
Abstract
Hepatocyte transplantation has been used to treat liver disease. The availability of cells for these procedures is quite limited. Human embryonic stem cells (hESCs) and induced pluripotent stem cells (hiPSCs) may be a useful source of hepatocytes for basic research and transplantation if efficient and effective differentiation protocols were developed and problems with tumorigenicity could be overcome. Recent evidence suggests that the cell of origin may affect hiPSC differentiation. Thus, hiPSCs generated from hepatocytes may differentiate back to hepatocytes more efficiently than hiPSCs from other cell types. We examined the efficiency of reprogramming adult and fetal human hepatocytes. The present studies report the generation of 40 hiPSC lines from primary human hepatocytes under feeder-free conditions. Of these, 37 hiPSC lines were generated from fetal hepatocytes, 2 hiPSC lines from normal hepatocytes, and 1 hiPSC line from hepatocytes of a patient with Crigler–Najjar syndrome, type 1. All lines were confirmed reprogrammed and expressed markers of pluripotency by gene expression, flow cytometry, immunocytochemistry, and teratoma formation. Fetal hepatocytes were reprogrammed at a frequency over 50-fold higher than adult hepatocytes. Adult hepatocytes were only reprogrammed with six factors, while fetal hepatocytes could be reprogrammed with three (OCT4, SOX2, NANOG) or four factors (OCT4, SOX2, NANOG, LIN28 or OCT4, SOX2, KLF4, C-MYC). The increased reprogramming efficiency of fetal cells was not due to increased transduction efficiency or vector toxicity. These studies confirm that hiPSCs can be generated from adult and fetal hepatocytes including those with genetic diseases. Fetal hepatocytes reprogram much more efficiently than adult hepatocytes, although both could serve as useful sources of hiPSC-derived hepatocytes for basic research or transplantation.
Keywords
Introduction
Hepatocyte transplantation has been proposed as a cellular therapy for metabolic liver disease and acute liver failure. Primary hepatocyte transplantation has been used to partially correct patients with Crigler–Najjar syndrome type 1 (CN-1), urea cycle, glycogen storage disorders, and other metabolic liver diseases (1,3,5). This therapy shows promise despite continuing issues with cell engraftment and proper immunosuppression. The availability of transplantable hepatocytes remains an ongoing problem.
Stem cell-derived hepatocytes from human embryonic stem cell (hESC) (22) and induced pluripotent stem cell (hiPSC) (20,23) technology have the potential to provide a nearly unlimited source of hepatocytes for basic research or in vitro drug discovery if efficient protocols were developed to produce mature hepatocytes; however, issues concerning tumorigenicity still limit the clinical application of hESCs or hiPSCs. Furthermore, hiPSC generation is limited by low reprogramming efficiency as well as long duration from induction to cell line establishment. hiPSCs have been derived from multiple cell types including fetal lung and neonatal foreskin fibroblasts, mesenchymal stem cells (14), mouse adult and fetal hepatocytes (7,9,18), amnion epithelial (13,24) and mesenchymal cells from the umbilical cord matrix or amnion (2), and even one report with human adult hepatocytes (11). Furthermore, hiPSCs have also been derived from fibroblasts from patients with different genetic disorders, including inherited metabolic disorders of the liver (16). These disease-specific cell lines potentially serve as an unlimited source of disease-specific cells if they could be differentiated back to a phenotype that faithfully recreates the disease. Theoretically, disease-specific cells could be genetically corrected and used for autologous cell transplantation to correct the phenotype. The idea of patient-specific therapy has recently been put into question with a report that differentiated mouse iPSCs induce T-cell-dependent immune responses in syngeneic recipients (25).
Several groups have tried to increase reprogramming efficiencies by using all six transcription factors (10), by combination of transcription factors and chromatin modulation (4,17), or by addition of chemical compounds (12). However, if populations of cells are more easily reprogrammed, methods to increase reprogramming may not be necessary, and these cell populations could be useful for translational research and could also be good models for understanding the mechanisms of the reprogramming process.
Recently, an increased reprogramming capacity of mouse liver progenitor cells compared with differentiated liver cells was reported (7). There is evidence that mouse iPSCs retain epigenetic markings providing “memory” of the cell from which they were made (6) and that cell type of origin influences in vitro differentiation potential (15). A recent report demonstrated hepatic lineage stage-specific donor memory in mouse iPSCs, and interestingly, fetal hepatocyte-derived iPSCs demonstrated superior capacity for hepatic redifferentiation (9). Therefore, iPSCs from fetal human hepatocytes might be a particularly useful source of stem cell-derived hepatocytes.
