexamers of the Superscript First-strand Synthesis System for RT-PCR (Invitrogen).
The qRT-PCR analysis was used to measure the expression levels of selected mRNA transcripts. Primers were designed using the Primer3 software (Rozen and Skaletsky, 2000) on the basis of the respective GenBank sequences. The expression level of the reference gene hypoxanthine guanine phosphoribosyl transferase (
All other accession numbers and primer sequences are available on request. Reactions using equal amounts of total RNA from each sample were performed on a Light-Cycler instrument (Roche Diagnostics, Basel, Switzerland) with the QuantiTect SYBR Green RT-PCR reagent kit (Qiagen, Hamburg, Germany) according to the manufacturer's instructions. Product fluorescence was detected at the end of the elongation cycle. All melting curves exhibited a single sharp peak at the expected temperature.
Cerebrospinal Fluid Analysis
Immune mediators (IL-6, IL-10, IL-12p70, TNF, IFNγ, and CCL2) were measured using BD cytometric bead array (BD Biosciences, CA, USA) according to the manufacturer's instructions in 10 μL of pooled CSF samples obtained from animals injected with LPS at different time points. Three to four pools of CSF from up to five animals each were used for the various time points studied. The detection limit for all proteins was 20pg/mL.
Primary Cultures of Rat Choroid Plexus Epithelial Cells
The CP is composed of a vascularized stroma surrounded by a tight layer of epithelial cells. Therefore, to study to what extent the response observed
Epithelial cells from rat CP were prepared as described previously by Strazielle and Ghersi-Egea (1999) with minor modifications. Briefly, neonates (postnatal day 3 or 4) were killed and CP were dissected under conventional light microscopy (SZX7, Olympus). The tissue was rinsed twice in phosphate buffered saline (without Ca2+ and Mg2+) followed by a 25-min digestion with 0.1 mg/mL pronase (Sigma) at 37°C. Predigested tissue was recovered by sedimentation and briefly shaken in 0.025% of trypsin (Invitrogen) containing 12.5μg/ml DNAsel (Roche). The supernant was then withdrawn and kept on ice with 10% fetal bovine serum (Invitrogen). This step was repeated five times. Cells were pelleted by centrifugation and ressup-ended in culture media consisting of Ham's F-12 and DMEM (1:1) (Invitrogen) supplemented with 10% fetal bovine serum, 2 mmol/L glutamine (Invitrogen), 50μg/mL gentamycin (Sigma), 5 μg/mL insulin, 5 μg/mL transferrin, 5 ng/mL sodium selenite (ITS, Sigma), 10 ng/mL epidermal growth factor (Sigma), 2 μg/mL hydrocortisone (Sigma), 5 ng/mL basic fibroblast growth factor (Invitrogen). For further enrichment, cells were incubated on plastic dishes for 2h at 37°C. A differential attachment on plastic dish occurred, and supernant containing mostly epithelial cells was collected and placed for seeding on laminin (Boehringer Ingelheim, GmbH, Germany) coated transwells (Corning, Lowell, MA, USA). To assess purity, the cell monolayers were immunostained with an anti-transthyretin antibody (specific for CP epithelial cells) (kindly provided by Maria João Saraiva). Cell counting under the microscope revealed that at least 95% of the cells stained positive for transthyretin, thus confirming the purity of the cultures.
Stimulations were performed after the formation of confluent cell monolayers, approximately after 7 days in culture. The CP epithelial cells were stimulated with LPS (200 ng/mL) in the basal side (which corresponds to the membrane facing the blood
A total of 1,000 ng RNA were reverse-transcribed into first strand cDNA using Oligo-dt from the Superscript First-strand Synthesis System for RT-PCR (Invitrogen). The qRT-PCR analysis was used to measure the expression levels of selected mRNA transcripts.
Statistical Analysis
The values are reported as mean ± s.e. Statistical significance was determined using the nonparametric Mann-Whitney test, with differences considered significant at
Results
Reproducibility of the Gene Array Data
Two pooled CP samples from animals injected with LPS and killed at 1, 3, 6, 12, 24, or 72 h were compared with three pooled CP samples from saline-injected animals. The data were analyzed in the R-Bioconductor. Quality control using inter-array Pearson correlation and clustering based on variance allowed us to ensure reproducibility between the replicates (data not shown). After data normalization, differentially expressed genes were selected based on a false discovery rate of 5%. This analysis yielded a list of 252 up-regulated and 173 down-regulated genes at one or more of the experimental time points.
