
Abstract
The rate of neuronal differentiation of bone marrow stromal cells (BMSCs) in vivo is very low; therefore, it is necessary to elevate the number of BMSC-derived neurons to cure neurodegenerative diseases. We previously reported that tricyclodecane-9-yl-xanthogenate (D609), an inhibitor of phosphatidylcholine-specific phospholipase C (PC-PLC), induced BMSCs to differentiate into neuron-like cells in vitro. However, the neuronal type is not clear, and it is still unknown whether these neuron-like cells possess physiological properties of functional neurons and whether they can contribute to the recovery of neuron dysfunction. To answer these questions, we investigated their characteristics by detecting neuronal function-related neurotransmitters and calcium image. The results showed that these cells exhibited functional cholinergic neurons in vitro. Transplantation of these cholinergic neuron-like cells promoted the recovery of spinal cord-injured mice, and they were more effective than BMSCs. The number of cholinergic neurons was increased after injection with BMSC-derived cholinergic neuron-like cells, indicating their high differentiation rate in vivo. Moreover, the proportion of cholinergic neurons in host cells and secretion of acetylcholine were increased, and preservation of neurofilament was also observed in the lesion of mice implanted with BMSC-derived neurons, suggesting the neuronal protection of BMSC-derived neurons. Our findings provide both a simple method to induce the differentiation of BMSCs into cholinergic neuron-like cells and a putative strategy for the therapy of spinal cord injuries.
Keywords
Introduction
Bone marrow stromal cells (BMSCs) can provide functional improvement in the central nervous system (CNS) (14); however, the mechanism by which BMSCs promote functional recovery is still in issue. It has been shown that injected BMSCs in vivo could differentiate into neurons to improve neurological behavior (26, 39). However, only a very small percentage of grafted BMSCs can differentiate into nerve cells (27). Therefore, it is urgent to elevate the differentiation rate of BMSCs into functional neurons.
Different kinds of neurons have been reported based on the release of neurotransmitters. For example: (i) Cholinergic neurons are crucial in the physiology of locomotion, cognition, and behavioral response. Loss of cholinergic neurons results in Alzheimer disease and motor neuron degeneration (37). (ii) Amino acidergic neurons, such as γ-aminobutyrinergic neurons, are located in Purkinje cells and involved in pathogenesis of Huntington's disease and neurological disorders (16). (iii) Aminergic neurons, including dopaminergic, 5-hydroxytryptaminergic, and norepinephrinergic neurons, localized in the basal ganglia and lower brain stem, and are responsible for Parkinson's disease (25), sleep disorders (42), and regulation of the sleep–wakefulness cycle (6).
We previously found that tricyclodecane-9-yl-xanthogenate (D609), the specific inhibitor of phosphatidylcholine-specific phospholipase C (PC-PLC), induced the differentiation of human, rat, and mouse BMSCs into neuron-like cells in the absence of serum (45–47). However, it was not clear which kind of neuron was induced by D609. In this article, we investigated neuronal type and function in vitro, injected these cells into spinal cord-injured mice, and investigated their effect on locomotor recovery to determine whether these BMSC-derived neurons are more advantageous for neurological repair in vivo and the reasons why these cells promote neuronal recovery.
Materials and Methods
Cell Culture and Treatment
Rat BMSCs were isolated with a modified method originally described by Pittenger et al. (34). In brief, we took marrow aspirates from male Wistar rats (Shandong University Laboratory Animal center, Jinan, China) that were 4 weeks old, then suspended and cultured them in Dulbecco's modified Eagle's medium-low glucose (DMEM-LG) (Gibco, Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and 5 ng/ml fibroblast growth factor-2 (FGF-2; Yisheng, Zhuhai, China) at 37oC in humidified air with 5% CO2. Rat BMSCs were phenotypically characterized by flow cytometry analyses published by Wang et al. (47), and the BMSCs formed a homogeneous cell population devoid of hematopoietic cells.