There are no reports describing derivation of hiPSCs from human fetal hepatocytes or from hepatocytes from patients with an inborn error in metabolism. Here we provide a method to generate hiPSCs from adult and fetal human hepatocytes under entirely feeder-free conditions and report that reprogramming of fetal hepatocytes was over 50-fold more efficient than adult hepatocytes and can be accomplished with only three reprogramming factors.
Materials and Methods
Use of Animal- and Human-Derived Tissue
All animal experiments were performed in accordance with the University of Pittsburgh's Institutional Animal Care and Use Committee. Furthermore, all human tissue was obtained with informed consent and approved by the University of Pittsburgh's Institutional Review Board.
Cell Culture
The H1 hESC and all hiPSC lines were maintained on Matrigel (BD Biosciences, San Jose, CA, USA) and cultured in mTeSR™1 medium (Stem Cell Technologies, Vancouver, BC, Canada). Cells were passaged using 1 mg/ml dispase (Gibco, Carlsbad, CA, USA) using conventional stem cell culture techniques. All cell lines used for data collection were passage 30 or under. The male H1 hESC line was obtained from the WiCell Research Institute, Inc. (Madison, WI, USA). Primary human hepatocytes (see Table 1 for ages and gender) were isolated using a collagenase perfusion method (8,19). Viable cell count was used for plating, and plates were washed extensively to get rid of dead and unattached cells. Primary human hepatocytes were cultured in hepatocyte maintenance medium (HMM) (Lonza, Walkersville, MD, USA) and supplemented with 1× Penstrep (Invitrogen, Carlsbad, CA, USA) and 10−7 M insulin and dexamethasone (both Lonza). Primary human fetal hepatocytes (see Table 1 for ages and gender) were isolated by digesting the tissue in Eagle's minimum essential medium (EMEM; Lonza, Walkersville, MD, USA) containing 0.5 mg/ml of collagenase (Type XI, Sigma-Aldrich, St. Louis, MO, USA; Cat. #C7657) on a lab quake shaker/rotisserie (Barnstead Intl., Dubuque, IA, USA) for 40 min and otherwise processed as described for adult hepatocytes. Viable cell counts were used for plating, and cultures were changed out with fresh media 2 h after plating and washed extensively 18–24 h postplating to remove dead and unattached cells. Primary human fetal hepatocytes were cultured in Dulbecco's modified Eagle's medium (DMEM; Lonza) supplemented with 10% fetal bovine serum (FBS PAA Laboratories, Pasching, Austria), 10−7 M insulin, 1× Penstrep, and 10 ng/ml epidermal growth factor (EGF; BD Biosciences).
Details of Human-Induced Pluripotent Stem Cell (hiPSC) Induction Experiments on Adult and Fetal Hepatocytes

Information includes donor number, age, sex, viability of cells after isolation, and the number of hiPSC lines identified. OSKMNL = octamer-binding transcription factor 4 (OCT4), sex-determining region Y box 2 (SOX2), Krüppel-like factor 4 (KLF4), V-Myc myelocytomatosis viral oncogene homolog (C-MYC), NANOG, and LIN28.
hiPSC Inductions
Primary human adult and fetal hepatocytes were added to Matrigel-coated six-well plates (BD Biosciences) at 100,000 cells/well in their respective media and allowed to attach overnight. The following day, transduction media were given to the cells that consisted of each cell type's respective culture medium supplemented with 6 μg/ml polybrene (Millipore, Billerica, MA, USA) and lentiviruses delivering up to six reprogramming factors, octamer-binding transcription factor 4 (OCT4), sex-determining region Y box 2 (SOX2), Krüppel-like factor 4 (KLF4), V-Myc myelocytomatosis viral oncogene homolog (C-MYC), NANOG, and LIN28, (Thermo Fisher Scientific, Waltham, MA, USA) each at a targeted multiplicity of infection (MOI) of up to 10 (day 0). After 24 h, the cells were washed with DMEM/F12 (Thermo Fisher Scientific) three times. Cells were cultured in their respective media, and media were changed to mTeSR™1 on days 1–7. Media were refreshed every 2–3 days. Large hiPSC colonies appeared between days 16 and 21, and individual colonies were selected for expansion between days 25 and 35. Colony formation was determined by counting total number of individual hESC-like colonies formed in all wells. Reprogramming efficiency was determined by the following equation: (total number of hESC-like colonies/total number of cells plated) × 100. Details of each individual experiment (n = 18 adult and n = 8 fetal) can be found in Table 1.