Kinetic Profile of the Choroid Plexus Transcriptome After a Peripheral Inflammatory Stimulus
Figure 1 depicts the number of genes whose expression was up-regulated or down-regulated throughout the experimental period; it shows that the CP displays a rapid (acute phase) response to LPS that peaked at 3 to 6 h after LPS administration and that gradually returned to baseline by 72 h.
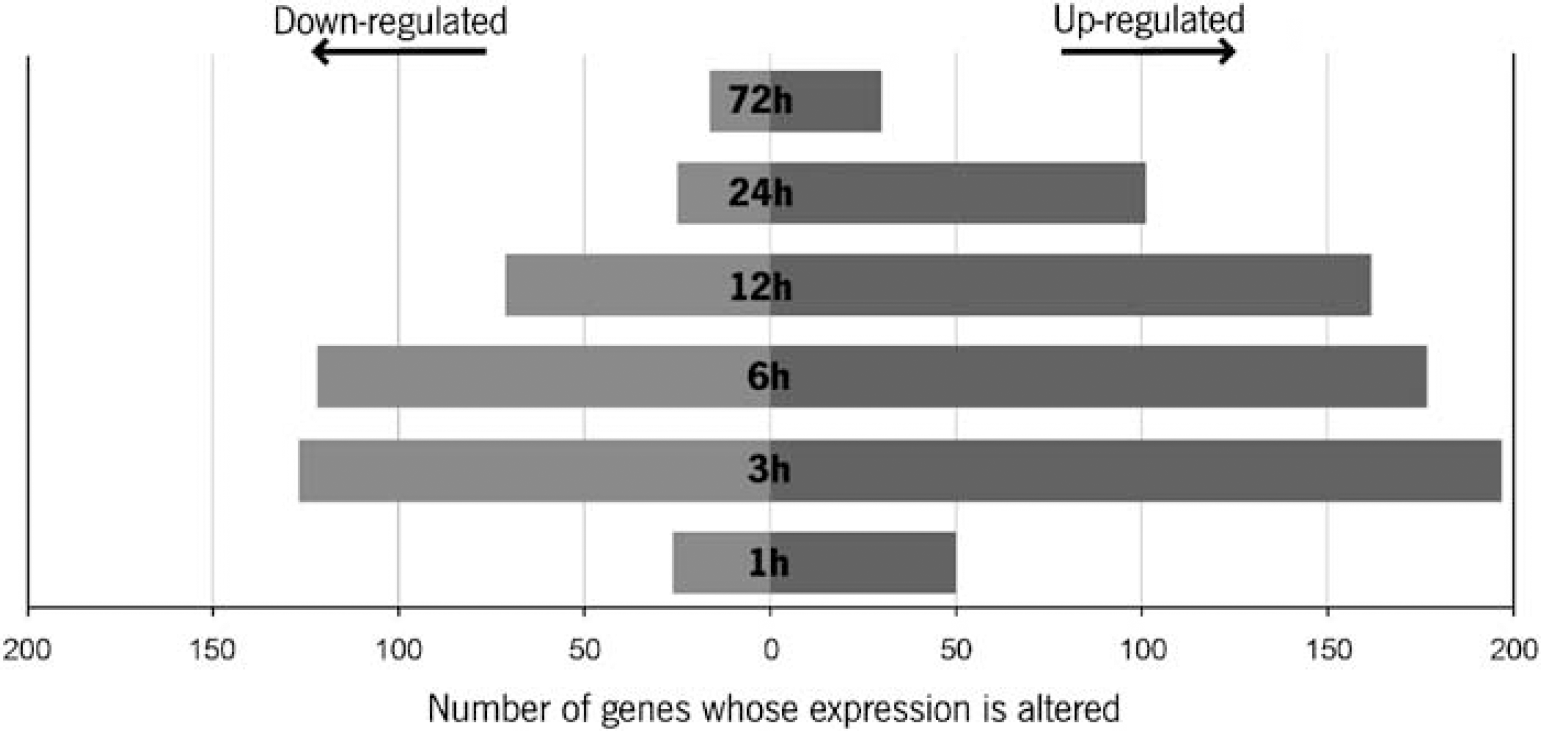
Kinetic profile of the CP response to LPS. The number of genes whose expression was found altered at 1, 3, 6, 12, 24, and 72 h after the peripheral injection of LPS as compared with saline-injected mice. The genes whose expression was up-regulated are represented in black, and those that were down-regulated are represented in grey.
