
Abstract
Human islet transplantation is a viable treatment option for type 1 diabetes mellitus (T1DM). However, pancreatic islet inflammation after transplantation induced by innate immune responses is likely to hinder graft function. This is mediated by incompatibility between islets and the blood interface, known as instant blood-mediated inflammatory reaction (IBMIR). Herein we hypothesized that portal venous administration of islet cells with human recombinant antithrombin (ATryn
Keywords
Introduction
Type 1 diabetes mellitus (T1DM) is a chronic, progressive autoimmune disorder characterized by destruction of insulin-producing β-cells. The morbidity and mortality associated with T1DM are mainly related to the development of long-term microvascular and macrovascular complications 1 . Insulin therapy has allowed T1DM to become a chronically manageable condition 2 ; however, concerted efforts globally have emerged to ameliorate the symptoms and complications of T1DM via restoration of β-cell mass function through islet transplantation3–5. With the constant global rise in the incidence of T1DM and a limited supply of donor pancreata from scarce organ donors, continuous efforts are being pursued to improve islet isolation procedures and prevent islet loss, especially in the immediate posttransplant period through a series of strategies 6 .
Intrahepatic islet infusion via the portal vein, also referred to as intraportal islet allotransplant, accounts for virtually all clinical islet transplants conducted world-wide 7 . However, a significant amount of intraportal islet mass is lost immediately posttransplant because of innate immune responses, thus triggering pancreatic islet inflammation, which is likely to hinder graft function. This process is mediated primarily by incompatibility between islets and the blood interface, a phenomenon known as instant blood-mediated inflammatory reaction (IBMIR) 8 . IBMIR has been linked to a variety of inflammatory mediators, such as binding of antibody and complement, rapid activation of the coagulation cascade, and innate cellular infiltration, all of which contribute to islet destruction and nonengraftment8,9. As a result, after transplantation, the final islet cell survival rate is approximately 20% to 40% compared to that of a healthy nondiabetic subject 10 . Furthermore, inflammation-induced venous thrombosis may develop. Inflammatory thrombosis is driven by tissue factor (TF) and thus manifests itself clinically with formation of thrombi with entrapped islets11,12, resulting in massive thrombosis in the portal vein and a decrease in the engraftment of islets. Despite continued procedural improvements in isolation techniques 13 and strategies devised to selectively block the immune responses against the graft, insulin independence among autologous islet transplantation patients is rather poor, ranging at about 40% 14 . Thrombin, a serine proteinase generated from the zymogen prothrombin, is considered to be the key enzyme in the clotting reaction and therefore plays an important role in the IBMIR reaction. Indeed, studies have shown that inhibition of thrombin abrogates the IBMIR triggered by isolated human islets and reduces the adverse effects of IBMIR in clinical islet transplantation 15 and therefore poses as an attractive therapy option for T1DM.
Antithrombin (AT) is a non-vitamin K-dependent protease that inhibits coagulation by lysing thrombin and Factor Xa (FXa). AT activity is also markedly potentiated by heparin and is associated with an anti-inflammatory and antiprotease activity 16 . It also inhibits neutrophil activation 17 . ATryn® (human recombinant antithrombin) is an anticoagulant manufactured by GTC Biotherapeutics (Framingham, MA, USA). It is a milk-derived biological product produced by genetically engineered (GE) goats to produce human AT 18 . In view of ATryn® properties, we hypothesized that portal venous administration of islet cells with ATryn®, which plays a central role in the physiological regulation of coagulation and exerts indirect anti-inflammatory activities, may offset coagulation abnormalities such as disseminated intravascular coagulation (DIC) and IBMIR observed during pancreatic islet engraftment.
Materials and Methods
Animals and Experimental Protocol
All animal procedures were performed in the Department of Experimental Research of the Lille University, Lille, France, in accordance with French regulations for animal experiments. A total of n = 28 healthy female piglets (3–4 months) weighing 26 ± 2 kg (Pannier's breeding, Wylder, France) were used for the study. Pancreases were obtained from seven donors, and three groups were constituted for the transplantation with digested pancreatic tissue (0.15 ml/kg): control using NaCl 0.9% (n = 7), heparin (25 UI/kg) (n = 7) (Sanofi, Gentilly, France), and ATryn® (500 UI/kg) (n = 7). ATryn® (a gift from GTC Biotherapeutics) powder (7 UI/mg) was dissolved in solution distilled water.
