
Abstract
Previously, we found that the intravenous administration of human adipose tissue-derived mesenchymal stem cells was a promising therapeutic option for autoimmune thyroiditis even when the cells were transplanted into a xenogeneic model without an immunosuppressant. Therefore, we explored the comparison between the therapeutic effects of syngeneic and allogeneic adipose tissue-derived stem cells on an experimental autoimmune thyroiditis mouse model. Experimental autoimmune thyroiditis was induced in C57BL/6 mice by immunization with porcine thyroglobulin. Adipose tissue-derived stem cells derived from C57BL/6 mice (syngeneic) or BALB/c mice (allogeneic) or saline as a vehicle control were administered intravenously four times weekly. Blood and tissue samples were collected 1 week after the last transplantation. Adipose tissue-derived stem cells from mice were able to differentiate into multiple lineages in vitro; however, mouse adipose tissue-derived stem cells did not have immunophenotypes identical to those from humans. Syngeneic and allogeneic administrations of adipose tissue-derived stem cells reduced thyroglobulin autoantibodies and the inflammatory immune response, protected against lymphocyte infiltration into the thyroid, and restored the Th1/Th2 balance without any adverse effects. However, different humoral immune responses were observed for infused cells from different stem cell sources. The strongest humoral immune response was induced by xenogeneic transplantation, followed by allogeneic and syngeneic administration, in that order. The stem cells were mostly found in the spleen, not the thyroid. This migration might be because the stem cells primarily function in systemic immune modulation, due to being given prior to disease induction. In this study, we confirmed that there were equal effects of adipose tissue-derived stem cells in treating autoimmune thyroiditis between syngeneic and allogeneic transplantations.
Keywords
Introduction
Hypothyroidism is a state in which the thyroid gland does not produce enough thyroid hormone to maintain the body's normal metabolism. Hypothyroidism is one of the most common endocrine diseases in humans, with the majority of cases resulting from iodine deficiency or autoimmune-mediated mechanisms (25). In autoimmune thyroiditis, antibodies against thyroid peroxidase or thyroglobulin cause the gradual destruction of follicles in the thyroid gland. Accordingly, the disease can be detected clinicopathologically by looking for these antibodies in the blood. Autoimmune thyroiditis is also characterized by the invasion of the thyroid tissue by leukocytes, mainly T-lymphocytes (4). Thyroiditis patients are predominantly between 30 and 50 years of age when diagnosed, and the prevalence of the disease increases with age, in both men and women.
Mesenchymal stem cells (MSCs) are nonhematopoietic, multipotent progenitor cells (33) capable of differentiating into mesenchymal lineages, including adipocytes, osteoblasts, chondroblasts, and myoblasts (14,18,32,34). Moreover, these cells can also acquire the phenotype of cells from unrelated germline lineages such as neural cells (2) and hepatocytes (39). Using the differentiation potential of MSCs, many studies have focused on the possibility of MSCs being used to repair damaged tissues or cells (19,31,48). MSCs were initially isolated from bone marrow but have also been found in many other adult tissues, including skeletal muscles, adipose tissues, peripheral blood, cord blood, and placenta (16,17,19, 23,24,35,44,49,53). While much research has focused on bone marrow-derived MSCs (BMMSCs), recent preclinical and clinical studies have focused on the use of adipose tissue-derived MSCs (ATMSCs) because of the less invasive and less painful procedures required to obtain the source tissue, the readily culture-expanding nature of the cells, and the possibility of harvesting abundant numbers of cells for transplantation (45,49). Furthermore, the numbers of resident MSCs in fat, along with proliferation and differentiation potentials, are less affected by donor age than are those found in bone marrow (7,27). For these reasons, it has been deemed acceptable to use ATMSCs as an alternative source of MSCs.
In addition to their differentiation capacity, MSCs have been highlighted as potent regulators of the immune response due to their immunomodulatory activity (30,46). Recent data have demonstrated a broad spectrum of immunoregulatory capabilities of MSCs, including the ability to a) inhibit the proliferation of T-cells stimulated with mitogens, allogeneic cells, or specific antigens (1,3,15,51), b) modulate the type 1 T helper cell (Th1)/Th2 balance by either inhibiting proinflammatory cytokines or promoting anti-inflammatory cytokines (1,29,50), c) increase regulatory T (Treg) cells (1,40), d) inhibit activated B-cell proliferation and immunoglobulin production (13,36), e) inhibit the maturation of monocytes and hematopoietic progenitor cells into dendritic cells (37,51), and f) inhibit the cytotoxicity of natural killer cells (41,42). ATMSCs have been characterized by a large body of in vitro research as having same or similar immunomodulatory effects as BMMSCs (8). The generation of severe graft-versus-host diseases in mice was prevented by the transplantation of ATMSCs (47). Regarding organ-specific autoimmune diseases, systematic infusion of human ATMSCs significantly ameliorated inflammatory bowel disease, collagen-induced arthritis, and autoimmune hearing loss in mice (20,21,52). These uses of ATMSC are part of the reason why an increasing number of researchers continue to turn their attention toward preclinical studies involving ATMSCs (9).
Previously, we reported the efficacy of human ATMSCs on a mouse autoimmune thyroiditis model. Although the injected cells were derived from different species, xenografted human ATMSCs showed the beneficial effects of decreased thyroglobulin autoantibody (TgAA) production and decreased lymphocyte infiltration in thyroid gland lesions (11). In this study, we address whether mouse ATMSCs could induce an equivalent immunomodulatory effect on autoimmune thyroditis to that exhibited by human ATMSCs. Although ATMSCs isolated from both humans and mice are known to have similar properties with respect to immunophenotypes and differentiation capacities, a few differences do in fact exist. Mouse ATMSCs are hard to expand in culture and, unlike human ATMSCs, show weak expression of the cluster of differentiation 73 (CD73) surface marker (22). MSCs derived from mice are immunophenotypically heterogeneous due to variations in tissue source, methods of isolation and culture, and strain differences (5,26). Thus, murine MSCs remain to be fully characterized (5). For further clinical application in humans, especially for autoimmune diseases, it is also important to be aware of efficacy and safety differences between autologous and allogeneic stem cell transplantation.