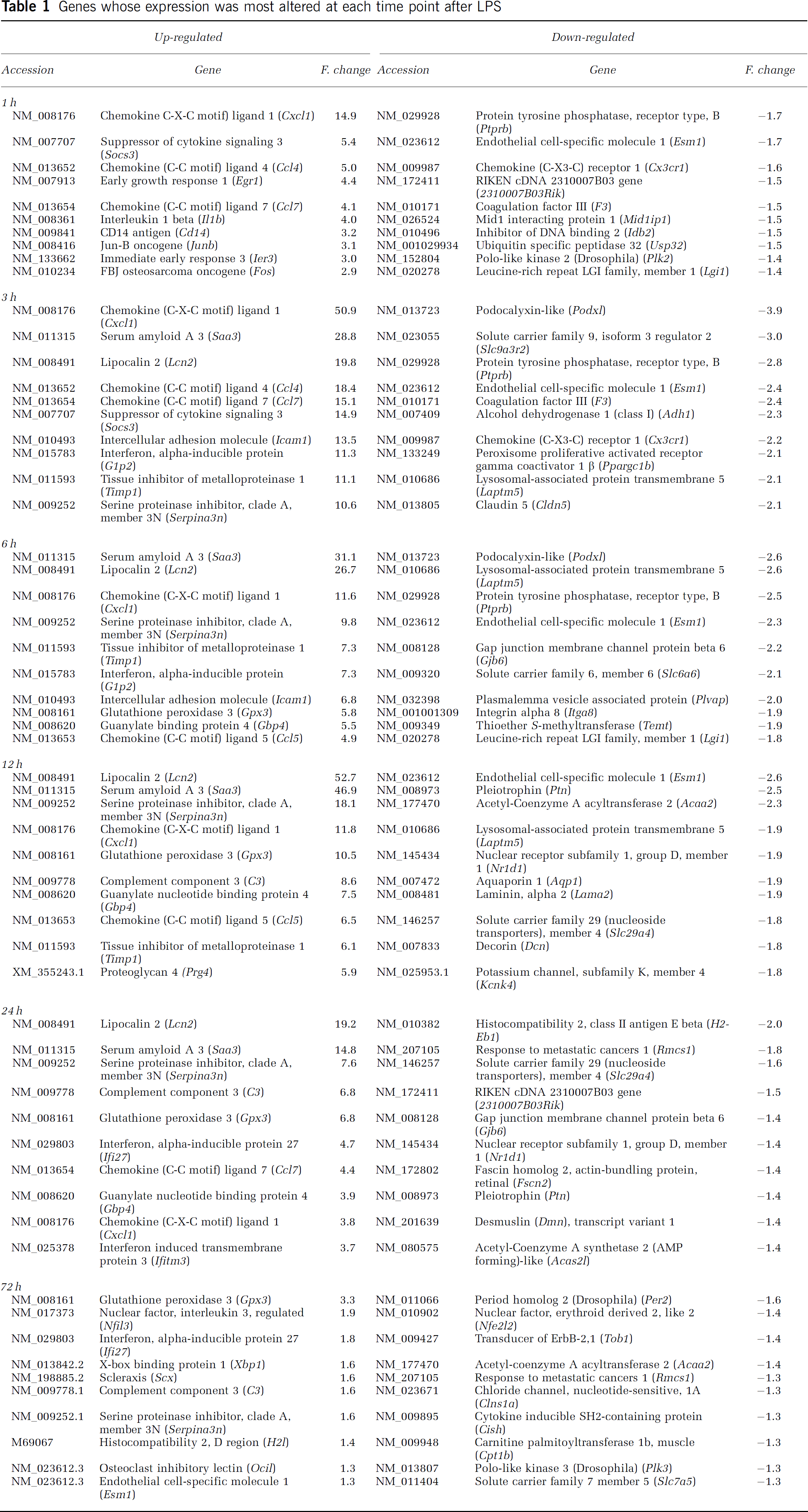
When genes were grouped with respect to the fold changes (Table 1), it became evident that the fold change was of higher magnitude for up-regulated genes (complete list of genes is available as Supplementary data Table S1).
Genes whose expression was most altered at each time point after LPS
Identification of Altered Gene Pathways
Gene ontology and biological pathway analyses of differentially expressed genes, performed using the Ingenuity software and the DAVID program, showed that the biological pathways mostly altered are associated with the innate immune response (Table 2).
Clustering of the genes whose expression was altered in the choroid plexus upon peripheral LPS injection

denotes up-regulated
denotes down-regulated.