Pancreatectomy, Pancreatic Tissue Isolation, and Graft
All surgical procedures were performed under gen eral anesthesia, induced following administration of buprenorphine at 0.03 mg/mg by subcutaneous injection (nalbuphine; Merck, Paris, France), an intramuscular injection of 0.3 mg/kg of midazolam (Hypnovel®; Roche, Neuillysur-Seine, France), and 5 mg/kg of ketamine (Ketamine Virbac®; Virbac, Carros, France). Tracheal intubation was performed subsequently with 0.2 ml/kg/h of intravenous propofol (Diprivan® 1%; AstraZeneca, Rueil-Malmaison, France). In all cases, partial pancreatectomy was performed with cautious dissection to maintain normal vascularization and to minimize warm ischemia until final excision of the tail of the pancreas. The explanted pancreas was immediately cooled in Hank's solution (Sigma-Aldrich Chimie, Saint-Quentin-Fallavier, France) at 4°C.
Within 30 min of cold ischemia, the pancreas was distended by infusion of 250 mg of collagenase and neutral protease (SERVA Electrophoresis Gmbh, COGER SAS, Paris, France) using an 18-gauge catheter (Optiva®; Ethicon, Pomezia, Italy) inserted in the pancreatic duct. Pancreatic tissue was isolated using state-of-the-art techniques as previously described 19 . The digested pancreatic tissue was resuspended in Connaught Medical Research Laboratories medium (CMRL) (Thermo Fisher Scientific, Illkirch, France) and supplemented with porcine serum, respectively, from each recipient and transported at room temperature to the operating theater.
Blood samples were drawn by a catheter placed in the jugular vein. After each sample, the catheter was flushed with saline buffer. Pancreatic digested tissue was placed in a 60-ml syringe with ATryn® or heparin and was injected at 2 ml/min. Cell injection was performed on anesthetized animals via a catheter placed directly in the portal vein. All hemodynamic parameters were monitored during injection. After injection, animals were woken and a catheter was maintained for blood sampling.
The same conditions were used for control experiments. All studies were performed in agreement with French regulations and approved by the institutional ethics committees of Lille University.
Blood Sampling and Biological Measurements
Blood samples were collected before injection (T0) and after 30 min, 1 h, 2 h, 3 h, 6 h, 12 h, 24 h and were collected from the arterial catheter in sterile tubes, and plasma was immediately separated after centrifugation at 4,815 × g for 10 min. Leukocyte, hemoglobin, and platelet counts were obtained on ethylenediaminetetraacetic acid (EDTA) (Sigma-Aldrich Chimie) anticoagulated blood. For coagulation assays, blood was collected in tubes containing 3.8% sodium citrate (Sigma-Aldrich Chimie). Prothrombin time (PT) and fibrinogen levels were rapidly measured by standard procedures. Immunoassay methods were used to determine quantitative thrombin–AT (TAT) complexes (Enzygnost TAT micro; Siemens, Munich, Germany). Fibrin monomer (Liatest FM; Diagnostica Stago, Asnieres, France) and D-dimer (Innovance DDI; Siemens, Marburg, Germany) were performed by immu-noturbidimetric assay. Quantitative AT activity was determined by chromogenic assay (Berichrom AT; Siemens). Inflammation [tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6)] parameters were measured with an ELISA method (Porcine Quantikine ELISA; R&D Systems, Lille, France).
Toxicity Test
ATryn toxicity was performed in human islets (n = 3 preparations). Human pancreata were isolated from adult, brain-deceased donors in agreement with our institu tional ethical committee, and islets were isolated according to a modified version as described previously 20 . Purification was achieved with Biocolle (Biochrom Ltd., Cambridge, UK) continuous density gradients using a COBE 2991 cell separator (Terumo BCT, Lakewood, CO, USA) 21 .
Purified islets were cultured with or without ATryn® (5 UI/ml = 0.7 mg/ml) for 24 h. We reproduced our clinical protocol, and we used the culture medium CMRL with 1 g/L of glucose (Thermo Fisher Scientific, Courtaboeuf, France) with penicillin/streptomycin and human serum albumin (HSA) 0.625% (Vialebex, Les Ulis, France).