In this study, we investigated the immunophenotype characteristics of culture-expanded mouse ATMSCs and the immunomodulatory effect of mouse ATMSCs on autoimmune thyroiditis. Serological, immunological, and histological differences between syngeneic and allogeneic stem cells were also investigated.
Materials and Methods
Experimental Animals
Ten male C57BL/6 and BALB/c mice were purchased from ORIENT BIO (Sungnam, Korea) as donors of ATMSCs. Thirty C57BL/6 female mice (ORIENT BIO) were used as recipients in the in vivo studies. The recipients were 10 weeks of age at the time of use. This study was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the Samsung Biomedical Research Institute (SBRI). SBRI is an Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC International)-accredited facility and abides by the guidelines of the Institute of Laboratory Animal Resources (ILAR).
Isolation and Preparation of Mouse ATMSCs
Adipose tissues were obtained from axillary and inguinal subcutaneous fat from both C57BL/6 and BALB/c mice at 16–22 weeks of age. The subcutaneous adipose tissues were washed twice with Dulbecco's phosphate-buffered saline without calcium chloride and magnesium chloride (DPBS, Gibco BRL, Rockville, MD, USA) and were chopped. The tissues were dissociated by 0.1% collagenase I (Gibco BRL) under gentle agitation for 80 min at 37°C. The digested tissues were filtered through a 70-μm nylon sieve (BD Biosciences, San Jose, CA, USA) to remove cellular debris and centrifuged at 470 × g for 5 min to obtain a pellet. The pellet was treated with erythrocyte lysis buffer (BD Biosciences), resuspended DPBS, and recentrifuged at 470 × g for 5 min twice. The pellet was resuspended in modified Dulbecco's modified Eagle's medium (DMEM; WELGENE, Daegu, Korea) containing 5% adult bovine serum (ABS; Gibco BRL), 5 ng/ml basic fibroblast growth factor (bFGF; Invitrogen, Carlsbad, CA, USA), 5 ng/ml epithelial growth factor (EGF; Invitrogen), 0.1× ITS™+Premix (BD Biosciences), 0.2 mM ascorbic acid (Sigma-Aldrich, St. Louis, MO, USA), and 2 mM N-acetyl-cysteine (NAC; Sigma-Aldrich). The suspension was filtered through a 40-μm nylon sieve (BD Biosciences), and the collected cell fraction was cultured overnight at 37°C in 5% CO2 and 3% O2 in modified DMEM. After 24 h, nonadherent cells were removed by washing with PBS and the medium replaced. The cells were maintained for 4–5 days until confluent (passage 0) with medium changes every 2–3 days. When cells reached 80–90% confluence, they were detached using TrypLE express (Gibco BRL) and subculture expanded until passage 1 or 2.
Immunophenotype of Mouse ATMSCs
ATMSCs from each mouse strain were immunophenotyped by flow cytometric analysis. Fluorescein isothiocyanate (FITC)-conjugated hamster anti-mouse CD29 (clone: Ha2/5, 555005; BD Biosciences), phycoerythrin (PE)-conjugated rat anti-mouse CD31 (clone: MEC13.3, 553373; BD Biosciences), FITC-conjugated rat anti-mouse CD34 (clone: RAM34, 555733; BD Biosciences), FITC-conjugated rat anti-mouse CD44 (clone: IM7, 553133; BD Biosciences), FITC-conjugated rat anti-mouse CD45 (clone: 30-F11, 553079; BD Biosciences), PE-conjugated rat anti-mouse CD73 (clone: TY/23, 550741; BD Biosciences), FITC-conjugated mouse anti-rat/mouse CD90.1 (clone: HIS51, 554894; BD Biosciences), PE-conjugated rat anti-mouse CD 90.2 (clone: 53-2.1, 553006; BD Biosciences), PE-conjugated rat anti-mouse CD105/Endoglin (clone: 209701, FAB1320P; R&D Systems, Minneapolis, MN, USA), FITC-conjugated mouse anti-mouse H-2Kb [C57BL/6 major histocompatibility complex (MHC) class I, clone: AF6-88.5, 553569; BD Biosciences], FITC-conjugated mouse anti-mouse H-2Kd (BALB/c MHC class I, clone: SF1-1.1, 553565; BD Biosciences), PE-conjugated mouse anti-mouse I-Ab (C57BL/6 MHC class II, clone: AF6-120.1, 553552; BD Biosciences), and FITC-conjugated mouse anti-mouse I-Ad (BALB/c MHC class II, clone: AMS-32.1, 553547; BD Biosciences) were used for cell surface antigen detection. ATMSC immunophenotypes were analyzed using a FACSCalibur flow cytometer (BD Biosciences) using CELLQuest software version 5.1; BD Biosciences). Isotype control antibodies (550616, 553960, 554647, 554648, 554688, 554689, 556923, and 559532, BD Biosciences) were used to estimate the nonspecific binding of target primary antibodies to cell surface antigens.
For further immunophenotyping comparison studies at other passages (passage 3–5), StemXVivo™ MSC expansion medium (CCM004, R&D Systems) was used for the expansion of mouse ATMSCs.
Differentiation Capacity of Mouse ATMSCs
Adipogenesis
ATMSCs were exposed to adipogenic induction media (AIM) consisting of 10% fetal bovine serum (FBS; Gibco BRL)/DMEM, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX, I5879, Sigma-Aldrich), 1 μM dexamethasone (D4902, Sigma-Aldrich), 10 μg/ml insulin (4506-01, Millipore, Billerica, MA, USA), 100 μM indomethacin (I7378, Sigma-Aldrich), and antibiotic/ antimycotic (a.a, 15240112, Gibco BRL). After 6 days in AIM, ATMSCs were cultured in adipogenic maintenance medium (AMM) for 2 days. AMM consisted of 10% FBS and 10 μg/ml insulin in DMEM. ATMSCs were maintained at 37°C for 3 weeks, with the media changed every third day. Cells were stained using the Adipogenesis Assay Kit (ECM950, Chemicon, Temecula, CA, USA) according to the manufacturer's protocol. Other ATMSCs were grown in modified DMEM as a control. Oil Red O (Millipore) was utilized to visualize fat droplets.