Fresh D609 (Sigma-Aldrich, St. Louis, MO, USA) was dissolved in water. To induce BMSCs to differentiate into neurons, we administered 4 or 8 μg/ml D609 in the DMEM-LG to BMSCs in the absence of FBS when the cells reached subconfluence, as was described in our previous article (47). The morphological changes in the cells were observed under a phase-contrast microscope.
Western Blot Analysis
Western blot and the relative protein quantity analysis were performed as described previously (15). Briefly, cells were lysed in protein lysis buffer (Beyotime, Shangai, China) and boiled for 10 min. The protein concentration was determined with the Coomassie brilliant blue (Solarbio, Beijing, China) protein assay method. BMSC protein extracts were applied to 12% sodium dodecyl sulfate (SDS)-polyacrylamide gel (Bio-Rad, Hercules, CA, USA) and transferred onto a nitrocellulose membrane (Millipore, Billerica, MA, USA). The membrane was incubated with choline acetyltransferase (ChAT), tyrosine hydroxylase (TH), dopamine β-hydroxylase (DBH), 5-hydroxytryptamine (5-HT), glutamic acid decarboxylase 65 (GAD65), anti-microtubule-associated protein 2 (MAP-2), β-actin antibodies (all from Santa Cruz Biotechnology, Santa Cruz, CA, USA) and anti-neuronal nuclei (NeuN) (Abcam, Cambridge, MA, USA) at 4°C overnight. After washing once in PBS, the membrane was incubated with appropriate horseradish peroxidase-conjugated secondary antibodies (Santa Cruz) for 1 h at room temperature. The immunoreactive bands were chromogenously developed with 3,3′-diaminobenzidine (Sangon, Shangai, China). The relative quantity of proteins was analyzed by Quantity One software (Bio-Rad) and normalized to β-actin levels.
Immunofluorescence Assay
Cells of different groups were fixed with 4% paraformaldehyde for 15 min and incubated with primary antibodies: anti-NeuN, islet 1 (ISL1), motor neuron and pancreas homeobox 1 (HB9), MAP-2 (Abcam), and synapsin (Santa Cruz). Next, the cells were washed with PBS three times and incubated with appropriate secondary antibodies (FITC-goat anti-rabbit IgG; Zhongsan Biological Technology, Beijing, China) at room temperature for 30 min, then washed with PBS three times. The samples were evaluated under a laser scanning confocal microscope (Leica, Solms, Germany). The immunofluorescence technique allows semiquantitative evaluation of protein expression (3). The relative intensity of proteins per cell was calculated as [(the intensity of cells – the intensity of background) × the area of cells]/total number of cells in random visual fields. At least 200 cells for each sample were counted.
Intracellular Calcium Measurement
Ca2+ imaging experiments were performed as described by Ciccolini (11). In brief, cells were loaded with 2 μM Fluo3 plus pluronic F127 (Invitrogen) for 30 min at room temperature (20–22°C) in an extracellular medium (EM) containing (in mM) 140 NaCl, 3 KCl, 1 MgCl2, 2 CaCl2, 10 glucose (all Sangon), 10 HEPES (Sigma-Aldrich). After incubation with Fluo3, the cells were washed with EM and incubated for a further 30 min to allow complete de-esterification of the dye. Fluorescence images (excitation at 488 nm) were monitored with a laser scanning confocal microscope (Leica), using image sizes of 256 × 256 pixels and acquisition rates of 0.5–7.5 frames/s. Neurotransmitters at a concentration of 500 μM were made in EM and delivered by bath exchange using a small volume loop injector (Yikang, Jiangxi, China; 200 μl). Ascorbic acid (Sangon; 500 μM) was added to dopamine solutions to prevent oxidation. The fluorescence changes (%ΔF/F0) for individual cells were calculated by using the formula %ΔF/F0 = (F1 – F0) × 100/F0, where F1 was the fluorescence averaged over the pixel of a cell soma after a stimulus, and F0 was the average fluorescence of that cell before stimulus application, averaged over three images. Background intensity was zero. The Ca2+ imaging data in the present study were obtained at 20–22°C.
