
Abstract
In previous work, we engineered functional cell sheets using bone marrow-derived mesenchymal stem cells (BM-MSCs) to promote islet graft survival. In the present study, we hypothesized that a cell sheet using dermal fibroblasts could be an alternative to MSCs, and then we aimed to evaluate the effects of this cell sheet on the functional viability of human islets. Fibroblast sheets were fabricated using temperature-responsive culture dishes. Human islets were seeded onto fibroblast sheets. The efficacy of the fibroblast sheets was evaluated by dividing islets into three groups: the islets-alone group, the coculture with fibroblasts group, and the islet culture on fibroblast sheet group. The ultrastructure of the islets cultured on each fibroblast sheet was examined by electron microscopy. The fibroblast sheet expression of fibronectin (as a component of the extracellular matrix) was quantified by Western blotting. After 3 days of culture, islet viabilities were 70.2 ± 9.8%, 87.4 ± 5.8%, and 88.6 ± 4.5%, and survival rates were 60.3 ± 6.8%, 65.3 ± 3.0%, and 75.8 ± 5.6%, respectively. Insulin secretions in response to high-glucose stimulation were 5.1 ± 1.6, 9.4 ± 3.8, and 23.5 ± 12.4 μIU/islet, and interleukin-6 (IL-6) secretions were 3.0 ± 0.7, 5.1 ± 1.2, and 7.3 ± 1.0 ng/day, respectively. Islets were found to incorporate into the fibroblast sheets while maintaining a three-dimensional structure and well-preserved extracellular matrix. The fibroblast sheets exhibited a higher expression of fibronectin compared to fibroblasts alone. In conclusion, human dermal fibroblast sheets fabricated by tissue-engineering techniques could provide an optimal substrate for human islets, as a source of cytokines and extracellular matrix.
Introduction
The outcomes of islet transplantation have improved significantly since the introduction of the Edmonton protocol in 2000 1 , but the long-term survival of the transplanted islet β cells has not been satisfactory 2 . There are several disadvantages of intraportal islet transplantation, including an instant blood-mediated inflammatory reaction 3 , the risk of exposure to high concentrations of immunosuppressive drugs 4 , and complications such as portal vein thrombosis and bleeding5,6. To overcome these disadvantages, researchers have attempted to transplant islets into extrahepatic sites including the kidney capsule, pancreas, spleen, omental pouch, testis, thymus, and subcutaneous locations7–15. Among them, the subcutaneous sites are ideal because of the minimal invasiveness involved. In addition, islets transplanted into a subcutaneous site can be removed easily and biopsied if needed.
However, the outcomes of islet transplantation into subcutaneous sites are still unsatisfactory because of poor vascularization, which restricts the blood supply to the graft. To create a vascularized subcutaneous site, several biomaterials combined with growth factors such as vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) have been developed9,10,14,16–19.
To overcome the poor vascularization in the subcutaneous site, we developed a method for performing islet transplantation using tissue-engineered cell sheet of islets and bone marrow-derived mesenchymal stem cells (BM-MSCs), and succeeded in the reversal of diabetes mellitus (DM) in a rodent model 20 . A cell sheet has a trophic effect, and it preserves the endogenous extracellular matrix (ECM) and adhesive factors21,22.
Before this technique can be applied in clinical settings, it is necessary to determine the effects of the cell sheets on human islets and to identify more convenient cell sources as a substrate. In the present study, we used human dermal fibroblasts for this purpose because fibroblasts can be harvested easily and are known to play important roles in promoting vascularization through the secretion of VEGF and FGF and to improve islet viability23–26. We fabricated cell sheets using human dermal fibroblasts, and we investigated the effects of the cell sheets on the viability and functioning of human islets.
Materials and Methods
Islet Isolation and Shipment
Human islets were isolated from consenting multiorgan deceased donors at the University of Alberta as previously described 27 with intent for clinical transplantation, and were only made available for research when the islet yield fell below that of the minimal mass required for clinical transplantation as previously described 28 . The human islets isolated at the University of Alberta were then shipped by air to Nagasaki University in accordance with the guidelines of the Ethics Committee at the University of Alberta. Consent for the research use of the islets was obtained from the donors' relatives. All experiments were conducted at Nagasaki University in accordance with the guidelines of the Ethics Committee at the Nagasaki University Graduate School of Biomedical Sciences.