Osteogenesis
ATMSCs were maintained in osteogenic induction medium [10% FBS, 100 μM dexamethasone, 50 mM ascorbic acid-2-phosphate (Sigma-Aldrich), 1 M glycerol-2-phosphate (Sigma-Aldrich), 1× a.a, DMEM-low glucose (WELGENE)] for up to 3 weeks, with fresh media added every third day. Cells were fixed with 4% paraformaldehyde (PFA; WAKO Pure Chemical Industries, Ltd., Osaka, Japan) and stored at 4°C until stained. As in the adipogenesis procedure, ATMSC cultures were grown in modified DMEM as a control. Alizarin red S (Sigma-Aldrich) was utilized to visualize the mineralized matrix.
Chondrogenesis
To promote chondrogenic differentiation, 106 ATMSCs were centrifuged in a 15-ml polypropylene tube (Corning Inc., Corning, NY, USA), and the pellets were cultured in chondrogenic medium (NH Chondro Diff. Media; Miltenyi, Hochschule Hannover, Germany). Three weeks later, pellets were harvested, and sections of paraffin-embedded pellets were stained with toluidine blue (Sigma-Aldrich) for identification. ATMSC cultures were grown in modified DMEM as a control. The undifferentiated status of ATMSCs was also confirmed by alcian blue staining (IHC World, Woodstock, MD, USA).
Neurogenesis
To promote neurogenic differentiation, ATMSCs were cultured in DMEM (high glucose; WELGENE) containing 10% FBS, 10 ng/ml FGF, and 10 ng/ml EGF for 24~48 h and then maintained in neurogenic medium [high-glucose DMEM supplemented with 10% FBS, 2% dimethyl sulfoxide (DMSO; Sigma-Aldrich), 200 μM butylated hydroxyanisole (BHA; B1253; Sigma-Aldrich), 5 μg/ml insulin, 0.5 mg isobutylmethylxanthine (IBMX, I7018; Sigma-Aldrich), 100 μM adenosine 3′5-cyclic monophosphate sodium salt monohydrate (cAMP; A6885; Sigma-Aldrich), 1 μM hydrocortisone (Sigma-Aldrich), and 1× a.a] for up to 2 weeks, with fresh media added every 2 days.
The culture medium from a chamber slide (LABTEK™, Thermo Fisher Scientific, Inc., Rochester, NY, USA) was aspirated and rinsed three times with DPBS without calcium and magnesium. Cells were fixed with 4% PFA for 1 h at 4°C and rinsed with DPBS three times. Then cells were permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 15–20 min at room temperature and rinsed with DPBS three times. Nonspecific binding was blocked with 5% normal goat serum (Abcam plc., Cambridge, UK)/DPBS for 1 h at 4°C. The slide was rinsed with DPBS three times. Anti-neuronal-specific enolase (NSE, ab64721; Abcam) or anti-neuron-specific class III β-tubulin (antibody name TuJ-1, MAB1637; Millipore) antibodies were subsequently added and the slide incubated overnight at 4°C. The slide was rinsed with DPBS three times and incubated with secondary antibodies (FITC-conjugated goat anti-rabbit IgG antibody or PE-conjugated goat anti-mouse IgG antibody, respectively; both from Abcam) for 1 h at 4°C. Then, the slide was rinsed with DPBS three times, stained with 4′,6-diamidino-2-phenylindole (DAPI; Thermo Fisher Scientific Inc.) for 2 min, and rinsed with DPBS three times. The slide was then coverslipped and observed under a Zeiss Axiovert 200 M fluorescence microscope (Carl Zeiss, Oberkochen, Germany).
In Vivo Study
Induction of Autoimmune Thyroiditis and Administration of Mesenchymal Stem Cells
For the induction of autoimmune thyroiditis, 100 μg of porcine thyroglobulin (pTg; Sigma-Aldrich) was emulsified in 100 μl Freund's complete adjuvant (CFA; Sigma-Aldrich) and was given subcutaneously to each mouse on day 0. The second subcutaneous injection was given on day 14 using the same amount of pTg in incomplete Freund's adjuvant (IFA; Sigma-Aldrich). Subsequently, 5 × 105 C57BL/6 ATMSCs (the syngeneic group; n = 10) or the same amount of BALB/c ATMSCs (the allogeneic group; n = 10) were administered intravenously on days 0, 7, 14, and 21. Control mice were treated identically but infused with 150 μl of saline instead of cells (n = 10). Two mice in each treated group were given fluorescently labeled ATMSCs using the conjugated red fluorochrome Cell Tracker CM-DiI (Invitrogen) for identification in histopathological sections. All mice were sacrificed on day 28. Blood was collected from the posterior vena cava and the sera stored at −70°C until use. The spleen and thyroid were removed from all mice. The splenic weight of all mice was measured soon after resection, and the spleen-to-body-weight ratios were calculated.
An additional animal study was conducted for flow cytometric analysis to determine the proportion of interleukin (IL)-17-producing CD4+ cells and Treg cells in the spleen (three or four mice in each treated group), for additional studies on humoral responses to infused cells (six mice in each treated group), and for confocal microscopic examination of transplanted CM-DiI-labeled mouse ATMSCs (three more mice in each treated group, to give a total of five mice in each treated group).
Thyroglobulin Autoantibody Levels in Sera and in Culture Supernatants of Splenocytes
Antibodies to pTg were detected by ELISA according to the method described in our previous study (11). Briefly, 100 μl of pTg diluted to 100 μg/ml in a coating buffer pH 9.6 (KOMA BIOTECH, Seoul, Korea) was used to coat flat-bottomed 96-well microtiter plates (Costar 3590; Corning) overnight at 4°C, followed by two washes with PBS–0.05% Tween 20 (PBST; KOMA BIOTECH). Blocking solution [PBS–1% bovine serum albumin (BSA; KOMA BIOTECH)] was applied for 2 h at 37°C and then washed again with PBST. The sera from individual mice were diluted 1:1,000 in blocking solution and incubated in the wells overnight at 4°C. Secondary antibody of horseradish peroxidase-conjugated goat anti-mouse IgG (1030-05; Southern Biotech, Birmingham, AL, USA), diluted 1:5,000 in PBS–1% BSA, was added after extensive washing and incubated for a further hour before washing. The substrate, tetramethylbenzidine (TMB; KOMA BIOTECH) 100 μl/well, was applied for 20 min before stopping the reaction with 25 μl/well of 1 M H2SO4 (Sigma-Aldrich), after which the optical density was read at 450 nm (VERSAmax; Molecular Devices, Sunnyvale, CA, USA).