Animals and Spinal Cord Injury (SCI) Model
Forty adult male C57BL/6J mice were obtained from the Department of Laboratory Animal Science, Peking University Health Science Center (PUHSC), China. All surgical interventions and postoperative animal care were performed in accordance with the policies established in the Guide to the Care and Use of Experimental Animals provided by the National Research Council.
The spinal cord injury model was executed as described previously (5). The adult mice (weight 20–30 g) were deeply anesthetized with an intraperitoneal injection of 3% pentobarbital sodium (Solarbio); the dorsal muscles were exposed and a laminectomy was performed on the T8-T10 level of spinal cord. Subsequently, mice received a mild contusion on T9 using an impactor (3 g × 2.5 cm). After contusion, the muscles and skin were sutured, and the animals were given postoperative antibiotics (Cisen Pharmaceutical Company, Shandong, China) and saline to prevent infection as well as dehydration. In the sham-operated mice, laminectomy was carried out without compression on spinal cord.
Cells Preparation and Transplantation
To track the injected cells in vivo, BMSCs were labeled with 5-bromo-2-deoxyuridine (BrdU, Sigma-Aldrich), which is incorporated into proliferating cells and has been utilized in many studies (50). Cells were incubated with 10 μM BrdU in medium for 48 h, and half of them were treated with 4 μg/ml D609 in serum-free DMEM and bFGF for 24 h. BrdU labeling efficiency exceeded 98% for both groups of BMSCs. For transplantation, cells were suspended in PBS to 5 × 104 cells/μl, and cell viability, estimated by trypan blue dye exclusion (Solarbio), ranged from 90% to 95%. Then, the cells were kept on ice and used within 1 h.
Seven days after spinal injury, the spinal cord was reexposed for cellular transplantation. Animals were randomly divided into three groups: (i) PBS-injected group (PBS group), (ii) transplantation of BMSCs (BMSCs group), and (iii) transplantation of BMSCs treated with D609 (D609-BMSCs group). A total of 2 × 105 cells were transplanted into the cord at the lesion site using a Hamilton syringe (Tokyo, Japan). Only animals in which the SCI was complete were included in the current analysis and were used both for behavioral tests and for histological/immunohistochemical analysis.
Assessment of Locomotor Recovery
Injured mice were tested on the following tasks after cell transplantation: Basso mouse scale (BMS) (5), footprint analysis (12), inclined plane test (33), and swim test (40, 52).
BMS: The locomotor recovery of spinal cord-injured mice was assessed by BMS, which ranges from 0 to 9, and reflects the ankle movement, plantar placement, and coordination. Mice were tested on a non-slippery surface, and the behavioral testing was first done at 24 h after cell transplantation and then, twice every week throughout the 4-week survival period.
Footprint analysis: The fore limbs and hind limbs of mice were dyed with different colors to distinguish between them, and animals were allowed to walk across a narrow, paper-covered wooden beam, 1 m in length and 4 cm wide. Stride length was measured between the central pads of two consecutive prints on each side. A series of at least eight sequential steps was used to determine the mean values. Because no steps were observed during the first week after severe injury in mice, footprint analysis was performed from the seventh day after spinal cord injury.
Inclined plane test: Mice were placed on an inclined plane, which was a 28 × 30 cm plane covered with a grooved, 1-mm-thick rubber surface. The angle of inclination was then gradually increased toward the vertical position until the mice can no longer remain in place at the starting position. The greatest height at which the mice can maintain a stable position for 5 s was recorded.
Swim test: The injured mice were gently put into a rectangular Plexiglas basin (Yikang; 150 × 40 × 13 cm), which was deep enough that the mice cannot touch the bottom of the basin. The normal animals could swim by paddling with their hind limbs and their tail, holding their forelimbs immobile under their chin. By contrast, thoracic spinal cord-injured mice prefer forelimbs for their forward movement. Thus, we counted the number of both right and left hind limb paddling for 30 s to reflect the locomotor recovery, and the average was collected.