Cell Culture
The shipped human islets were cultured at 37°C under 5% CO2 overnight in CMRL-1066 medium (Mediatech, Manassas, VA, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA, USA), 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen) before their use in this study. Human dermal fibroblasts (TIG-118 cells) were purchased from the Japanese Collection of Research Bioresources Cell Bank (Osaka, Japan). TIG-118 cells were cultured in minimum essential medium (MEM) (Invitrogen) supplemented with 10% FBS (Invitrogen), 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen) at 37°C in a humidified air atmosphere with 5% CO2.
Fabrication of Cocultured Fibroblast Sheets with Human Islets
We seeded fibroblasts at a density of 5.0 × 105 cells/dish onto a 35-mm- diameter temperature-responsive culture dish (UpCell®; CellSeed, Tokyo, Japan) with 2 ml of MEM. After the fibroblasts were cultured for 72 h, the MEM was removed and the human islets were seeded onto fibroblasts with 2 ml of CMRL-1066 medium. Following an additional cocultivation at 37°C in a humidified air atmosphere with 5% CO2 for 72 h, the human islets spontaneously detached together with the fibroblasts as a cell sheet by a transfer of the temperature-responsive culture dish to another incubator at 20°C for 1–2 h.
The islets were divided into three groups: the islets-alone group, the coculture with fibroblasts group (the islets-with-fibroblasts group), and the islet culture on fibroblast sheet group (the islets-on-fibroblast-sheet group). The culture conditions of each group are shown in Figure 1. In the islets-alone group, 500 human islets were cultured in a 35-mm dish with an ultralow-attachment surface (Corning Life Sciences, Corning, NY, USA) with 2 ml of CMRL-1066 medium. In the islets-with-fibroblasts group, 500 human islets were cocultured with preseeded fibroblasts in a 35-mm dish with an ultralow-attachment surface with 2 ml of CMRL-1066 medium. In the islets-on-fibroblast-sheet group, 500 human islets were seeded onto a 35-mm temperature-responsive culture dish with preseeded fibroblasts with 2 ml of CMRL-1066 medium. In the latter two groups, 5.0 × 105 fibroblasts were preseeded and cultured for 72 h before the cocultivation with human islets in MEM.

Illustrations of islet culture groups. (A) Islets-alone group. Human islets were cultured in a 35-mm dish with an ultralow-attachment surface. (B) Islets-with-fibroblasts group. Human islets were cocultured with fibroblasts in a 35-mm dish with an ultralow-attachment surface. (C) Islets-on-fibroblast-sheet group. Human islets were seeded onto a 35-mm temperature-responsive culture dish.
Immunohistochemistry
Immunohistochemistry by insulin staining was performed to confirm the presence of β cells on the harvested cell sheets. After detachment of the fibroblast sheet cultured with human islets was achieved by decreasing the temperature, each cell sheet was fixed with 4% paraformaldehyde (PFA; Wako Pure Chemical Industries Ltd., Osaka, Japan). Cell sheets were embedded in paraffin (Paraplast Plus; Leica Biosystems Inc., Richmond, IL, USA). Hematoxylin and eosin (H&E; Muto Pure Chemicals Co. Ltd., Tokyo, Japan) staining was performed according to the standard staining protocol. For immunological staining, the sections were deparaffinized with xylene and rehydrated with graded ethanol. After treatment with preheated antigen retrieval solution (Dako, Glostrup, Denmark), the slides were allowed to cool at room temperature (RT) for 40 min, and then were treated with peroxidase-blocking solution (Dako) at room temperature (RT) for 10 min. Mouse monoclonal anti-human insulin antibody (1:100 dilution; Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used as the primary antibody. After reaction with the primary antibody at RT for 120 min, the sections were treated with a horseradish peroxidase (HRP)-labeled polymer conjugated to goat anti-mouse immunoglobulins (Dako) at RT for 60 min. Then the sections were stained with 3,3-diaminobenzidine tetrahydrochloride (DAB) containing hydrogen peroxide (Dako). The results of the staining were observed under an inverted microscope (Nikon, Tokyo, Japan).