Viability (blue trypan test), percentage of recovery, percentage of insulin secretion (20 mM glucose), and apoptosis (Cell Death Detection ELISA PLUS; Roche, Boulogne-Billancourt, France) were evaluated. Each condition was performed six times, and insulin secretion was normalized to intracellular insulin content.
Statistical Analysis
All data were expressed as mean ± standard error of the mean (SEM). Statistical analyses were performed using analysis of variance (ANOVA) with Tukey's post hoc test for repeated measures during the first 24 h of the experience. Analyses were performed using StatView 6® (SAS, Cary, NC, USA), and values of p < 0.05 were considered statistically significant.
Results
Heparin is a heterogeneous group of straight-chain anionic mucopolysaccharides called glycosaminoglycans that have anticoagulant properties. Previous studies have shown that heparin in combination with AT inhibits thrombosis by inactivating activated FX and inhibiting the conversion of prothrombin to thrombin 22 . However, studies comparing the AT efficacy of heparin and recombinant ATryn® have not been carried out. The purpose of this investigation was to evaluate the effect of heparin versus ATryn® on the mortality rate and DIC following intraportal islet administration in piglets. Three groups were constituted for digested pancreatic tissue transplantation (0.15 ml/kg): control, NaCl 0.9% (n = 7); gold standard, heparin (25 UI/kg) (n = 7); and human recombinant ATryn® (500 UI/kg) (n = 7). In both the control and heparin groups, one animal died following intraportal islet administration at 25 and 150 min, respectively, due to a massive portal thrombosis, while no deaths occurred in the ATryn® group (Fig. 1A). The autopsy results revealed a massive portal thrombosis (Fig. 1B) that was associated with cardiogenic shock manifested by sinus tachycardia, hypotension, followed by ventricular tachycardia with ventricular fibrillation. Postoperative complications, such as hemorrhagic complication as hemoperitoneum, were absent in the animals. A low serum albumin concentration was observed in the first hours of the experiment, increasing after 2 h and stabilizing for the rest of the experiment in the heparin and ATryn® groups. However, there were no differences in hepatic response to islet injection between groups (Fig. 1C). These data suggest that intraportal islet administration could be associated with a low hemodilution, increasing at 2 h. In the ATryn® group, a rapid increase in AT activity was observed after 30 min (200% activity), which escalated to 800% activity within 3 h. After 5 h, AT activity was maintained over 200% until the end of the experiment (Fig. 1D).

ATryn® administration in vivo prevents death and DIC. (A) Kaplan–Meier survival curves of piglets treated with NaCl 0.9% (n = 7), heparin (25 UI/kg) (n = 7), or ATryn® (500 UI/kg) (n = 7) from time 0 to 24 h. (B) Photograph of livers after sacrifice. (C) Serum albumin (g/L) from time 0 to 24 h. (D) Plot of plasma antithrombin activity expressed as percentage of activity. Data are expressed as means ± SEM. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001, two-way ANOVA and Tukey's post hoc test. DIC, disseminated intravascular coagulation; SEM, standard error of the mean; ANOVA, analysis of variance.
ATryn® Functions as a Potent Anticoagulant
We next investigated the effect of ATryn® versus NaCl (placebo) and heparin (reference treatment) versus NaCl on coagulation activity by using markers of coagulation, that is, TAT complex, fibrin monomer, platelets, prothrombin, and D-dimer. We have observed an initial peak in TAT activity (2 h) in the ATryn® versus NaCl group compared to the heparin versus NaCl group. In fact, TAT activity was significantly higher in the ATryn® versus NaCl group compared to the heparin versus NaCl group (Fig. 2A). A rapid and significant increase in fibrin monomers was observed in the NaCl group compared to the heparin and ATryn® groups (Fig. 2B).

Levels of markers of coagulation. (A) TAT (μg/L), (B) fibrin monomer (μg/ml), (C) platelets (%), (D) prothrombin (%), and (E) D-dimer (mg/L) at baseline up to 24 h in relation to treatment with NaCl, heparin, or ATryn® in the piglet. Data are expressed as means ± SEM. ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001, two-way ANOVA and Tukey's post hoc test. ANOVA, analysis of variance; TAT, thrombin–antithrombin; SEM, standard error of the mean.