Identification of Transplanted Cells by BrdU
To ensure cells implanted into spinal cord can be identified by BrdU, the colocalization of BrdU and BMSCs or BMSC-derived neurons was detected by immunofluorescence assay in vitro. Briefly, after pretreatment with BrdU, cells were incubated with primary mouse monoclonal anti-BrdU, rabbit polyclonal anti-neuron-specific enolase (NSE) antibody (Wuhan Boster Biological Technology, Wuhan, China), or anti-ChAT antibody (Santa Cruz). Subsequently, the appropriate FITC-conjugated or TRITC-conjugated secondary antibodies (Zhongshan Bio-Tech, Guangdong, China) were added. Propidium iodide (PI; Sigma-Aldrich) staining was utilized to label the cellular nucleus. The samples were evaluated under a laser scanning confocal microscope.
Histological Processing and Immunohistochemistry
Four weeks after transplantation, the animals were anesthetized and perfused with 4% paraformaldehyde (PFA; Sigma-Aldrich). The spinal cords including the site of contusion injury were dissected and immersed in PFA for 2 h, then embedded in optimal cutting temperature embedding medium (Tissue-Tek, Sakura Finetek, Torrance, CA, USA). Serial 7-μm-thick cryosections were collected and mounted on poly-d-lysine-coated slides (Dingguo Changsheng Biotechnology, Beijing, China). Forty to 60 sections were collected for each mouse. The maximum distance of collected spinal cords from the injury core to rostral or caudal site was 1.0 mm.
Analysis of the distribution and differentiation of the injected cells was performed with double immunofluorescence staining. Sections on slides were incubated with the primary rabbit polyclonal anti-NSE antibody or anti-ChAT antibody, then with the monoclonal anti-mouse BrdU antibody, at 4°C overnight. Appropriate FITC-conjugated or TRITC-conjugated secondary antibody was applied for 30 min at 37°C. To detect specific antibody binding, an irrelevant rabbit IgG (Santa Cruz Biotechnology), which was an isotype IgG, was used as a negative control. PI staining was utilized to label the cells in spinal cord. Sections were analyzed by laser scanning confocal microscopy.
Measurement of Neuronal Sparing
Cresyl violet and NeuN staining were utilized to assess neuronal preservation in the host spinal cords. In brief, sections were stained with cresyl violet using a staining kit (GenMed, Shanghai, China) (specific staining for neurons). Samples were observed under a phase-contrast microscope (Nikon, Tokyo, Japan) and neurons were dyed purple. The numbers of neurons in certain areas from different sections (relative number of neurons) were counted. For NeuN evaluation, sections were stained with the immunofluorescence method mentioned above. The total number of NeuN+ cells was also counted. The numbers obtained for the BMSCs or D609-BMSCs group was normalized to the PBS group.
Assessment of Axon Regeneration
Neurofilament (NF) staining was used to analyze regeneration of axons in the host spinal cord tissue according to the method of Xu et al (51). Briefly, sections of different groups were stained with primary anti-NF and FITC-conjugated secondary antibody (Santa Cruz Biotechnology) before PI staining. The samples were observed under a laser scanning confocal microscope. The number of NF+ neurons was counted and normalized to the PBS group.
D609 increased the expressions of ChAT and GAD65 in BMSCs. (A) Immunofluorescence assay showed the expression of glutamic acid decarboxylase 65 (GAD65) and choline acetyltransferase (ChAT) in the neuronal differentiation of bone marrow stromal cells (BMSCs) induced by tricyclodecane-9-yl-xanthogenate (D609). Scale bar: 40 μm. (B) Quantification of proteins shown in (A). *p < 0.05; **p < 0.01; n = 3. (C) Western blot analysis of proteins after treatment with 4 μg/ml D609 for 24 h. (D) Semiquantification of the Western blot assay. *p < 0.05; **p < 0.01; n = 5. Ctr, control; DBH, dopamine β-hydroxylase; 5-HT, 5-hydroxytryptamine; TH, tyrosine hydroxylase. ChAT and GAD65 coexisted in single neuron-like cells. The double immunofluorescence staining showed the colocalization of ChAT and GAD65 in D609-induced cells (indicated by the arrows). The magnification of the cell denoted with an asterisk in the D609 group is shown in the last panel. Scale bars: 40 μm (upper and middle panels); 8 μm (lower panel).