Immunocytochemistry (ICC)
hiPSCs were plated on Matrigel-coated coverslips in 24-well plates (both BD Biosciences), fixed using 4% paraformaldehyde (PFA; Sigma-Aldrich) for 20 min at room temperature and permeabilized with 0.1% Triton X (Sigma-Aldrich) for 15 min for OCT4, SOX2, and NANOG at room temperature. Cells were not permeabilized for stage-specific embryonic antigen 4 (SSEA4) staining. Cells were rehydrated with phosphate-buffered saline (PBS; Gibco) and 0.5% bovine serum albumin (BSA; Sigma-Aldrich) in PBS. Blocking was performed in 2% BSA for 45 min. Primary antibody was diluted in 0.5% BSA and incubated for 60 min. Secondary antibody was diluted in 0.5% BSA and incubated for 60 min. Nuclei were stained using Hoechst 33342 (Life Technologies, Carlsbad CA, USA) in PBS for 1 min. Cells were mounted using gelvatol [polyvinyl acetate (PVA), Sigma-Aldrich; Cat. #P-8136; glycerol, Sigma-Aldrich; Cat. #G-9012; sodium azide, Fisher Chemical, Waltham, MA, USA; Cat. #S227-100] and refrigerated until images were captured on an Olympus Provis AX70 fluorescent microscope with Magnafire 2.1B software (Olympus, Melville, NY, USA). The following primary antibodies were used: rabbit anti-OCT3/4 (Santa Cruz sc-9081, Santa Cruz, CA, USA) 1:250, rabbit anti-SOX2 (Millipore AB5603) 1:300, rabbit anti-NANOG (Abcam ab-80892, Cambridge, MA, USA) 1:100, SSEA-4-Alexa Fluor 555 (BD Biosciences 560218) 1:100. The following secondary antibodies were used: goat anti-rabbit Alex Fluor 488 (Invitrogen) 1:500 and donkey anti-rabbit Texas Red (TR) (Santa Cruz sc-2784) 1:250. Alkaline phosphatase activity (Vector Laboratories, Burlingame, CA, USA) was measured as an indicator of pluripotency.
Teratoma Formation
hiPSCs were grown in six-well plates until about 85% confluence. The entire six-well plate was treated with 1 mg/ml dispase (Gibco), and cells were centrifuged at 200 × g for 5 min in a 50-ml conical tube (BD Biosciences). Cells were resuspended in 1 ml of mTeSR™1 and 1 ml of undiluted Matrigel, and equal volumes were transplanted subcutaneously between the scapulas of three male nonobese diabetic/severe combined immunodeficient interleukin-2 γ chain receptor knockout (NOD/SCID IL-2γ -/-) mice less than 4 months old (Jackson Laboratories, Bar Harbor, ME, USA) (dorsal injection). All animals were housed according to the University of Pittsburgh Animal Husbandry protocols. Tumors were excised and fixed with 10% formalin (Sigma-Aldrich) overnight at 4°C. Tumor sections were processed, and sections were made and stained for hematoxylin and eosin (Sigma-Aldrich).
Gene Expression
RNA was isolated using TRIzol® reagent (Life Technologies) according to the manufacturer's protocol. RNA integrity and purity were confirmed. DNase-I-treated RNA of each sample was reverse-transcribed by PCR with RT-PCR reagents (Promega, Fitchburg, WI, USA) according to the manufacturer's protocol. The resulting cDNA was analyzed by quantitative real-time PCR using specific TaqMan® assays and TaqMan® Gene Expression Master Mix (Applied Biosystems, Carlsbad, CA, USA) according to the manufacturer's protocol. Ct values were entered into the following equation to determine the arbitrary unit value 1 × 109 × e(-0.6931 × Ct) (21) and then normalized to cyclophilin A. All samples were run and analyzed on an ABI Prism 7000 Sequence Detection System. The following TaqMan® assays were used: cyclophilin A (Hs99999904_m1), OCT4 (Hs03005111_g1), SOX2 (Hs01053049_s1), NANOG (Hs02287400_g1), growth differentiation factor-3 (Gdf3; Hs00220998_m1), human telomerase reverse transcriptase (hTert; Hs00972656_m1), DNA (cytosine-5-)-methyltransferase 3 β (DNMT3B; Hs00171876_m1), C-MYC (Hs00905030_m1), KLF4 (Hs00358836_m1), Brahma/switch 2 (BRM/SWI2)-related gene 1 (Brg1 Hs00231324_m1), and BRG1-associated factor 155 (Baf155; Hs00268265_m1). The hESC line H1 was used as a positive control, and the IMR90 female fetal fibroblast line (ATCC, Manassas, VA, USA) was used as a negative control.