TgAA was detected in the culture supernatants of splenocytes after removing the spleens from four mice per group on day 28 and culturing them at 37°C and in a humidified atmosphere (2.5 × 105 cells/well) in a final volume of 200 μl with or without concanavalin A (ConA; 2.5 μg/ml; Sigma-Aldrich) and lipopolysaccharide (LPS; 2.5 μg/ml; Sigma-Aldrich) in 96-well flat-bottomed plates. After 72 h, cell culture supernatants from individual wells were collected and stored at −70°C until use. The procedure for the detection of pTg antibodies in cell culture supernatants was the same as that described above, with the exception that the samples were diluted 1:100 in PBS–1% BSA. These culture supernatants were also used for the analysis of the levels of multiple cytokines.
Measurement of Thyroxine (T4) and Thyroid-Stimulating Hormone (TSH)
The levels of T4 and TSH in serum samples were assayed using commercial ELISA kits (Rodent T4 and Rodent TSH, respectively, Endocrine Technologies, Inc., Newark, CA, USA). According to the manufacturer, the minimum detectable concentration of T4 by the rodent T4 ELISA kit is 2 ng/ml and that of TSH by the rodent TSH ELISA kit is 0.2 ng/ml.
Multiple Cytokine Levels in Sera and Splenocyte Culture Supernatants
Serum samples or splenocyte culture supernatants were assayed using a multiplex cytokine ELISA kit for tumor necrosis factor-α (TNF-α), interferon (IFN)-γ, interleukin (IL)-1α, IL-1β, IL-2, IL-4, IL-6, IL-10, IL-12p(40), IL-15, IL-17, monocyte chemotactic protein-1 (MCP-1), and regulated upon activation normal T-cell expressed and secreted (RANTES) (Millipore).
Flow Cytometric Analysis of Splenic Cell-Surface Markers
FITC-conjugated rat anti-mouse CD3, PE-conjugated rat anti-mouse CD4, PE-conjugated rat anti-mouse CD8a, PE-conjugated rat anti-mouse CD11b, and PE-conjugated rat anti-mouse CD45R/B220 (BD Pharmingen™) were used for the flow cytometric analysis of spleen cells. Cell analysis was performed with a FACSCalibur flow cytometer (BD Biosciences) using CELLQuest software, as previously described (11).
Determination of the Proportion of Treg Cells by Flow Cytometry
Spleen cells were analyzed for Treg cell markers using FITC-conjugated rat anti-mouse CD4, allophycocyanin (APC)-conjugated rat anti-mouse CD25, and PE-conjugated rat anti-mouse forkhead box P3 (FoxP3) antibodies (all from BD Biosciences) according to our previous study (12).
Determination of the Proportion of CD4+IL-17+ Cells by Flow Cytometry
Spleen cells were analyzed to determine the proportion of CD4+IL-17+ cells using FITC-conjugated rat anti-mouse CD4 and peridinin–chlorophyll protein–cyanine 5.5 (PerCP-cy5.5)-conjugated rat anti-mouse IL-17A antibodies (both from BD Biosciences). Before intracellular staining, spleen cells were stimulated for 4 h with 25 ng/ml of phorbol myristate acetate (Sigma-Aldrich) and 250 ng/ml of ionomycin (Sigma-Aldrich) in the presence of GolgiStop (BD Biosciences). Intracellular staining was conducted using an intracellular staining kit (BD Biosciences) according to the manufacturer's protocol. Flow cytometric analysis was performed on a FACSAria instrument (BD Biosciences).
Histopathology of Thyroid Glands
Thyroid glands were cryosectioned at 4-μm thickness, fixed, and immunostained for CD3. Briefly, serial 4-μm-thick sections were obtained and washed with PBS, pH 7.2, followed by treatment with 1% PBS/H2O2 (51-7548KE; BD Biosciences) solution to block endogenous peroxidase. Nonspecific binding was blocked by 10% goat nonimmune serum blocking solution (Invitrogen). Anti-mouse CD3 rat IgG (clone 17A2; Biolegend, San Diego, CA, USA) diluted 1:100, was used subsequently and incubated for 1 h at room temperature. The sections were then incubated with biotinylated anti-rat Ig (1: 50, 51-7605KC; BD Biosciences) for 30 min at room temperature, washed three times in PBS, and incubated with streptavidin– horseradish peroxidase (51-7547KC; BD Biosciences) for 30 min. Color was detected using a solution of 0.05% diaminobenzidine (51-7549KE; BD Biosciences) and 0.2% H2O2 at room temperature for 10 min. Sections were counterstained with hematoxylin (BBC Biochemical, Dallas, TX, USA) and coverslipped using mounting medium (#4112, Thermo Fisher Scientific Inc.). Primary antiserum was substituted with PBS in some sections to constitute the negative control.
Thyroid glands were fixed in 10% neutral buffered formalin (Sigma-Aldrich), embedded in paraffin, sectioned at 4 μm, stained with hematoxylin and eosin (H&E; BBC Biochemical), and then examined with the aid of an Olympus CKX41 light microscope (Tokyo, Japan). Mononuclear cell infiltration in the thyroid gland was graded as previously described (10).
Biodistribution of Transplanted Mouse ATMSCs
Five mice from both the syngeneic and the allogeneic ATMSC groups were injected with Cell Tracker (CM-DiI) dyelabeled (red fluorescence) ATMSCs for the cell biodistribution study. The presence of CM-DiI-labeled cells was examined in various tissues including thyroid, spleen, liver, lung, kidney, heart, and muscle. Five sections were analyzed from each tissue. The cells were counterstained with DAPI mounting medium (Vector Laboratories, Inc., Burlingame, CA, USA) and observed under a Radiance 2100 laser scanning confocal microscope (Bio-Rad, Hercules, CA, USA). Excitation wavelengths of 405 and 543 nm were used.