Assay of Acetylcholine Release
The serum of spinal cord-injured mice was collected 4 weeks after transplantation of PBS or cells, and the release of acetylcholine in serum was quantified by use of a commercially available Elisa kit (4A Biotech, Beijing, China) according to the manufacturer's instructions. The data are expressed relative to a standard curve prepared for acetylcholine.
Analysis of IL-2 and IL-10 Release
The concentrations of interleukin 2 (IL-2) and interleukin 10 (IL-10) in serum of SCI mice 4 weeks after cell transplantation were analyzed with Elisa kit (R&D, Mineapolis, MN, USA) according to the manufacturer's instructions.
Statistical Analyses
All experiments were performed in duplicate and repeated three to five times. Data are expressed as mean ± SE. SPSS 11.5 software (IBM, Armonk, New York USA) was used for statistical calculations. One-way ANOVA (followed by the Scheffé F test for post hoc analysis) was used for statistical analysis. A value of p < 0.05 was considered statistically significant.
Results
D609 Increased the Expression of ChAT and GAD65
To determine which kind of neuron-like cells were induced by D609, we examined the expressions of ChAT, GAD65, TH, DBH, and 5-HT, the specific markers of cholinergic, amino acidergic, and aminergic neurons, respectively. Both the results of immunofluorescence assay and Western blot in Figure 1 showed that after treatment with D609 for 24 h, BMSC-derived neuronal cells had strong expression of ChAT and GAD65 but not TH, DBH, or 5-HT compared with the control group. These results suggested that the neuron-like cells induced by D609 might be cholinergic or γ-aminobutyrinergic (GABAergic) neurons.
In addition, double immunofluorescence staining was performed to confirm whether the same differentiated cell express both ChAT and GAD65, or whether they give rise to mixed populations, each of which expresses a different neurotransmitter. The results showed that ChAT and GAD65 were coexpressed by most cells after D609 treatment (Fig. 2, shown by arrow), and only a small number of cells expressed one neurotransmitter (Fig. 2, indicated by an asterisk). Our results were consistent with the modified Dale's principle (18), which demonstrated the coexistence of two or more neurotransmitters within the same neuron, although the mechanism is still unclear.
D609 Induced Differentiation of BMSCs Into Cholinergic Neuron-Like Cells
To further determine whether these cells developed physiological properties of functional neurons, we performed intracellular calcium imaging in the cells (Fig. 3), which has been used to identify neuronal function in vitro (35). In the control group, there was no obvious change in Ca2+ concentration after stimulation with neurotransmitters. In contrast, increases in Ca2+ were evident in D609-treated cells after the addition of acetylcholine (Ach), a neurotransmitter of cholinergic neurons. The fluorescence intensity peaked within 10 s, quickly declined, and eventually reached the resting state (Fig. 3A, C). However, stimulation with other neurotransmitters such as γ-aminobutyric acid (GABA), l-glutamate (l-Glu), and dopamine (DA) did not induce an increase in Ca2+ concentration. These results demonstrated that the D609-induced neurons were cholinergic neuron-like cells.