Flow Cytometry
After a 3- to 5-min exposure to accutase (Gibco), cells were counted, and viability was determined using the trypan blue (Gibco) exclusion assay. The cells were resuspended at a concentration of 107/ml in PBS enriched with human albumin (Sigma-Aldrich) and ethylene glycol tetraacetic acid (EGTA; Sigma-Aldrich). We characterized the cells according to their expression of nuclear and surface markers: NANOG, OCT3/4, SOX2, SSEA-3, SSEA-4, TRA1-60, and TRA1-81 (all from BD Biosciences). The staining procedure was the same for both types of markers: a minimum number of 0.5 × 106 viable cells per tube were incubated for 45 min at 4°C with appropriate amount of monoclonal antibodies directly conjugated with six fluorochromes: fluorescein isothiocyanate (FITC), phycoerythrin (PE), cyanine chrome 5 (Cy5), allophycocyanine (APC), Alexa 466 (Al.466), and Alexa 667 (Al.667) (according to manufacturer dilutions). After staining, the samples were fixed with 2% PFA for 10 min at room temperature. The evaluation of nuclear markers required a fixation–permeabilization procedure (Perm/Wash Buffer), as described in the manufacturer's protocol (BD Biosciences, San Jose, CA), before the staining procedure. Appropriate nonspecific fluorescence-conjugated antibodies of identical isotype were used as negative controls in order to evaluate positive cells for each specific antibody used. Four-color flow cytometry acquisition was performed by dual-laser FACSCalibur® equipped with CellQuest® software (BD Biosciences). To analyze and immunophenotype different cell subpopulations, at least 0.3 × 106 cells per tube were acquired. Sequential gating was implemented based on negative control staining profiles. All samples were analyzed by FlowJo® software (Tree Star Inc., Ashland, OR, USA).
Transduction Efficiency
Primary human adult and fetal hepatocytes were plated on Matrigel in six-well plates at 100,000 cells/well in their respective media and allowed to attach overnight. The following day, transduction media were given to the cells that consisted of each cell type's respective culture medium supplemented with 6 μg/ml polybrene and green fluorescent protein (GFP) lentivirus (Thermo Fisher Scientific) at MOIs of 5, 10, 25, and 50 in triplicate wells (day 0). After 24 h, the cells were washed with DMEM/F12 (Thermo Fisher Scientific) three times, and cells were given new cell culture media (day 1). On day 3, fluorescent images of the cells were taken using a Nikon Eclipse Ti inverted fluorescent microscope and Elements software (Nikon, Melville, NY, USA). Ten random areas of each well from each MOI were taken for a total of 10 pictures per MOI. Transduction efficiency was determined by the following equation: (GFP+ cells/total cells) × 100.
Toxicity Assays
Adult and fetal hepatocytes were isolated as described earlier. Ten thousand cells/well were plated on 96-well plates (BD Biosciences) coated with Matrigel in their respective cell culture media and allowed to attach overnight. The next day, each respective cell type's medium was supplemented with 6 μg/ml polybrene and a lentivirus delivering GFP at a MOI of 0–60. The next day, the cells were washed and given their respective cell culture medium. Cells for the 24- and 72-h time points were analyzed for total double-strand DNA using the Quanti-iT™ assay (Invitrogen), caspase activity by Caspase-Glo® 3/7 assay (Promega), and CellTiter-Glo® cell viability assay (Promega) according to manufacturer's instructions.
Statistical Analysis
Reprogramming efficiency and gene expression analysis were analyzed using a two-tailed Student's t test. The incidence of hESC-like colony formation was analyzed using a Fisher's exact test where colony formation in an individual experiment was scored either as a yes or a no. All toxicity assays were analyzed using a two-way ANOVA with a Bonferroni posttest. A value of p < 0.05 was determined as significant.