Humoral Response to Infused Cells
For the detection of antibodies against administered ATMSCs, flow cytometric cross-matching for circulating anti-C57BL/6 ATMSC antibodies or anti-BALB/c ATMSC antibodies was conducted using sera collected from each recipient and from control mice. Specifically, recipient mouse sera (1:25 dilution in DPBS) and control mouse sera (non-ATMSCs infused, negative control) were incubated with donor mouse ATMSCs for 30 min at 37°C. After washing, mouse ATMSCs were incubated with FITC-conjugated anti-mouse IgG antibody (1:1,000 per 106 ATMSCs; AP308F; Millipore) for 1 h at 4°C, then washed in DPBS and assayed in a flow cytometer (FACSAria, BD Biosciences). Fluorescence signals from 10,000 cells were counted and the percentage of FITC-positive cells recorded. Circulating anti-human ATMSC IgG antibodies were also evaluated in recipient and control mouse sera collected from the previous study (11).
Statistical Analysis
All results are expressed as means ± SEMs (standard error of the mean). Data obtained from different groups were compared using a one-way analysis of variance (ANOVA) followed by either post hoc Turkey's multiple comparison tests or Dunnett's test. Differences with a confidence level of 95% or higher were considered statistically significant (p < 0.05). All statistical analyses were conducted using SPSS version 17.0 (SPSS Inc., Chicago, IL, USA).
Results
Preparation of Mouse ATMSCs
A lot of debris from connective tissues and red blood cells (RBCs) were seen along with ATMSCs in the isolation step. Since ATMSCs can adhere to plastic, the nonadherent cells in the culture were removed during changes of medium, and the spindle-shaped ATMSCs were retained. After passage 3, mouse ATMSCs in the modified DMEM exhibited increased cell size and a decreased proliferation rate and a halt in proliferation after passage 5. These expansion limitations were seen in ATMSCs from both C57BL/6 and BALB/c strains.
Comparison of Mouse ATMSC Immunophenotype Characteristics
The immunophenotypic characteristics of each of the ATMSCs from C57BL/6 and BALB/c mice were analyzed by flow cytometry. We also used commercial available media, StemXVivo™, to compare MSC immunophenotypes. StemXVivo™ medium maintained ATMSCs in an undifferentiated morphology with constant cell size and proliferation rate even at passage 5. There were no significant differences in the expression of cell surface antigens between cells cultured in the modified DMEM and StemXVivo™ medium. The FACS results are summarized in Table 1. ATMSCs from both mouse strains positively expressed CD29, CD44, and CD90.2 and did not express CD90.1, CD31, CD34, and CD45. Unlike human ATMSCs, the expression of CD73 was mild (C57BL/6) or mild moderate (BALB/c), and that of CD105 was moderate (both strains). The CD73 and CD105 markers are known to be strongly expressed in human ATMSCs. ATMSCs from BALB/c positively expressed MHC Class I (H-2Kd), but those from C57BL/6 did not (H-2Kb). ATMSCs from both mouse strains (C57BL/6 and BALB/c) did not express MHC class II (I-Ab and I-Ad, respectively).
Comparison of Immunophenotypes of Adipose Tissue-Derived Stem Cells in Different Mouse Strains (n = 4–5)

Data are expressed as mean ± SD (%, except for cell size), SD, standard deviation; P, passage; CD, cluster of differentiation; MHC, major histocompatibility complex.
Differentiation Capacity of Mouse ATMSCs
To determine whether mouse ATMSCs differentiate into different cell types, we cultured ATMSCs from both mouse strains in the appropriate differentiation media. The ATMSCs showed differentiation capacity, with no differences between the two strains. As shown in Figure 1, ATMSCs cultured in the modified DMEM remained undifferentiated (Fig. 1A). The undifferentiated status of ATMSCs was also confirmed by Oil Red O, Alizarin Red S, and alcian blue staining (data not shown). ATMSCs differentiated into adipocytes containing Oil Red O-positive lipid droplets (Fig. 1B). Alizarin Red S staining confirmed the osteogenic differentiation of ATMSCs. Intensive Alizarin Red S staining indicated that calcium deposition had occurred (Fig. 1C). Upon chondrogenesis, the pellets of ATMSCs showed chondrogenic morphology when stained with toluidine blue (Fig. 1D), and upon neurogenesis, cells positive for NSE (Fig. 1E) and neuron-specific class III β-tubulin (Fig. 1F) were detected.

Differentiation of mouse adipose tissue-derived stem cells (ATMSCs) into adipocytes, osteocytes, chondrocytes and neuronal cells. (A) ATMSCs maintained in control medium remained undifferentiated. (B) ATMSCs differentiated into adipocytes contained Oil Red O-positive lipid droplets. (C) ATMSCs differentiated into osteocytes, which were stained with Alizarin Red S. (D) Aggregated chondrocytes derived from ATMSCs were stained with toluidine blue. Two weeks after neurogenic differentiation, the cells stained positive for neuronal-specific enolase (NSE) (E) and neuron-specific class III β-tubulin (antibody name: TuJ-1) (F).
Spleen-to-Body Weight Ratio
We examined whether the immune reactions occurred due to the systemic transplantation of allogeneic ATMSCs in a mouse model. There were no particular behavioral changes among the groups during observation. The spleen of each mouse was harvested immediately after necropsy. Although the mean body weights were similar among groups, the mean spleen weights of both ATMSC-treated groups were lower than that of the control group. Spleen-to-body weight ratios in the syngeneic and allogeneic ATMSC groups were significantly lower than that of the control group (p = 0.028). The calculated values were 0.00708 ± 0.00057, 0.00550 ± 0.00032, and 0.00586 ± 0.00028 in the control group, syngeneic, and allogeneic ATMSC groups, respectively [the calculated value of age-matched normal female C57BL/6 mice (n = 9) was 0.00377 ± 0.00014].
TgAA Levels in Sera and in Culture Supernatants of Splenocytes
In autoimmune thyroiditis, antibodies against thyroglobulin are produced, and they cause the gradual destruction of follicles in the thyroid gland. To demonstrate the immunomodulatory properties of MSCs, we examined whether TgAA levels were reduced by the systemic transplantation of ATMSCs. Elevated TgAA levels were found in the sera and in splenocyte culture supernatants of the autoimmune thyroiditis (control group) (Fig. 2). The levels of serum TgAA in ATMSC-treated groups (both syngeneic and allogeneic) were considerably lower than those in the control group (p = 0.011) (Fig. 2A). Syngeneic ATMSCs significantly reduced TgAA levels in splenocyte culture supernatants (p = 0.037) (Fig. 2B) and in culture supernatants of LPS-stimulated splenocytes (p = 0.004) (Fig. 2C) in comparison to those in the control group. Allogeneic ATMSCs also significantly reduced TgAA levels in culture supernatants of LPS-stimulated splenocytes; however, the difference in TgAA levels in culture supernatants of nonstimulated splenocytes did not reach statistical significance.