Differentiated cells showed responses to exogenous acetylcholine (Ach). (A) Montage of fluorescence micrographs is shown to illustrate responses to Ach in D609-treated cells (indicated by the arrow). The image in the first panel was the cells administered with D609 under bright field, and the time (in seconds) after the application of Ach was shown at the bottom of each panel. Scale bar: 40 μm. (B, C) Relative intensity of cellular Ca2+ per cell in the control and D609-treated groups, respectively. The arrows show the time when neurotransmitter was added. GABA, γ-aminobutyric acid; l-Glu, l-glutamate; DA, dopamine; %ΔF/F0, change in fluorescence.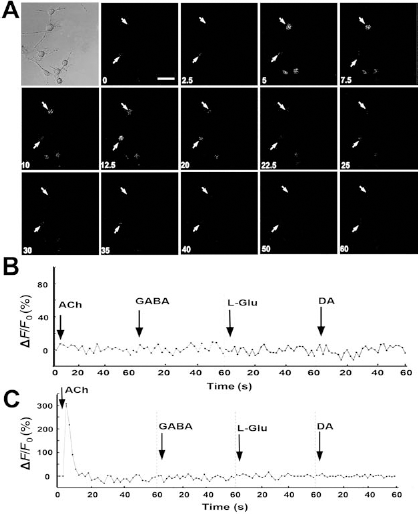
Since cholinergic neurons are frequently motor neurons, we analyzed the expression of motor neuronal markers to make sure the cells we harvested were cholinergic neurons. As shown in Figure 4, the levels of neuronal markers such as NeuN and MAP-2 were increased dramatically by D609, and immunofluorescence staining of MAP-2 and synapsin showed that cells exhibited typical dendritic morphology after D609 administration. In addition, the markers of motor neurons including ISL1 and HB9 were enhanced in the D609-treated groups. These results confirmed that D609 could induce BMSCs to differentiate into cholinergic neuron-like cells.
D609-Induced Cholinergic Neuron-Like Cells Facilitated the Recovery of SCI Mice
Cholinergic neurons are motor neurons that play pivotal roles in the transmission of signals between neurons and muscles. Spinal cholinergic neurons are affected in spinal cord injury and in diseases such as spinal muscular atrophy (17). Therefore, we investigated the influence of D609-induced cholinergic neuron-like cells on locomotor recovery in mice with spinal cord injuries.
There was no significant difference in body weight among the Sham, PBS, BMSCs and D609-BMSCs groups (data not shown). However, the behavioral indices in the four groups were diverse to a great extent (Fig. 5). The BMS score declined to zero, and severe toe dragging with no steps were observed in the PBS, BMSCs and D609-BMSCs groups after spinal cord injury, indicating that the surgery was successful. As time went on, locomotor activity in PBS-injected mice increased spontaneously and the occasional plantar stepping was detected 4 weeks after transplantation. Seven days after cell transplantation, the BMSCs and D609-BMSCs injected groups showed significant increases in their average stride lengths during stepping compared to the PBS group. Besides, the two groups also exhibited higher BMS scores, more hind limb paddling during swimming, and a longer time for holding position on the inclined plane. All the results showed that BMSCs facilitated functional recovery of SCI mice. In addition, mice that received injection of D609-induced neurons had much better behavioral indices 2 weeks after surgery compared with BMSCs group. These results showed that BMSC-derived cholinergic neuron-like cells improved functional recovery in SCI mice.
D609 increased the levels of motor neuron markers. (A) Immunofluorescence assay showed the levels of the neuronal marker neuronal nuclei (NeuN) and the markers of motor neurons including islet (ISL1) and motor neuron and pancreas homeobox 1 (HB9) in the BMSC- and D609-treated groups in vitro. (B) Quantification of proteins shown in (A). *p < 0.05; **p < 0.01; n = 4. (C) Western blot showed the bands and quantification of NeuN and microtubule-associated protein 2 (MAP-2) in different groups. **p < 0.01; n = 4. (D) MAP-2 and synapsin staining showed a typical dendritic morphology of cells after D609 treatment. Scale bars: 20 μm. Analysis of functional recovery of the mice in which PBS, BMSCs, or D609-induced neurons (D609-BMSCs) were injected. A series of behavioral tests, (A) Basso Mouse Scale (BMS), (B) footprint analysis, (C) inclined plate test, and (D) swim test, showed transplantation of D609-induced neurons exhibited better recovery of spinal cord-injured (SCI) mice. The arrow indicates the time when PBS or cells were injected into SCI mice. *p < 0.05 versus the PBS group; **p < 0.01 versus the PBS group; #p < 0.05 versus BMSCs group; ##p < 0.01 versus the BMSCs group. At least six mice per group were collected for analysis.