Results
Donor characteristics for the hepatocyte cases used in these studies are presented in Table 1. For clarity, adult hepatocytes are defined here as any hepatocytes isolated from postnatal livers. Fetal hepatocytes were isolated from tissue ranging from 89 to 168 days of gestation (Table 1). Adult and fetal hepatocytes were freshly isolated and seeded on Matrigel in their respective culture medium and allowed to attach overnight. The average viability according to trypan blue exclusion was 83% and 94% for adult and fetal cells, respectively. The cells were transduced with lentiviruses delivering the reprogramming factors for 24 h, and medium was changed every 2–3 days until hESC-like colonies appeared (see Materials and Methods). Large hESC-like colonies appeared between days 16 and 21, and individual colonies were selected for expansion between days 25 and 34 and were seeded onto new Matrigel-coated plates in mTeSR™1 medium. Fully reprogrammed colonies were easily distinguishable by their similarity to hESC colonies (Fig. 1). All selected colonies had high nuclei to cytoplasm ratio, had well-defined borders, and were otherwise morphologically indistinguishable from hESCs (Fig. 1). Three hiPSC lines were isolated from two different adult hepatocyte cases; one was from a 13-month-old patient diagnosed with CN-1 syndrome, and two lines were derived from a 12-year-old female organ donor. Attempts at reprogramming 12 other adult and pediatric cases failed to generate hiPSC colonies (n = 18 total experiments with adult hepatocytes). These cases ranged from 4 months to 39 years. In contrast, 37 hiPSC lines were isolated from five different fetal hepatocyte cases. Importantly, one line, FH1, was reprogrammed with only three factors (OCT4, SOX2, and NANOG). Attempts at reprogramming two other fetal hepatocyte cases failed to generate hiPSC colonies (n = 8 total experiments with fetal hepatocytes). Of the 40 lines generated, eight were selected and examined for characteristics of fully reprogrammed cells. Two adult (AH1 and AH-CN) and six fetal (FH1, FH2, FH12, FH14, FH15, FH29) hepatocyte hiPSC lines were further characterized for pluripotency for this report. All eight of these hiPSC lines were characterized by morphology, gene expression, flow cytometry, and ICC of pluripotency markers and teratoma formation. All other hiPSC lines established (both adult and fetal derived) were expanded and cryopreserved and banked for later use. All of the characterized cell lines were carried out to at least passage 35 and maintained characteristic hESC-like morphology and a consistent proliferation rate.

Generation of hiPSCs from adult and fetal hepatocytes in entirely feeder-free conditions. Adult or fetal hepatocytes were given lentiviruses delivering the reprogramming factors in their respective culture media overnight (day 0). The culture medium was changed to mTeSR™1 on days 1–7. The pictures are representative images of human induced pluripotent stem cell (hiPSC) lines grown on Matrigel in mTeSR™1. The hiPSC lines are morphologically indistinguishable from the H1 human embryonic stem cell (hESC) line and grow in colonies with well-defined borders and high nuclei to cytoplasm ratio. Original magnification: 100×. (A) Representative colony of fetal hepatocyte-derived hiPSC clone FH1. FH1 was derived using only octamer-binding transcription factor 4 (OCT4), sex-determining region Y box 2 (SOX2), and NANOG. (B) Representative colony of adult hepatocyte-derived hiPSC clone AH1. (C) Representative colony of adult hepatocyte-derived hiPSC clone made from cells of a patient diagnosed with Crigler–Najjar syndrome type 1 (CN-1). Clone AH-CN. (D) Representative colony of H1 hESCs. Scale bar: 100 μm.
Quantitative RT-PCR was performed using human specific TaqMan® assays. The hESC line H1 was used as a positive control, and the IMR90 fibroblast line was used as a negative control. All hiPSC lines expressed OCT4, SOX2, NANOG, GDF3, hTERT, and DNMT3B at levels comparable to H1 hESCs, although the latter two were the most variable between different hiPSC clones (Fig. 2). Flow cytometry analysis demonstrated that the hiPSC lines analyzed expressed the pluripotency surface markers SSEA-3, SSEA-4, TRA1-60, and TRA1-81 and nuclear markers OCT4, SOX2, and NANOG at levels comparable to H1 hESCs (Fig. 3). IMR90 fibroblasts were negative for all markers analyzed. The expression of TRA1-60 and 1-81 were the most variable between the different lines examined. Alkaline phosphatase activity, a property of pluripotent cells, was detected in adult and fetal-derived hiPSCs and H1 hESCs; IMR90 fibroblasts were negative (data not shown). The data generated by flow cytometry were confirmed by the analysis of the expression and localization of expressed genes characteristic of reprogrammed cells by immunofluorescence. As with the flow data, the clones expressed nuclear OCT4, SOX2, and NANOG and surface marker SSEA4, and again, IMR90 fibroblasts showed no specific reactivity (data not shown).

Gene expression analysis of pluripotency genes of hiPSC lines from adult and fetal hepatocytes. The H1 hESC line is used for a positive control, and the IMR90 fibroblast line is used as a negative control. All expression level values are normalized to cyclophilin A. All lines express the pluripotency markers OCT4, SOX2, NANOG, human telomerase reverse transcriptase (hTERT), growth differentiation factor 3 (GDF3), and DNA (cytosine-5)-methyltransferase 3β (DNMT3B).