Absorbance of thyroglobulin autoantibody (TgAA) from control, syngeneic, and allogeneic adipose tissue-derived stem cells (ATMSCs). Normal saline, half a million C57BL/6 ATMSCs, and half a million BALB/c ATMSCs were administered to mice in the control, syngeneic, and allogeneic groups, respectively. One week after the last cell transplantation, blood samples and spleen cells were collected. Spleen cells from the mice were cultured with or without lipopolysaccharide (LPS). The absorbance of TgAA was analyzed (A) in sera and in culture supernatants of splenocytes either (B) without or (C) following LPS stimulation. Data obtained from the groups were compared using one-way analysis of variance and a post hoc multiple comparison. *Significant (p < 0.05) differences from the control.
T4 and TSH
We examined changes in the levels of T4 and TSH (hormones related to thyroid function) in sera from mice with autoimmune thyroiditis. The concentrations of T4 were 5.2 ± 0.8, 8.0 ± 0.2, and 7.7 ± 1.6 μg/dl in the control, syngeneic, and allogeneic groups, respectively (mean ± SEM; reference range: 4.5–12.6). Serum TSH levels were also measured in the control, syngeneic group, and allogeneic groups (n = 10). TSH in the control group was undetectable except for in three mice (0.36, 8.66, and 0.28 ng/ml). TSH was undetectable in the syngeneic group, while it was undetectable in all but one mouse of the allogenic group (0.3 ng/ml). TSH from 12 normal healthy controls was also undetectable except for in four mice (0.23, 0.26, 0.50, 0.57 ng/ml).
Multiple Cytokine Levels in Sera and in Culture Supernatants of Splenocytes
Changes in cytokine expression upon syngeneic or allogeneic ATMSC transplantation were investigated in sera from autoimmune thyroiditis mice. Mean serum cytokine levels in each group are summarized in Table 2. There were no differences in cytokine expression patterns between the syngeneic and allogeneic ATMSC groups. The mean serum levels of IFN-γ, IL-6, and TNF-α, and the ratio of IFN-γ/IL-4 decreased approximately two- to threefold in both types of ATMSC transplantation compared to those seen in the control group. Further, the serum level of IL-2 decreased significantly.
Comparison of the Levels of Various Cytokines and IFN-γ/IL-4 Ratios in Sera Taken From Mice With Experimental Autoimmune Thyroiditis (pg/ml, Except for the IFN-γ/IL-4 Ratio)

Data obtained from groups were compared using a one-way analysis of variance and a post hoc multiple comparison. ATMSCs, adipose tissue-derived stem cells; IFN, interferon; IL, interleukin; MCP-1, monocyte chemotactic protein-1; RANTES, regulated on activation, normal T-cell expressed and secreted; TNF, tumor necrosis factor.
Significant (p < 0.05) differences from the control.
Analysis of the expression levels of various cytokines was also conducted in the culture supernatants of splenocytes. In ConA-stimulated splenocytes, IFN-γ levels and IFN-γ/IL-4 ratios of both ATMSC-treated groups were significantly lower than those of the control group (p = 0.006 and p = 0.014, respectively) (Table 3). RANTES levels from the culture supernatants of LPS-stimulated splenocytes significantly decreased in the syngeneic (539.1 ± 99.8 pg/ml) and allogeneic (623.1 ± 115.5 pg/ml) ATMSC groups in comparison to the control group (1,384.9 ± 180.8 pg/ml, p = 0.001). TNF-α levels significantly decreased only in the allogeneic ATMSC group (288.4 ± 91.7 pg/ml, p = 0.037) compared to the control group (553.8 ± 96.5 pg/ml). These levels showed a tendency to decrease in the syngeneic ATMSC group (308.5 ± 37.5 pg/ml), but this did not reach statistical significance.
Comparison of the Levels of Various Cytokines and IFN-γ/IL-4 Ratios in Culture Supernatants From Spleen Cells Stimulated With ConA (pg/ml, Except for the IFN-γ/IL-4 Ratio)

Data obtained from groups were compared using a one-way analysis of variance and a post hoc multiple comparison. ATMSCs, adipose tissue-derived stem cells. ConA, concanavalin A.
Significant (p < 0.05) differences from the control.
Flow Cytometric Analysis of the Splenic Cell Surface Markers
Spleen cell surface markers were examined to determine whether syngeneic or allogeneic ATMSC transplantation influenced the immunophenotype proportions of splenocytes from autoimmune thyroiditis mice. Flow cytometric analysis showed no significant differences in the expression of CD3, CD4:CD8, CD11b, and B220 among the groups. Compared to the control group, CD11b (macrophage-1 antigen; MAC-1)-positive spleen cells in both ATMSC-treated groups showed a tendency to decrease, but these differences did not reach statistical significance (data not shown).
Determination of the Proportion of Treg Cells by Flow Cytometry
Spleen cells were analyzed to determine the proportion of Treg cells. The proportions of CD4+FoxP3+ cells were 1.4 ± 0.4 (n = 3), 2.3 ± 0.3 (n = 4), and 3.0 ± 0.8 μg/dl (n = 4) in the control, syngeneic, and allogeneic groups, respectively (mean ± SEM, p = 0.245). The proportions of CD4 + CD25 + FoxP3 + cells were 1.1 ± 0.3 (n = 3), 1.6 ± 0.2 (n = 4), and 2.6 ± 0.7 μg/dl (n = 4) in the control, syngeneic, and allogeneic groups, respectively (mean ± SEM, p = 0.193). Treg cell expansion in the spleen of MSC-treated mice showed a tendency to increase but did not reach statistical significance.
Determination of the Proportion of CD4+IL-17+ Cells by Flow Cytometry
The proportions of CD4+IL-17+ cells were 0.9 ± 0.1 (n = 3), 0.6 ± 0.1 (n = 4), and 0.5 ± 0.03 μg/dl (n = 3) in the control, syngeneic, and allogeneic groups, respectively (mean ± SEM, p = 0.101). The proportion of CD4+IL-17+ cells in the spleen of MSC-treated mice showed a tendency to decrease but did not reach statistical significance.