Islet Viability and Survival Assay
In each group, 3 days after the start of the islet culture, calcein-acetoxymethyl (AM) and propidium iodide (PI) (Cellstain Double Staining Kit; Dojindo, Kumamoto, Japan) were used to analyze the viability of the islets. The assay solution was prepared as previously described 20 . The percentage of viable cells in each islet was counted under a fluorescence microscope (Nikon). For viability assay, five individual experiments were performed for each group. Total viability was calculated by dividing the number of viable cells by the number of all cells assessed. Because it is well known that the islet mass decreases during culture 27 , we evaluated the islet survival rate by assessing the islet mass based on islet equivalents at baseline and on day 3 and day 7 after the cultures. For survival assays, six individual experiments were performed for each group.
Insulin Secretion Assay
Three days after the islets were cultured, we conducted a glucose-stimulated insulin secretion test for each group. Krebs solution was prepared by adding together 25 mM 2-[4-(2-hydroxyethyl)-1-piperazinyl]ethanesulfonic acid (HEPES), 115 mM NaCl, 24 mM NaHCO3, 5 mM KCl, 1 mM MgCl2 · 6H2O, 0.1% bovine serum albumin (BSA), and 2.5 mM CaCl2 · 2H2O (Wako Pure Chemical Industries). Islets were sequentially incubated in Krebs solution containing 2.8, 28, and then 2.8 mM glucose again; each incubation period was 1 h. The supernatant was collected after each incubation, and the amount of insulin secreted was measured by an enzyme-linked immunosorbent assay (ELISA) (ELISA Kit; Abcam, Cambridge, MA, USA) according to the manufacturer's instructions. The stimulation index was calculated by dividing the amount of insulin secreted under the high-glucose concentration by that under the initial low-glucose concentration. The data were obtained from six individual experiments.
Measurement of Cytokine Secretions
Three days after the start of the islet culture, the culture media were collected and frozen at −80°C until they were used for assays. We used an ELISA Kit (Abcam) to measure the secretions of the human cytokines transforming growth factor-β1 (TGF-β1) and interleukin-6 (IL-6) according to the manufacturer's instructions. Five individual experiments were performed for the TGF-β1 assay, and eight individual experiments were performed for the IL-6 assay.
Western Blot Analysis
Fibronectin (FN) is an ECM protein that is known to mediate a cellular interaction 29 . The expression of FN in fibroblast sheets was evaluated by Western blot. Fibroblasts were seeded at a density of 5.0 × 105 cells/dish onto 35-mm-diameter temperature-responsive culture dishes or 35-mm-diameter dishes with an ultralow-attachment surfaces with 2 ml of MEM. Following a 72-h culture period, the fibroblasts and fibroblast sheets were collected from the dishes and lysed with Triton-deoxycholate (DOC) lysis buffer (Sigma-Aldrich, St. Louis, MO, USA). The supernatant was collected and stored at −80°C until analysis after centrifugation. The measurement of protein was performed with a bicinchoninic acid assay (BCA) kit (Pierce, Rockford, IL, USA), and the protein was mixed with loading buffer, followed by incubation at 95°C for 5 min. The 8% acrylamide gels were used for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred onto Immobilon-P membranes (Merck Millipore, Bedford, MA, USA), followed by blocking with 5% skim milk (MEGMILK SNOW BRAND Co. Ltd., Tokyo, Japan) in TBST [10 mM Tris-HCl, pH 7.8 (Nacalai Tesque, Kyoto, Japan), 100 mM NaCl (Wako Pure Chemical Industries), 0.1% Tween 20 (Wako Pure Chemical Industries)] for 60 min at RT. The membranes were incubated with rabbit polyclonal anti-FN antibody (1:500 dilution; Abcam) at 4°C overnight, and then incubated with HRP-conjugated secondary antibody (1:2,500 dilution; GE Healthcare, Buckinghamshire, UK) for 90 min at RT. The protein was visualized using ECL Prime detection reagent (GE Healthcare). β-Actin was used as a loading control. The expression of FN was quantified using ImageJ software [National Institutes of Health (NIH), Bethesda, MD, USA]. The data were obtained from four individual experiments. A detailed description of the methods was previously described 30 .