Flow cytometry analysis of pluripotency marker expression of hiPSC lines from adult and fetal hepatocytes. The H1 hESC line is used for a positive control, and the IMR90 fibroblast line is used for a negative control. All lines express the pluripotency marker stage-specific embryonic antigen 3 (SSEA3), SSEA4, TRA1-60, TRA1-81, OCT4, SOX2, and NANOG.
Teratoma formation is considered the definitive test of complete reprogramming to pluripotency. Each hiPSC line derived from either adult or fetal hepatocytes generated complex teratomas within 8–12 weeks in NOD/SCID IL-2γ-chain receptor knockout mice. Representative pictures of structures resembling all three germ layer tissues, such as neural tissue and neural rosettes (ectoderm), chondroid tissue (mesoderm), and intestinal epithelium (endoderm) are shown in Figure 4.

Teratoma formation of selected clonally expanded hiPSC lines from both adult and fetal hepatocytes. The adult hepatocyte line depicted is AH1, and the fetal hepatocyte line depicted is FH1. The H1 hESC line is used for a positive control, and the IMR90 fibroblast line is used for a negative control (data not shown). Cell lines were subcutaneously injected into the backs of nonobese diabetic/severe combined immunodeficient interleukin-2 γ chain receptor knockout (NOD/SCID IL-2γ -/-) mice. All adult and fetal-derived hiPSC lines formed complex teratomas displaying cells from all three germ layers. FH1 and AH1 hiPSC lines display cell types ranging from neural tissue (ectoderm), chondroid tissue (mesoderm), and intestinal epithelium (endoderm). Scale bars: 50 μm.
Only three hESC-like colonies were identified from 18 experiments with 14 different donors when adult cells were used for attempted reprogramming. A total of 19,950,000 adult hepatocytes were exposed to reprogramming factors, and that resulted in three hiPSC colonies that were identified by their morphology and collected, expanded, and cryopreserved. Vastly different results were obtained when reprogramming was attempted with fetal hepatocytes. A total of 80 hESC-like colonies were identified from eight experiments with seven different donors when fetal cells were used for attempted reprogramming. A total of 4,900,000 fetal hepatocytes were exposed to reprogramming factors. Of the 80 total hiPSC colonies generated from fetal hepatocytes, 37 hiPSC colonies with ES-like morphology were selected, expanded, and cryopreserved. Reprogramming efficiency, defined as (number of hESC-like colonies formed/total number of cells exposed to the vectors) × 100, are 2.41 × 10−5 and 1.23 × 10−3 for adult and fetal hepatocytes, respectively (Fig. 5A). This represents a highly significant and over a 50-fold higher reprogramming efficiency of fetal hepatocytes compared to adults (p = 0.0015). Furthermore, 75% of the fetal hepatocyte cases yielded hESC-like colonies compared to only 11% of the adult hepatocyte isolations (Fig. 5B). The incidence, or likelihood, of hESC-like colony formation in any given experiment from adult versus fetal hepatocytes is also highly significant (Fisher's exact test with a value of p = 0.0028).
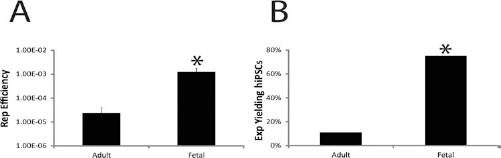
Fetal hepatocytes reprogram at over a 50-fold higher rate compared to adult hepatocytes. (A) Reprogramming (Rep) efficiency, defined as (number of hESC-like colonies formed/total number of cells exposed to the vectors) × 100, was more than 50-fold higher for fetal hepatocytes compared to adult hepatocytes. *p = 0.0015 versus adult hepatocytes, and data are SEM of n = 18 adult experiments (Exp) and n = 8 fetal experiments. (B) Fetal hepatocytes form hESC-like colonies at a higher incidence or likelihood than adult hepatocytes (Fisher's exact test). *p = 0.0028 versus adult hepatocytes. Data are SEM of n = 18 adult experiments and n = 8 fetal experiments.
Fetal cells reprogrammed with a higher efficiency than the adult cells, and so we examined the donor characteristics of the adult cases to determine if there was a consistent trend toward more efficient reprogramming in younger donors. As shown in Table 1, the cases where reprogramming was achieved were derived from two donors, one 12 years and the other 13 months of age. Reprogramming efficiency does not appear to be related to donor age in the adult cells, as the youngest “adult” donor, 4 months of age and two cases from 1-year-old donors also failed to reprogram. Many younger donors were examined, and in total, 11 of the 14 adult cases tested were from donors 12 years or under. Thus, from these 14 cases, there is no clear connection between donor age and reprogramming efficiency postbirth. However, we did not specifically design the experiment to examine reprogramming efficiency in different age groups.