Histopathology of Thyroid Glands From Autoimmune Thyroiditis Mice
Mononuclear cell infiltration in the thyroid gland significantly decreased in both ATMSC-treated groups in comparison to the control group (p = 0.012). The mean infiltration index was 1.6 ± 0.3, 0.4 ± 0.3, and 0.5 ± 0.2 in the control, syngeneic, and allogeneic groups, respectively (mean ± SEM). Autoimmune thyroiditis is characterized by invasion of the thyroid tissue by leukocytes, mainly T-lymphocytes. Thus, we conducted immunohistochemical analyses of thyroid glands using antibodies against the T-cell marker CD3 and compared the infiltration of CD3+ cells among the groups. In the control group, immunoperoxidase staining of frozen thyroid sections showed that CD3+ cells had infiltrated between the follicles of the thyroid glands. However, CD3+ cells were hardly seen in the thyroid glands of both ATMSC-treated groups, suggesting that ATMSCs suppressed T-lymphocyte infiltration into the thyroid glands (Fig. 3).

Immunohistochemical staining of mouse thyroid for CD3. Thyroids were removed 1 week after the last cell transplantation. Immunohistochemical staining for cluster of differentiation 3 (CD3) was performed in thyroid tissues from (A) the saline-treated control group, (B) the syngeneic ATMSC-treated group, and (C) the allogeneic ATMSC-treated group (magnification: 400×). CD3+ cells (arrows) were found in the thyroiditis control group, but not in the ATMSC-treated groups.
Biodistribution of Transplanted ATMSCs
Five mice each from the syngeneic and allogeneic groups were administered fluorescently labeled (CM-DiI red fluorescent tracker dye) ATMSCs for identification in histopathological sections (Fig. 4). Five sections were analyzed from each tissue. CM-DiI-labeled ATMSCs were mostly present in the spleen (Fig. 4E, F), and some CM-DiI-labeled ATMSCs were presented in the lung and liver. In other organs, such as the thyroid (Fig. 4B, C), a few very small fluorescent dots were seen, but they were not considered as CM-DiI-labeled ATMSCs due to their weak signals.
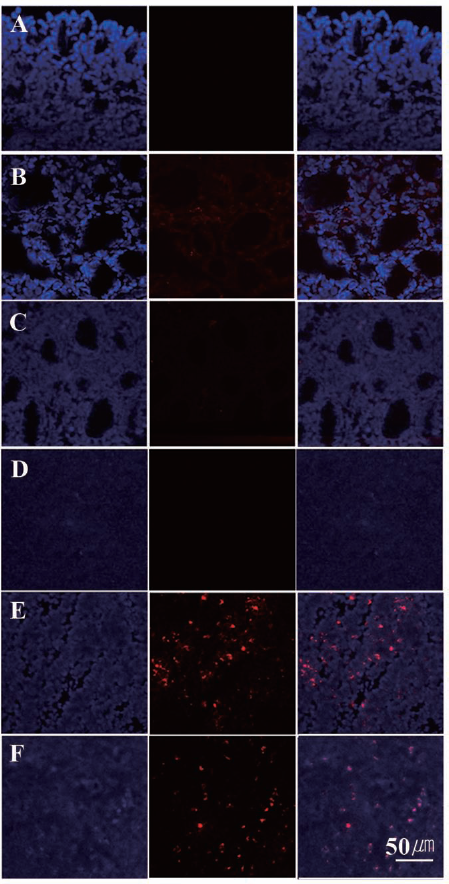
Biodistribution of ATMSCs after intravenous injection. The existence of fluorescently labeled (CM-DiI, red) adipose tissue-derived stem cells (ATMSCs) was examined in various organs including thyroids (A, B, C) and spleens (D, E, F). Tissues were obtained from the saline-treated control group (A, D), the syngeneic ATMSC-treated group (B, E), and the allogeneic ATMSC-treated group (C, F). Fluorescently labeled cells were visualized mostly in the spleens of ATMSC-treated mice (red, middle column). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; blue, left column), and the images were merged (right column).
Humoral Response to Infused Cells (ATMSCs)
All groups exhibited increases in humoral responses to the administered ATMSCs after four administrations compared to after a single administration. The extent of the humoral response on syngeneic and allogeneic ATMSCs was similar after a single administration of stem cells once. However, the level of humoral response to allogeneic ATMSCs substantially increased compared to the response to syngeneic ATMSCs after four administrations (Table 4). The level of IgG against xenogeneically transplanted cells was higher than that against allogeneic transplanted cells (Table 4).
Humoral Response to Infused Adipose Tissue-Derived Stem Cells (ATMSCs): Percentage of IgG-Positive Transplanted Cells (%)

Discussion
In this study, we investigated the potential benefits of ATMSCs on experimental autoimmune thyroiditis (EAT). EAT shares a number of clinical, histological, and immunological features with Hashimoto's thyroiditis. EAT model mice were prepared according to the procedure described in Chen et al. by using the inoculation of thyroglobulin with adjuvant (6). EAT model mice are typically killed between day 21 and 28 after the first immunization to analyze the outcome of interest (6). Autoimmune thyroiditis is characterized by elevated TgAA levels and infiltration of lymphocytes into the thyroid. Although autoimmune thyroiditis is an organ-specific autoimmune disease, it can influence systemic cytokine profiles, leading to exaggerated synthesis of proinflammatory cytokines (10). Therefore, the restoration of systemic cytokine levels is another valuable factor to verify improvement in autoimmune thyroiditis. Our previous study revealed that human ATMSCs could reduce host immune responses and improve the Th1/Th2 balance by downregulating Th1 cytokines even when transplanted xenogeneically (11). For further clinical applications in humans, it is important to know the differences between autologous and allogeneic stem cell transplantation, especially for autoimmune diseases. Therefore, we compared the characteristics of MSCs derived from adipose tissue from two different strains and analyzed efficacy and safety differences between syngeneic and allogeneic ATMSCs.