Confirmation of Array Results by Quantitative Real-Time-Polymerase Chain Reaction on a Set of Relevant Genes
Within each pathway, and using RNA extracted from CP pools of an independent experiment, a number of genes (
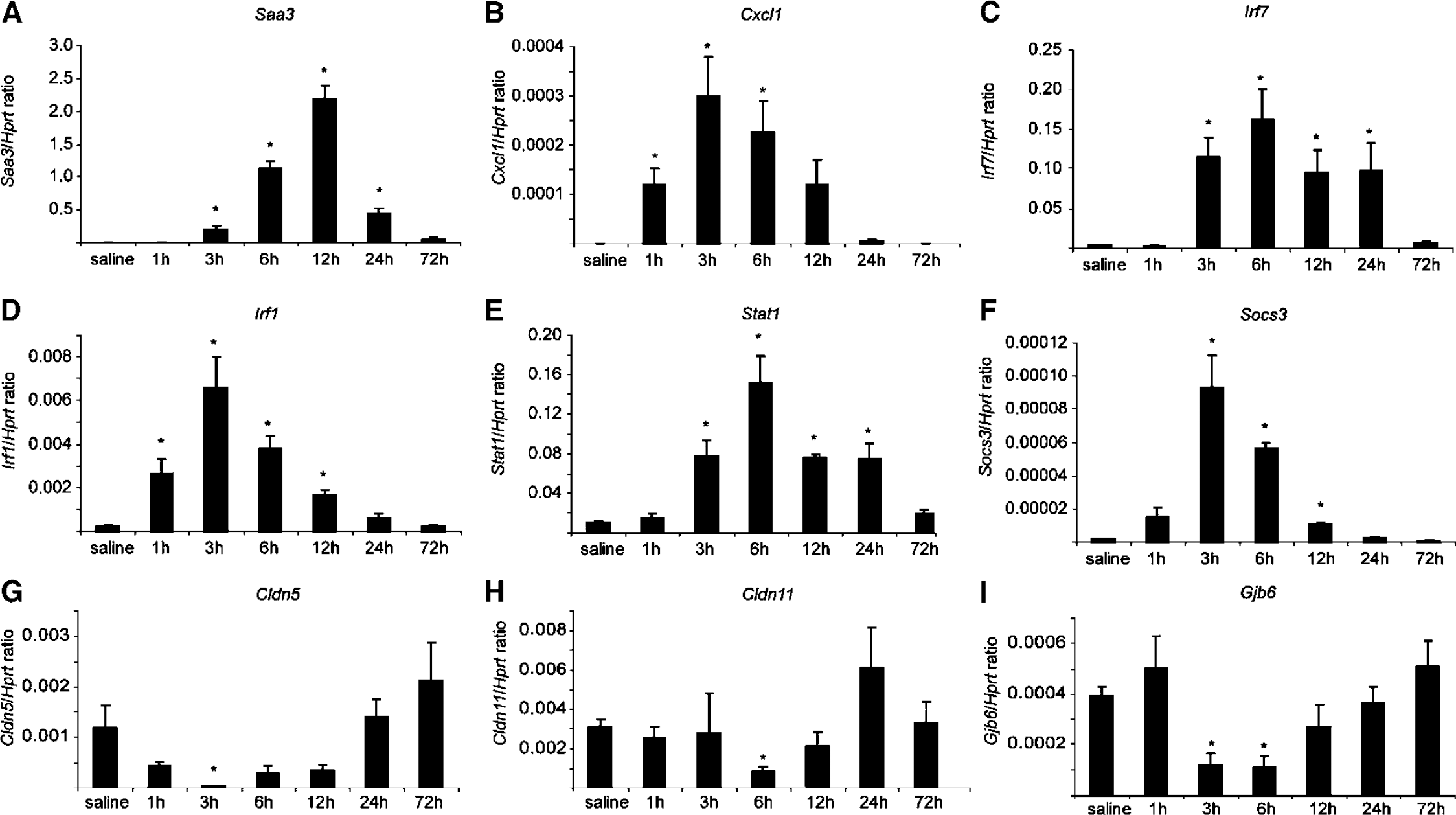
Analysis, by qRT-PCR, of the gene expression kinetic profile of selected genes. Confirming the array results, the expression of
Cerebrospinal Fluid Concentration of Cytokines
A significant correlation was found between the CP mRNA and the protein CSF concentration of the chemokine CCL2 (Figure 3A) and the cytokine IL-6 (Figure 3B). The CSF levels of CCL2 (n = 4) and IL-6 (
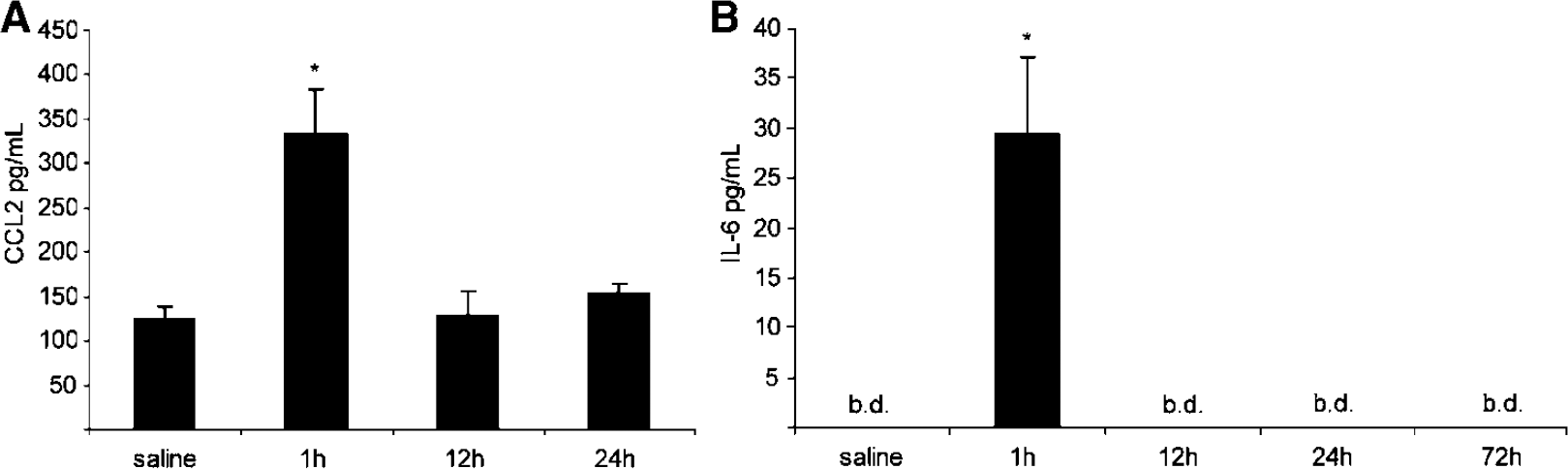
The CSF levels of CCL2 and IL-6 after peripheral injection of LPS. The concentration of CSF CCL2 (
Response of Primary Cultures of Rat Choroid Plexus Epithelial Cells to Various Stimuli
To study whether the observed response
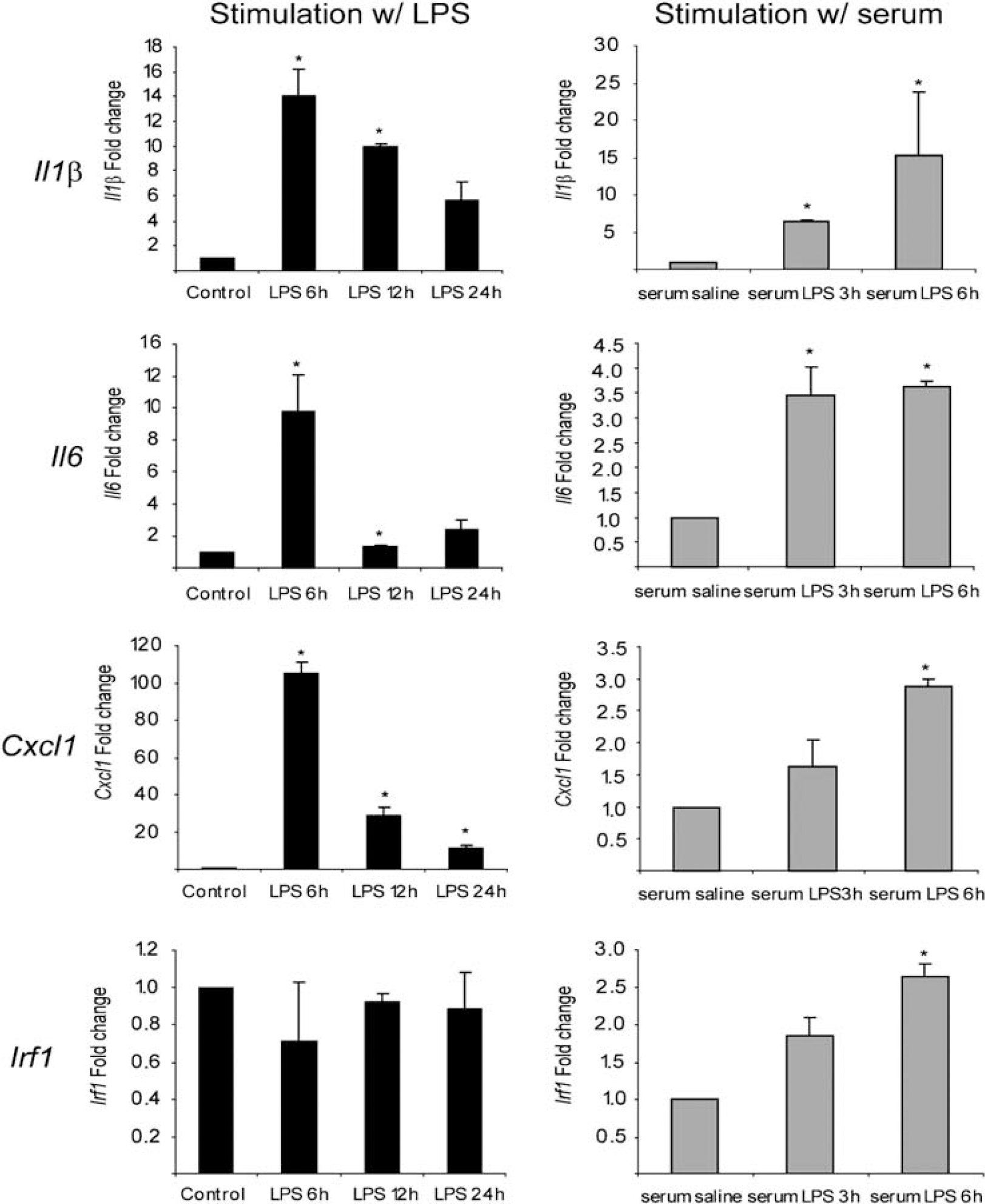