Electron Microscopy Analysis
After the culture of the islets on fibroblast sheets for 3 days, each cell sheet was fixed and dehydrated as described 20 . For scanning electron microscopy (SEM), each dehydrated cell sheet was treated with t-butyl alcohol, followed by freeze drying. The surface of each cell sheet was coated with gold and observed under an electron microscope (JSM-6700F; JEOL, Tokyo, Japan). For transmission electron microscopy (TEM), the dehydrated cell sheets were embedded in Epon 812 (TAAB Laboratories Equipment Ltd., Berkshire, UK), and the sections were cut with an ultramicrotome (Ultracut S; Leica, Vienna, Austria). After being stained with uranyl acetate (Cerac Inc., Milwaukee, WI, USA) and lead nitrate (Wako Pure Chemical Industries), the cell sheets were observed under a transmission electron microscope (JSM-1210; JEOL).
Statistical Analysis
The statistical analyses were accomplished using GraphPad PRISM6 for Mac (GraphPad Software, San Diego, CA, USA). The data are expressed as means ± standard deviation (SD). All data were collected in at least four individual experiments. The data were analyzed using an analysis of variance (ANOVA) with Tukey's multiple comparison test when comparing more than two groups, and unpaired t-tests when comparing two groups. A value of p < 0.05 was considered significant.
Results
Characteristics of Cocultured Fibroblast Sheets with Human Islets
Upon their arrival, we confirmed the morphological integrity of the human islets shipped from the University of Alberta by performing dithizone (DTZ) staining (Fig. 2A). The fibroblasts (5.0 × 105 cells) seeded onto 35-mm temperature-responsive culture dishes proliferated and became confluent within 3 days (Fig. 2B). Three days after the start of the fibroblast culture, we seeded human islets on the fibroblasts (Fig. 2C). The morphological integrity of the human islets cultured on the fibroblasts was further confirmed at this stage using DTZ staining (Fig. 2D).

Characterization of human islets cultured on fibroblast sheets. (A) Shipped human islets were stained with DTZ for characterization. Scale bar: 100 μm. (B) Fibroblasts seeded onto a 35-mm temperature-responsive culture dish grew to confluence within 3 days. Scale bar: 500 μm. (C) The seeded human islets adhered to the fibroblast sheet. Scale bar: 500 μm. (D) Human islets on the fibroblast sheets were stained with DTZ for characterization. Scale bar: 200 μm. (E, F) Cocultured fibroblast sheets with human islets could be harvested without dropping the islets off by decreasing the temperature. Scale bars: 200 μm (E), 1 cm (F). (G) H&E staining of a detached cocultured fibroblast sheet with human islets. Scale bar: 200 μm. (H) β Cells of islets cultured on the fibroblast sheets were stained with insulin. Scale bar: 200 μm.
The human islets adhered to the fibroblast sheet within 3 days, and each fibroblast sheet was detached without the islets dropping off when the temperature was decreased to 20°C (Fig. 2E, F). A histological examination revealed the presence of intact islets attached to each fibroblast sheet (Fig. 2G) as well as insulin-positive cells within the islets (Fig. 2H).
Effects of Coculturing with Fibroblasts or Fibroblast Sheets on the Survival and Viability of Human Islets
To determine whether coculturing with fibroblasts or a fibroblast sheet improves islet viability, we performed a viability assay after 3 days of culture. Figure 3A shows the results of the calcein-AM and PI double staining after 3 days of cultivation. In the islets-alone group, the majority of cells located in the central area of the islets were dead. Smaller numbers of islets were dead in the islets-with-fibroblasts group and in the islets-on-fibroblast-sheet group. The islet viability values of the islets-alone, islets-with-fibroblasts, and islets-on-fibroblast-sheet groups were 70.2 ± 9.8%, 87.4 ± 5.8%, and 88.6 ± 4.5%, respectively, with significant differences (Fig. 3B).