The initial culture of mouse ATMSCs contained heterogenous cell populations. One population of round cell clusters could be eliminated by negative selection following enzyme treatment in subculture. Mouse ATMSCs positively expressed CD29, CD44, and CD90 and did not express CD31, CD34, and CD45. CD90 can be used as a marker for a variety of stem cells. In mice, there are two alleles: CD90.1 [thymocyte antigen 1.1 (Thy1.1), Thy 1a] and CD90.2 (Thy1.2, Thy 1b). The CD90.2 marker is expressed in most strains of mice, whereas CD90.1 is expressed in some strains such as the AKR/J (interleukin 3 receptor, α chain, mutation 1) and PL (phosphodiesterase 6B) mouse strains (http://www.sinobiological.com/CD90-THY-1-a-4516.html). ATMSCs from both C57BL/6 and BALB/c mice were positive for CD90.2 and negative for CD90.1. In some samples, CD34 was mildly expressed. Unlike human ATMSCs with high purity, CD73 and CD105 (positive markers in human MSCs) were only partially expressed in mouse ATMSCs. The MHC class I expression pattern was different between the two strains. The immunophenotypes of ATMSCs from passages 1–5 cultured in StemXVivo™ medium were similar. Moreover, immunophenotyping results acquired from ATMSCs from passages 1 and 2 cultured in modified DMEM, and from those cultured in STEMXVivo™ medium, were similar.
Although the immunophenotypes of mouse ATMSCs from these two strains were not identical, both exhibited capacities for adipogenic, osteogenic, chondrogenic, and neurogenic differentiation when cultured in the presence of established lineage-specific differentiation factors. There were no differences in differentiation abilities between ATMSCs from the two mouse strains. We conducted in vivo experiments with a murine EAT model and collected data including spleen-to-body weight ratios, TgAA levels, systemic cytokine levels, splenocyte immunophenotypes, and thyroid histopathology. Our data in this study shows that allogeneic mouse ATMSCs are capable of protecting against lymphocyte infiltration and of restoring the Th1/Th2 balance as well as syngeneic ATMSCs. These results are in accordance with our previous EAT study using human ATMSCs (11).
The mean spleen-to-body weight ratio was increased in all mice with autoimmune thyroiditis compared with normal mice. However, syngeneic or allogeneic transplantation of ATMSCs decreased the spleen-to-body weight ratio significantly compared with saline treatment. Thus, spleen enlargement was probably due to immunization with “porcine Tg” foreign protein but was reduced by the transplantation of ATMSCs. Serum TgAA levels were elevated upon EAT induction, but the concentrations of T4 and TSH remained within the reference ranges. These findings can be ascribed to the fact that the animals did not develop hypothyroidism. Serum TgAA levels in syngeneic or allogeneic ATMSC groups were considerably lower than in the control group. TgAA levels in culture supernatants of LPS-stimulated splenocytes decreased significantly in both the syngeneic and allogeneic ATMSC-treated groups in comparison to the control group.
EAT is considered a model of Th1-mediated disease. In vivo administration of IFN-γ exacerbates the severity of the pathology (38), and neutralization of IFN-γ reduces the incidence and the severity of EAT (43). ATMSC transplantation, both syngeneic and allogeneic, decreased the average serum levels of the cytokines IFN-γ, IL-6, and TNF-α approximately two- to threefold and significantly decreased the serum IL-2 levels, as well as the ratio of IFN-γ/IL-4, in comparison to those of the control group. ATMSC treatment significantly decreased the levels of IFN-γ and RANTES and the IFN-γ/IL-4 ratio from the culture supernatants of mitogen-stimulated splenocytes. Th17 cells, distinct from Th1 or Th2 cells, are known to play a crucial role in the induction of autoimmune diseases (28). The proportion of CD4+IL-17+ cells in the spleens of ATMSC-treated mice showed a tendency to decrease, although it did not reach statistical significance.
Regardless of their syngeneic or allogeneic origin, ATMSCs were able to decrease TgAA production and suppress the production of Th1-type cytokines. However, the differences in humoral immune responses were evident. IgG production against allogeneic ATMSCs (BALB/c–ATMSCs) was more pronounced than syngeneic ATMSCs (C57BL/6–ATMSCs). C57BL/6–ATMSCs lacked MHC Class I (H-2Kb), whereas BALB/c–ATMSCs expressed MHC class I (H-2Kd) according to our results of immunophenotyping. The result of immunophenotyping suggested that H-2Kd would be conducive to elevated humoral responses against BALBc–ATMSCs relative to C57BL6–ATMSCs. It can be a possible explanation for the stronger antibody response to allogeneic ATMSCs.
The strongest humoral immune response was induced by xenogeneic (human) ATMSC transplantation, followed by allogeneic and then syngeneic administration, in that order. Human ATMSCs as well as mouse ATMSCs showed therapeutic effects on EAT, so these antibodies induced by the humoral response might be binding antibodies, not neutralizing antibodies.
Another key feature of MSCs is their ability to selectively migrate into sites of injury. Our histopathological data demonstrated that the administration of ATMSCs (both syngeneic and allogeneic) to EAT mice suppressed T-lymphocyte infiltration into the thyroid glands. We therefore examined the biodistribution of systemic transplanted ATMSCs to ascertain whether the stem cells affect the thyroid directly or indirectly. Unexpectedly, fluorescently labeled ATMSCs were mostly found in the spleen, not the thyroid. ATMSCs were administered in the early stages of EAT, so their migration to the spleen might be the result of ATMSCs primarily functioning in systemic immune modulation.
It was reconfirmed that ATMSCs were effective in treating autoimmune thyroiditis. In this study, syngeneic ATMSCs were isolated from healthy mice without EAT. However, in reality, the possibility of autologous stem cells being prepared from patients with diseases is high. It is not clear whether these ATMSCs will retain the same function as do ATMSCs from healthy donors. Further study is needed in order to explore this question by comparing syngeneic stem cells acquired from animals that have spontaneously or genetically induced autoimmune diseases and allogeneic stem cells from animals without diseases.
Footnotes
Acknowledgments
The authors thank the staff of the Laboratory Animal Research Center, Samsung Biomedical Research Institute, for their technical support and assistance. Special thanks to Mr. David Lee for his grammatical revisions and constructive criticisms toward enhancing the fluidity of this article. This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF-2012R1A1A2006701) to E. W. Choi. Further support was provided by a grant from the Samsung Biomedical Research Institute (SBRI AB20011/SMX1131211/SMX1140141) to E. W. Choi. The authors declare no conflicts of interest.