Analysis, by qRT-PCR, of the gene expression profile from rat CP epithelial cells. Primary cultures of rat CP epithelial cells were exposed to LPS alone for 6, 12, and 24 h; or for 6 h to serum obtained from rats 3 or 6 h after injection with LPS. Although both LPS and serum from LPS-injected rats induced the expression of
Characterization of the Choroid Plexus Acute Response
Degradation of the extracellular matrix is achieved through the action of proteases and the role of matrix metalloproteinases has been extensively investigated in this context (Flannery, 2006). We observed an up-regulation in the expression of genes encoding for proteases such as collagenase 13 (
Taken together, these observations suggest that the barrier function of the BCSFB may be transiently compromised.
Interestingly, the CP is not only able to activate these pathways but also to synthesize molecules responsible for the induction of inhibitory mechanisms, as illustrated by the results for proteins belonging to the SOCS family. In our array, the expression of genes encoding for the suppressors of cytokine signaling 2 and 3 (
Discussion
This study reveals that peripheral inflammatory stimuli can elicit an acute-phase response in the CP. As part of the BBBs, and because it determines the composition of the CSF, changes in the CP transcriptome can potentially play an important role in mediating the brain's response to inflammation. Notably, the kinetics of the CP response proved to be similar to that of the liver (Ceciliani et al, 2002; Gabay and Kushner, 1999) in terms of response onset and shut-off; and in some of the acute-phase proteins secreted (e.g., SAA, LCN2, IL-6, IL-1β) into the CSF. In addition, the liver and the CP acute responses share common signaling pathways, for example, NF-kB, JAK/STAT, and MAPK. Despite the above commonalities, the CP epithelium expresses a number of distinct genes after being challenged with an acute inflammatory stimulus; these genes are implicated in its barrier function although, at least within the time frame of the present experiments, we did not observe any gross CP morphologic changes.