A trivial explanation for such large differences in reprogramming efficiency could be related to the infectivity of the cells with the lentiviruses. To determine if there were differences between adult and fetal cells with respect to transduction efficiency, a lentivirus carrying GFP was used in place of the reprogramming vectors at a MOI ranging from 0 to 50, and the percentage of GFP-positive cells was determined 72 h after viral exposure (Fig. 6A). The percentage of GFP-positive cells increased with increasing MOI in both fetal and adult cells. The data from adult cells were consistently higher than those observed with fetal cells; however, there were no significant differences between adult and fetal cells at any MOI.

Fetal hepatocytes' higher reprogramming efficiency is not due to transduction efficiency or toxicity to reprogramming factor viruses. (A) Adult and fetal hepatocytes were exposed to lentivirus delivering green fluorescent protein (GFP) at increasing multiplicity of infection (MOI). There was no significant difference in transduction efficiency between adult and fetal hepatocytes. (B–D) The potential toxicity of lentiviral exposure was investigated at increasing MOIs and analyzed 24 h after infection. Data were normalized to MOI 0. Toxicity was measured by caspase 3/7 activity (B), ATP content (C), and total double-strand DNA (D). Data are SEM of n = 3 adult and fetal experiments. No statistical differences were seen between adult and fetal cells in the three assays. LCU, luminescent counting unit.
Large differences in reprogramming efficiency could also result if reprogramming viruses were more toxic to adult cells. To test this, a lentivirus carrying GFP was used in place of the reprogramming vectors at MOI ranging from 0 to 60. Apoptosis levels were measured by caspase 3/7 activity, and cell viability was measured by ATP content and total double-strand DNA content 24 h after viral exposure. All data were normalized to MOI 0. Adult and fetal hepatocytes showed no statistical differences between caspase 3/7 activity (Fig. 6B), cell viability measured by ATP content (Fig. 6C), or total double-strand DNA levels (Fig. 6D) at all tested MOI; however, fetal cells trended toward higher caspase activity and lower amounts of ATP. Total double-strand DNA levels were consistent between adult and fetal cells at all tested MOI. Similar data were obtained when toxicity was investigated 72 h after viral exposure (data not shown).
It was previously shown in the mouse that the increased reprogramming efficiency was associated with the endogenous expression of reprogramming factors (OCT4, SOX2, NANOG, KLF4, and C-MYC) and mediators of epigenetic changes during reprogramming (BAF complex components BRG1 and BAF155) (7). Therefore, quantitative RT-PCR was performed using human-specific TaqMan® assays to determine the endogenous levels of expression of these genes in adult and fetal hepatocytes. Adult hepatocytes expressed higher levels of endogenous OCT4, SOX2, NANOG, and KLF4, while fetal hepatocytes showed higher expression of the two BAF complex members and C-MYC (Fig. 7). Again, there were no statistical differences between expression of these genes in adult and fetal hepatocytes.

Endogenous gene expression analysis of reprogramming factors and BAF (Brg1/Brm-associated factor) complex members BAF155 and BRG1 in cultured adult or fetal hepatocytes. Expression level values are normalized to cyclophilin A. Adult and fetal hepatocytes expressed levels of all markers analyzed. Data are SEM of n = 4 adult and n = 3 fetal samples. No statistical differences were observed between adult and fetal cells. KLF, Krüppel-like factor 4; C-MYC, V-Myc myelocytomatosis viral oncogene homolog; BRG1, Brahma/switch 2 (BRM/SWI2)-related gene 1.
Discussion
Here we report the generation of 40 new hiPSC lines derived from adult hepatocytes and fetal hepatocytes. To the best of our knowledge, this is the first report of generation of human fetal hepatocyte-derived hiPSCs, the generation of a metabolic disease-specific hiPSC line from an adult hepatocyte (CN-1), and also the generation of hiPSC lines in entirely feeder-free conditions. Furthermore, in one attempt, derivation of fetal-derived hiPSCs was possible using only OCT4, SOX2, and NANOG. We did not use C-MYC, an oncogene associated with continued tumorigenicity of iPSC clones when they are differentiated from the pluripotent state.