The results of the islet viability and recovery rate assay. (A) Double staining by calcein-AM and PI was performed. Smaller numbers of dead cells were observed in the islets-with-fibroblasts group and the islets-on-fibroblast-sheet group compared to the islets-alone group. Scale bar: 100 μm. (B) The islet viability of both the islets-with-fibroblasts group (n = 5) and the islets-on-fibroblast-sheet group (n = 5) was significantly higher than that of the islets-alone group (n = 5). *p < 0.05, **p < 0.01. (C) The islet survival rates of both the islets-with-fibroblasts and islets-on-fibroblast-sheet groups were significantly higher than that of the islets-alone group at 3 and 7 days after the cultivation. *p < 0.05, **p < 0.01, ***p < 0.001; n = 6 for each group.
The islet survival rates are shown in Figure 3C. The islet survival rate of the islets-on-fibroblast-sheet group was significantly higher than that of the islets-alone group at both 3 and 7 days after the cultivation.
Promoted Insulin Secretion by Coculture with Fibroblast Sheet
To investigate the effect of coculturing with fibroblasts or a fibroblast sheet on islet function, we performed the insulin secretion test on day 3. As shown in Figure 4A, the insulin secretion by the islets-with-fibroblasts group was significantly higher than that of the islets-alone group during the high-glucose incubation. In the islets-on-fibroblast-sheet group, the insulin secretion was significantly promoted at each concentration of glucose compared to the islets-alone group. As a result, the stimulation index of the islets-on-fibroblast-sheet group was significantly higher than those of the other two groups (Fig. 4B).

The insulin secretion test results and the cytokine secretion values. (A) The secretion of insulin stimulated by each concentration of glucose solution in the islets-on-fibroblast-sheet group was significantly higher than that in the islets-alone group. *p < 0.05, **p < 0.01; n = 6. (B) The stimulation index of the islets-on-fibroblast-sheet group was significantly higher than those of the islets-with-fibroblasts and islets-alone groups. *p < 0.05. (C) The amount of TGF-β1 secreted in the islets-with-fibroblasts group was significantly higher than that of the islets-alone group. **p < 0.01. The islets-on-fibroblast sheet group exhibited higher TGF-β1 secretions (but not significantly) compared to the islets-alone group; n = 5. (D) The amounts of IL-6 secreted by the islets-with-fibroblasts and islets-on-fibroblast-sheet groups were significantly higher than that of the islets-alone group. **p < 0.01, ****p < 0.0001; n = 8. NS, not significant.
Promoted Cytokine Secretion by Coculture with Fibroblasts or Fibroblast Sheet
The cytokine secretion results are shown in Figure 4C and D. The islets-with-fibroblasts group exhibited significantly higher TGF-β1 secretions compared to the islets-alone group. The islets-on-fibroblast-sheet group showed higher TGF-β1 secretions than the islets-alone group, but the difference was not significant. The levels of IL-6 of both the islets-on-fibroblast sheet group and the islets-with-fibroblasts group were significantly higher than that of the islets-alone group.
Elevated Expression of FN in the Fibroblast Sheet
To investigate the production of FN in the fibroblast sheets, we analyzed the protein expression of FN. The results of the Western blot analysis showed a higher expression of FN in the fibroblast sheets (Fig. 5A and B), suggesting that the fabrication of the fibroblast sheet promotes the production of FN.