There was no significant difference in platelet count and PT between groups at 24 h (Fig. 2C and D). There was no significant difference in platelet count between the three groups during the course of the experiment (Fig. 2C). However, there was a tendency of thrombocytopenia in the first 5 h of the experiment (Fig. 2D). D- Dimers reflecting activation of fibrinolysis were increased rapidly after islet administration in the NaCl group (Fig. 2E), achieving a maximum level at 3 h. However, this effect was lower in the ATryn® and heparin groups. ATryn® administration was associated with a significant decrease in coagulation parameters such as D-dimers and fibrin monomers compared to control.
ATryn® Does Not Modulate TNF-α and IL-6
Previous studies have shown that administration of ATryn® in human endotoxemia suppressed inflammatory markers such as TNF-α and IL-6 23 . In this study, we did not observe any significant change in TNF-α or IL-6 levels after ATryn® administration (Fig. 3A and B). In parallel, there was no significant change in leukocyte count and fibrinogen levels across the three groups at 24 h (Fig. 3C and D).
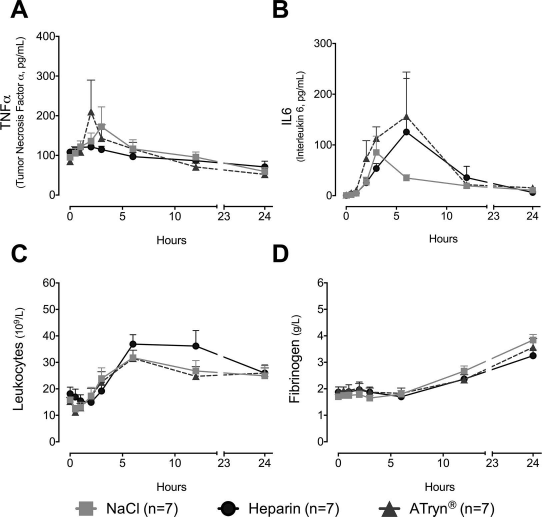
Levels of markers of inflammation. (A) TNF-α (pg/ml), (B) IL-6 (pg/ml), (C) leukocyte counts (109/L), and (D) fibrinogen (g/L) at baseline up to 24 h in relation to treatment with NaCl, heparin, or ATryn® in the piglet. Data are expressed as means ± SEM. Nonsignificant (ns), two-way ANOVA and Tukey's post hoc test. SEM, standard error of the mean; ANOVA, analysis of variance; TNF-α, tumor necrosis factor-α; IL-6, interleukin-6.
ATryn® Does Not Seem Toxic for Islets
Here we show for the first time that ATryn® treatment in human islet cultures in vitro had no effect on islet viability, recovery, or insulin secretion (Fig. 4). Thus, the mechanisms of these effects remain to be evaluated. More studies are needed to clarify if anticoagulant or anti-inflammatory effects of recombinant human AT (ATryn®) are implicated.

Characterization of the toxic effects of ATryn® in human islet cultures after 24 h. (A) Cell viability determined by the trypan blue exclusion test (%). (B) Islet recovery by cell counting using dithizone staining (%). (C) Insulin secretion (percentage of content) after stimulation with 20 mM glucose. (D) Apoptosis (absorbance in nanometers) determined by Cell Death Detection ELISA PLUS in the presence or absence of ATryn® (5 UI/ml = 0.7 mg/ml) in human islet cultures (n = 3 individual donors in duplicate) for 24 h. Data are expressed as means ± SEM. Nonsignificant (ns), Student's t-test. SEM, standard error of the mean.