Changes in the functionality and integrity of the BBB are relevant to the pathogenesis of a variety of diseases of the CNS (e.g., HIV-1 encephalitis, Alzheimer disease, MS, Parkinson disease, ischemia, and tumors) (Persidsky et al, 2006). For example, proinflammatory substances produced by microglia and specific disease-associated proteins or cells (β-amy-loid in Alzheimer disease and reactive T cells in MS) often mediate BBB dysfunction. Although it is not known exactly how the vascular endothelial cells that form the BBB respond to an inflammatory stimulus, it is plausible that the CP response shares many of the mechanisms already described for the BBB. A number of studies support the idea that changes in endothelial cell tight junctions and cytoskeletal organization facilitate leukocyte migration, through the interaction of chemokines and adhesion molecules, such as E-selectin and vascular cell adhesion molecule 1 with leukocytes (Engelhardt, 2008). The herein presented data for the CP show decreased expression of tight junction proteins and increased expression of proteins that facilitate migration. These changes are likely to underlie the preferential migration of T lymphocyte across the BCSFB that has been observed in animal models of MS (Brown and Sawchenko, 2007) and suggest a role for the CP in cell entry into the CSF and ultimately, into the brain parenchyma. Together with studies implying particular CP proteins in diseases such as Alzheimer disease (Sousa et al, 2007), this study warrants further studies to examine the CP transcriptome in neurologic and psychiatric diseases.
Apart from being a potential site of cell entry into the brain, the CP is also an active site of protein synthesis. Several inflammatory markers have been described in the CSF of individuals with brain pathology (Andreasen and Blennow, 2005; Giovannoni, 2006). The precise origin of these molecules is still equivocal, but CP epithelial cells might be important for their trafficking/entry into the CSF. The nature of the proteins secreted by the CP may change in response to disease or specific stimuli, as shown here for CCL2 and IL-6, and in previous studies for various cytokines, carrier proteins, and iron-related proteins (Marques et al, 2007, 2008; Hughes et al, 2002; Thibeault et al, 2001). In fact, several inflammatory chemokines (CCL2, CCL5, CXCL10, CXCL12, and CXCL13) whose gene expression is altered in the CP after peripheral LPS injection are detectable in the CSF of MS subjects (Trebst and Ransohoff, 2001) and CCL2 mRNA levels in the CP are increased after a single systemic bolus of LPS (Thibeault et al, 2001). The CP is known to secrete proinflammatory and antiinflammatory molecules, proteins that promote and inhibit extracellular matrix remodeling, and proteins to which neuroprotective and/or toxic properties have been ascribed to. Their beneficial or detrimental effects may depend on other factors such as age and the presence of pathogenic and/or disease conditions, which should next be investigated.
Proposed Model of the Choroid Plexus Response to Peripheral Inflammation