Although initial experiments with adult hepatocytes were conducted with four reprogramming factors (n = 4), and at 5 and 10 MOI for each factor, none of these experiments yielded reprogrammed colonies. Other investigators have reported that there is an enhanced efficiency of generating hiPSCs by a combination of six factors (10). Successful reprogramming of adult cells was finally accomplished when six factors were transduced. Fetal hepatocytes were found to reprogram over 50-fold more efficiently than adult hepatocytes even though only three or four reprogramming vectors were transduced. It was recently reported that increased reprogramming efficiency was obtained with a population of cells enriched for progenitor cells (7). Even though an enriched progenitor cell population was not used in our experiments, human fetal cells at the hepatoblast stage still show over a 50-fold higher reprogramming efficiency compared to adult in this feeder-free system.
Additional experiments demonstrated that the higher reprogramming efficiency of fetal cells was not due to a more efficient lentiviral infection of fetal hepatocytes than adult hepatocytes, as adult cells were actually transduced more efficiently than fetal cells (Fig. 6A). Likewise, the differences in reprogramming efficiency were not due to the lentivirus being more toxic to the adult hepatocytes because at all tested MOI and time points the adult and fetal cells showed no statistical difference between levels of apoptosis, ATP content, and total double-strand DNA (Fig. 6B–D). In fact, the trend was the viruses were more toxic to the fetal cells.
It was recently shown that an increased reprogramming efficiency of mouse liver progenitor cells, compared with differentiated liver cells, was associated with endogenous expression of reprogramming factors and BAF complex members BAF155 and BRG1, which mediate epigenetic changes during reprogramming (7). The BAF complex components achieve a euchromatic chromatin state and enhance reprogramming efficiencies by increasing the binding of reprogramming factors onto key pluripotency gene promoters (17). However, when endogenous levels of expression of these markers in human adult and fetal hepatocytes were examined, adult hepatocytes trended to higher levels of expression of the reprogramming factors (Fig. 7), although this was not statistically significant. Interestingly, fetal hepatocytes trended to higher expression levels of the BAF complex members BAF155 and BRG1, although again, this was not statistically significant (Fig. 7). This suggests that the increased reprogramming capacity of human fetal hepatocytes in a feeder-free system is not due to higher levels of expression of the reprogramming factors or known mediators of reprogramming. Future experiments will be needed to determine if there is a molecular basis for increased reprogramming capability of human fetal hepatocytes because it is not conserved from the mouse models currently published. Increased reprogramming efficiency in fetal cells could simply be due to the observation that fetal human hepatocytes replicate robustly in culture even in the absence of added growth factors and are more likely to be simple diploid cells. In contrast, adult cells are mainly polyploid (4n, 8n, and 16n) and proliferate poorly, in vitro, which together may make it more difficult to reprogram the adult hepatocytes.
Recent evidence suggests that at low passage numbers, mouse iPSCs may retain epigenetic “memory” of the donor cell from which they were made (6) and that cell type of origin influences in vitro differentiation potential (15). This has since been corroborated in the liver when a group demonstrated hepatic lineage stage-specific donor memory to mouse iPSCs (9). More importantly, these researchers showed that mouse fetal hepatocyte-derived iPSCs retained a superior capacity for hepatic redifferentiation; however, this was lost after continuous passaging. Although not studied here, future experiments will need to determine if human adult and fetal hepatocyte-derived hiPSCs differentiate efficiently to cells with mature hepatocyte characteristics and functions.
Fetal hepatocytes have some advantages for making new hiPSC lines compared to adults. They can be generated in far fewer experiments and at a lower expenditure of time and resources. With more clones to choose from, the best lines in terms of differentiation potential could be selected. Furthermore, fetal hepatocyte tissue serves as a potential source of tissue to generate disease-specific hiPSCs, as many elective abortions are due to identification of genetic abnormalities. Moreover, the fetal cells can be induced to pluripotency without the use of oncogenes such as C-MYC. Taken together, these results demonstrate that human adult and fetal hepatocytes can be reprogrammed to pluripotency using entirely feeder-free conditions, and the efficiency of reprogramming fetal hepatocytes is more than 50-fold higher than that observed with adult cells. Although less frequent, fetal hepatocytes can be reprogrammed using only three factors, OCT4, SOX2, and NANOG. Human fetal and adult hepatocyte-derived hiPSCs may be a useful source for stem cell-derived hepatocytes that could be used for basic hepatic biology research, drug metabolism, and toxicology studies, and, possibly, as a cell source for hepatocyte transplants.
Footnotes
Acknowledgments
This work was funded, in part, by Pfizer, Inc., #N01-DK-7-0004/HHSN267200700004C and RC1-DK086135 (S.C.S.) and National Institutes of Health grant T32EB001026 (M.C.H.) and DK48794 (I.J.F). The authors declare no conflict of interest.