Discussion
AT is a natural anticoagulant produced by the liver that plays a fundamental role in maintaining coagulation homeostasis by inhibiting thrombin and FXa and, to a lesser extent, FIXa and FXIa24,25. In addition to its anticoagulant function, it has also been shown to have potent anti-inflammatory and antiaggregant properties, mediated through the release of prostacyclins from endothelial cells26–28. The administration of AT seems to also be efficient to reverse the coagulation abnormalities in patients with signs of DIC 29 . AT derived from human plasma is therefore an essential class of therapeutics used in the treatment of life-threatening conditions resulting from trauma, infections, or sepsis. Like all other plasma derivates, AT concentrates are prepared from pools of human plasma ranging from 10,000 to 50,000 donations, which are required for industrial processing. More over, because of the human origin of the starting material, the major risk associated to plasma products is the transmission of blood-borne infectious agents 30 . Another source of AT replacement is human recombinant AT (ATryn®). ATryn® is produced through transgenic technology, which results in a purified AT substitute that is collected from milk of GE goats rather than from pooled human blood 18 .
The present study demonstrates that administration of ATryn® restored AT plasma concentrations without deleterious effects and was associated with a significant decrease in DIC. Although AT supraphysiological levels were achieved in our piglet model, we have not observed deaths nor hemorrhagic complications, while in the control (NaCl) and heparin groups we saw the death of one animal per group by portal thrombosis. This death was secondary to portal thrombosis provoked by local coagulation activation. Thus, the anticoagulant effect of ATryn® seems more robust in our piglet model. In humans, AT has been studied in severe sepsis with or without DIC31,32. A recent meta-analysis found AT to be beneficial in terms of mortality to treat patients with septic DIC, without any notable side effects 33 . Similar findings were reported in severe pneumonia with DIC 34 . In contrast, studies by Allingstrup et al. 35 did not find any beneficial effect of this treatment in patients with septic DIC or in patients with sepsis alone. In our model, animals with DIC who were treated with ATryn® had improved coagulation and viability.
Islet transplantation is a well-established therapeutic treatment for a subset of patients with T1DM. However, several pancreases are required to obtain a sufficient number of islets of Langerhans to establish insulin independence. A major drawback is the massive loss of islets, which occurs within minutes following the transplant. This has been attributed to coagulation abnormalities such as IBMIR that results in profound cellular degradation15,36, but also to activation of endothelial cells with the appearance of DIC and ischemia by a lack of vascularization 37 . IBMIR is also accompanied by activation of coagulation with decreased fibrinolysis as reflected in the elevation of TAT 38 . Inhibition of thrombin activation by melagatran has already shown efficacy in the reduction of IBMIR 15 . Thus, treatment with ATryn® may regulate coagulation activation and inflammation, via its anti-inflammatory properties and downregulation of neutrophils 39 , to improve the engraftment of islets. In our model, we observed a decrease in leukocyte counts in animals treated with ATryn®; however, we were unable to demonstrate any effect on the release of TNF-α and IL-6. Our findings suggest that ATryn® administration with islets directly in the portal vein may offset the recruitment and activation of leukocytes to the site of transplantation and thus improve engraftment. We deliberately chose a high dose (500 mg/kg) of ATryn® to be administered in animals in order to obtain a more effective and adequate anti-inflammatory effect. A striking correlation between AT activity and dosing (range, 50–150 IU/kg) has been previously demonstrated 40 . However, the anti-inflammatory effect requires doses >250 IU/kg 41 . The inflammatory response associated with islets has been long recognized as the primary cause of early damage to islets and graft loss after transplantation. Here we did not observe any significant change in TNF-α or IL-6 levels after ATryn® administration. However, we observed a gradual decrease, but nonsignificant, in leukocyte activation in animals treated with ATryn® compared to the heparin-treated group. These results are consistent with the inhibition of polynuclear chemotactism by ATryn® via the regulation of syndecan-4 receptor, which is important for the regulation of inflammatory effects of platelets 42 .
Conclusion
In conclusion, our results suggest that coadministration of ATryn® and pancreatic islets in the portal vein improves DIC. Although the anticoagulant effect is similar to that of heparin, ATryn® administration is associated with a better survival outcome for the animals. These studies also suggest that very high doses of ATryn® are not associated with side effects such as hemorrhagic complications. They also suggest that ATryn® may be a valuable therapeutic approach for DIC without risk for islets and subjects.
Footnotes
Acknowledgments
The authors acknowledge Dr. Yann Echelard, GTC Biotherapeutics, Framingham, MA, USA, for providing ATryn®. This work was supported by grants from LFB Biotechnology, France. The authors declare no conflicts of interest.