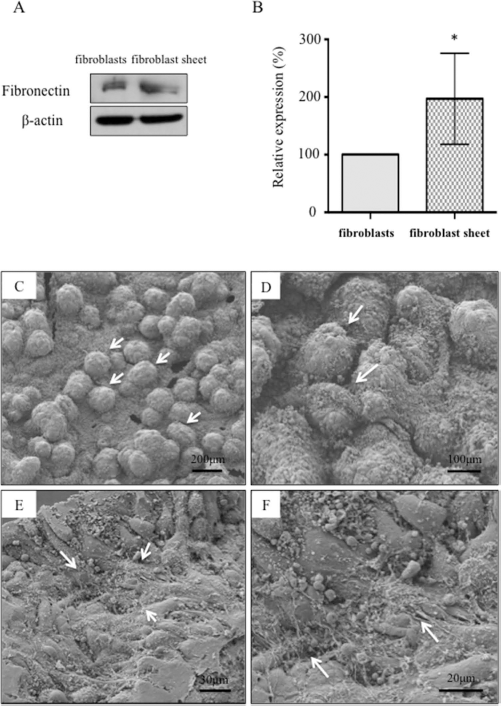
The elevated expression of FN in the fibroblast sheet and the preserved 3D structure of islets on the fibroblast sheet. (A) The Western blot analysis showed that the expression of FN in the fibroblast sheets was stronger than that in the fibroblasts. (B) The expression of FN in the fibroblast sheets was significantly higher than that in the fibroblasts. *p < 0.05; n = 4. (C, D) SEM demonstrated that the islets-on-fibroblast-sheets maintained a 3D structure. Scale bars: 200 μm (C), 100 μm (D). (E, F) The ECM was observed as a fibrous structure on the inferior side of the fibroblast sheets. Scale bars: 30 μm (E), 20 μm (F).
Ultrastructure of the Islets Embedded on the Fibroblast Sheet
We used SEM and TEM to investigate the morphology of human islets cultured on the fibroblast sheets. The SEM revealed that the islets embedded on a fibroblast sheet were in round or oval shape, maintaining their three-dimensional (3D) structure (Fig. 5C, D). ECM was observed on the inferior surface of the fibroblast sheets (Fig. 5E, F). We used TEM to observe the cell-to-cell connections, and we found that the cell-to-cell connections within the islets were maintained via tight junctions (Fig. 6A, B). Each fibroblast cell adhered via a tight junction, a gap junction, a desmosome, and an ECM (Fig. 6C, D). The ECM and adherens junctions were observed in the boundary between the islets and the TIG-118 dermal fibroblast cells (Fig. 6E, F). These results suggest that the islets adhered to the fibroblast sheets via the ECM and that cell–cell adhesion was maintained by adhesion factors.

TEM of cocultured fibroblast sheets with human islets. (A, B) The insulin granules were observed in the cytoplasm of β cells of the islets. Each β cell connected via a tight junction (white arrowheads). Scale bars: 1 μm (A, B). (C, D) Each fibroblast cell connected via a gap junction (black arrowhead), a desmosome (white arrows), and an ECM. Scale bars: 1 μm (C), 2 μm (D). (E, F) ECM and adherens junction (black arrows) were observed in the boundary between β cells and fibroblasts. Scale bars: 1 μm (E, F). TJ, tight junction; N, nucleus; F, fibroblast; GJ, gap junction; AJ, adherens junction.
Discussion
The results of the present study demonstrated that the cocultivation of islets with a fibroblast sheet can improve islet survival and function in vitro. Dermal fibroblasts could be an optimal cell source of cytokines, and dermal fibroblast sheets may have a protective effect on human islets as a 2D substrate with abundant ECM proteins.
Previous studies demonstrated that coculture with MSCs improved islet survival and function via cytoprotective effects by the secretion of several growth factors31–34. In the present study, we used dermal human fibroblasts as a cell source for the production of cytokines. The main reason for this choice of cells was that dermal fibroblasts can be easily harvested from the skin, and they expand rapidly ex vivo. Fibroblasts are also well known to play an important role in wound healing by the secretion of several cytokines 35 . Therefore, similarly to MSCs, fibroblasts seem to have protective effects that are conducive to islet survival and function. The present findings demonstrated elevated secretions of TGF-β and IL-6 in islets cocultured with fibroblasts. The role of TGF-β in the production of ECM has been revealed, and TGF-β1 gene therapy has been shown to have a protective effect on islet survival36–38.