Irrespective of the final determinants of the balance between benefit and detriment, the present data provide interesting insight into the signaling transduction pathways that ultimately lead to the observed gene expression profiles in response to a pathogenic stimulus (Figure 5). Previous work showed that serum concentrations of cytokines (e.g., IL-1β and IL-6) are increased after peripheral administration of LPS (Ramadori and Christ, 1999). Receptors for these cytokines are present in CP epithelial cells (Chodobski and Szmydynger-Chodobska, 2001) and were detected by our array analysis under basal conditions. Thus, LPS- and cytokine-mediated signaling transduction pathways may be simultaneously engaged by peripheral inflammation. In fact, the cytokine-mediated pathways may play a general role in the CP response as the secretome of LPS-treated cultured CP epithelial cells displays a much reduced number of altered proteins (Thouvenot et al, 2006). This interpretation is consistent with that of the present study in which the up-regulation of
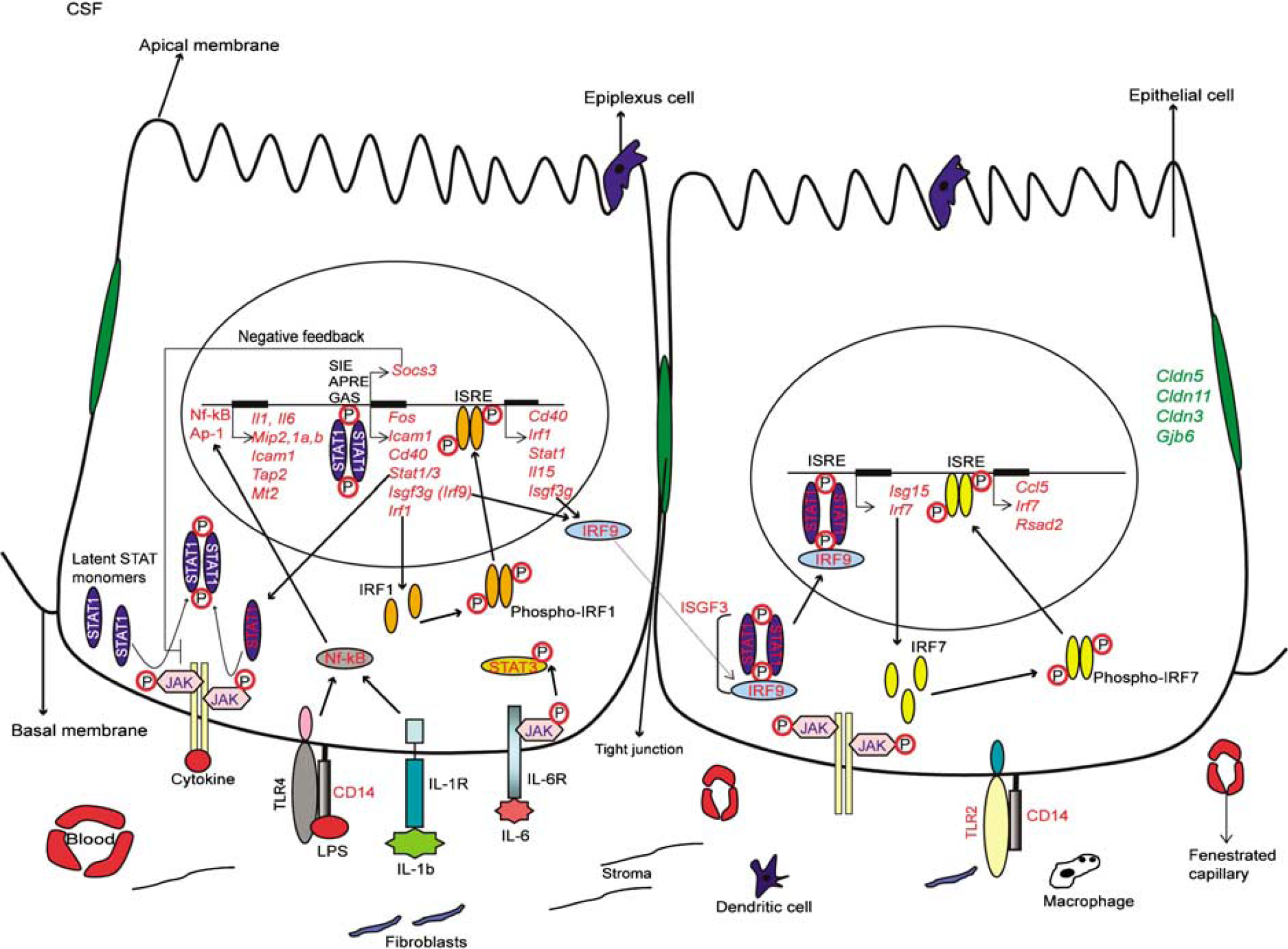
Suggested pathways of the CP epithelial cells response to peripheral inflammation. The genes whose expression was, at one or more time points, down-regulated are represented in green, and whose expression was up-regulated at one or more time points are represented in red.
With respect to LPS signaling, we confirmed that the gene encoding for the Toll-like receptor 4 (
Activation of TLRs and of receptors for IL-lβ, IL-6, and TNF results in the induction of several transcription factors, including interferon regulatory factor 3 (IRF3), activator protein-1, and NF-kB (Colonna, 2007; Honda and Taniguchi, 2006). Our analysis shows that several genes encoding for proteins belonging to the NF-kB, MAPK, STAT-JAK, and IRFs signaling pathways are induced in the CP during the acute-phase response. All of these pathways, together with others such as those mediated by cAMP (Reyes-Irisarri et al, 2008), participate in the development and regulation of innate and adaptative immune responses (Rawlings et al, 2004), and the CP, like the BBB endothelium (Laflamme et al, 1999; Laflamme and Rivest, 1999), responds to peripheral LPS by activating some of these pathways. Termination of the CP response at 72 h may be, at least in part, a consequence of the negative feedback inhibition of STAT signaling by SOCS/CIS (Naka et al, 2005), as some of the genes encoding for proteins in this pathway are up-regulated after LPS administration. A similar mechanism has been described in the BBB (Lebel et al, 2000).
In summary, this study shows that the CP can mount an acute-phase response. This response includes the release of proinflammatory and antiinflammatory immune modulators into the CSF, and a relaxation of the CP barrier properties. The evidence provided for a potentially important role of the CP in the cross talk between the immune system and the CNS warrants further studies that will add to our understanding on how homeostasis is maintained in the brain under normal and pathologic states.
Footnotes
Acknowledgements
This work was supported by grant POCTI/SAU-NEU/56618/2004 from the Portuguese Foundation for Science and Technology (FCT)/FEDER; and from a grant from the DANA foundation; Marques F, Falcao AM, and Rodrigues AJ are recipients of fellowships from FCT/FEDER.