IL-6 is one of the cytokines involved in inflammation, not only in the immune system but also in the nonimmune events in many types of tissue 39 . The serum IL-6 levels of individuals with DM are known to be elevated, contributing to their insulin resistance 40 . However, it was reported that IL-6 enhances insulin secretion by promoting glucagon-like peptide-1 (GLP-1) secretion 41 . Moreover, IL-6 secreted by MSCs has been known to be elevated when they are cocultured with damaged cells, and IL-6 is considered to have an antiapoptotic effect on islets31,42. Our present findings that coculture with fibroblasts improved islet viability and increased the insulin secretion by the secretion of cytokines are thus consistent with these previous studies, and these protective effects of fibroblasts may resemble those of MSCs.
In the field of cell transplantation, cell sheets created by tissue engineering have been developed in clinical research43,44. Compared to cell injection methods, cell sheets are known to demonstrate a higher degree of cell viability and engraftment at subcutaneous sites by vascularization via the secretion of angiogenic factors 45 . It was revealed in prior studies that islet sheets can be fabricated on a temperature-responsive dish using tissue-engineering techniques and that islet sheets transplanted into a subcutaneous site could survive and improve blood glucose levels33,46.
A temperature-responsive dish is coated with poly (N-isopropyl acrylamide), which can change from hydrophobic to hydrophilic at temperatures under 32°C. Cells cultured on such a dish can thus be harvested as a cell sheet without tripsinization and then maintain cell-to-cell junctions, preserving abundant ECM proteins that enhance the attachment of the cells to a subcutaneous site. The loss of native ECM proteins induced by enzymatic digestion during islet isolation has been shown to contribute to graft dysfunction 47 , and it was reported that the viability and function of islets were improved by entrapping islets using ECM proteins48,49.
In the present study, the elevated expression of FN in the fibroblast sheets was demonstrated by Western blotting, and the preserved cell-to-cell contact mediated by adherens-type junctions within islet cells was confirmed by TEM. FN is expressed in a variety of cells and mediates a cellular interaction with ECM proteins 29 . The main function of FN in this setting was to enhance insulin secretion, whereas other ECM components such as a collagen contribute to glucose metabolism 50 . It was reported that the cell-to-cell junctions play an important role in the maturation of β cells and insulin secretion 51 . In the present study, the islets cocultured with fibroblast sheets demonstrated higher viability, survival, and insulin secretion compared to islets cocultured with fibroblasts (not in sheets). The preserved ECM proteins and elevated expression of FN in fibroblast sheets may account for the additional observed benefits in the islets-on-fibroblast-sheet group.
In earlier studies, 3D scaffolds using biomaterials improved the survival and function of islets in vitro52,53. It was reported that 3D scaffolds reduced amyloid formation in human islet cultures 54 . In the present study, SEM revealed that human islets could be cultured while maintaining a 3D structure on a fibroblast sheet. These results suggest that fibroblast sheets could be an alternative substrate to maintain the natural structure of human islets and improve islet survival and functions.
In the field of regenerative medicine, islet transplantation could be an alternative therapeutic strategy to overcome the shortage of donors. It was reported that MSCs, embryonic stem cells (ESCs), and induced pluripotent stem cells (iPSCs) could differentiate into insulin-producing cells55–57, but the issue of safety regarding the tumorigenicity or cancer progression induced by those stem cells in humans remains controversial58–61. To introduce the use of those stem cells as a regenerative medicine technique in clinical settings, subcutaneous transplantation sites may be optimal, because the cells could be easy to access when a transplanted graft must be removed. It was reported that dermal fibroblasts transplanted into a subcutaneous site did not demonstrate overgrowth within the ECM even after 60 days posttransplantation 24 .
In conclusion, fibroblast sheets have beneficial effects on the in vitro survival and functioning of human islets as a 2D substrate, through not only the secretion of several cytokines but also the preservation of the ECM proteins. Although further studies using an in vivo model are required, this novel fibroblast sheet method could be a useful strategy in islet transplantation into subcutaneous sites.
Footnotes
Acknowledgments
We thank Mr. Takashi Suematsu for his technical assistance and advice on the ultrastructural analysis using the electron microscope. The authors declare no conflicts of interest.