BMSC-Derived Cholinergic Neuron-Like Cells Promote Neural Repair
To investigate the mechanism by which BMSCs and BMSC-derived neurons promote the recovery in SCI mice, we examined the differentiation of injected cells by immunohistochemical analysis.
First, we examined the localization of BrdU in BMSCs or BMSC-derived neurons in vitro to confirm the efficiency of BrdU labeling in cells transplanted in vivo. The results in Figure 6 showed that almost all of the BMSCs could be tagged with BrdU (shown in Merge), and after induction by D609, some of these BrdU-labeled cells expressed NSE or ChAT, suggesting that BrdU did not fade away during the process of differentiation. It was notable that the majority of the NSE+ or ChAT+ cells (above 90%) (shown in Fig. 6B) were also positive for BrdU. These results indicated that NSE+BrdU+ or ChAT+BrdU+ cells could be deemed as transplanted neurons induced by D609.
Transplanted cells labeled with BrdU in vitro and the distribution of these cells in vivo. (A) Confirmation of labeling efficiency with 5-bromo-2-deoxyuridine (BrdU) in vitro. Green: NSE or ChAT, to display neuron-like cells induced by D609; red: BrdU, to show the labeled cells; blue: propidium iodide (PI), to reveal all the cells in vitro. Scale bars: 20 μm. (B) Efficiency quantification of BrdU to tag neuron-like cells in the D609 treatment group. (C) Immunohistochemical assay showing the distribution of injected cells in host spinal cords. PI-positive cells (red), cells in the intact section of spinal cord; PI+BrdU+ cells (white), injected cells. Scale bar: 200 μm.
In addition, the distribution of implanted cells in the host spinal cord was analyzed by immunofluorescence assay. As shown in Figure 6C, cells in the intact spinal cord were stained by PI (shown in red) and those injected cells (PI- and BrdU-positive cells, shown in white) localized mainly in the gray matter of spinal cords, which consist of neuronal cell bodies, neuropil (dendrites and unmyelinated axons). and glial cells (astroglia and oligodendrocytes). Therefore, in the following experiments, we chose the region of gray matter for further study.
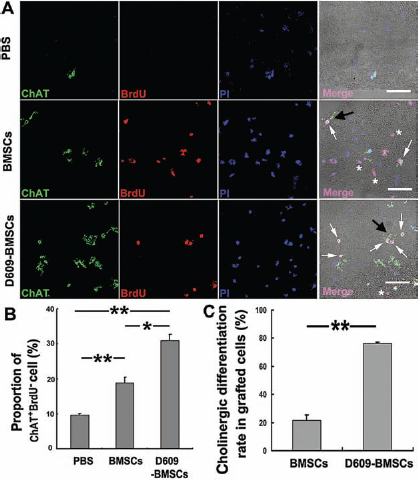
As shown in Figures 7 and 8, after being injected, only a few of the BrdU-positive BMSCs were colocalized with NSE and ChAT in the BMSCs group; most BMSCs exhibited negligible expression of NSE and ChAT (shown by asterisk). In contrast, the proportion of NSE+BrdU+ or ChAT+BrdU+ cells (shown by arrow) increased dramatically in the D609-BMSCs group, which suggested that the rate of neuronal differentiation in the D609-BMSCs group was increased significantly in comparison with the BMSCs group (Figs. 7A, C and 8A, C). Besides, some of these ChAT+BrdU+ cells exhibited neuronal dendrites (denoted by black arrow in Fig. 8A).
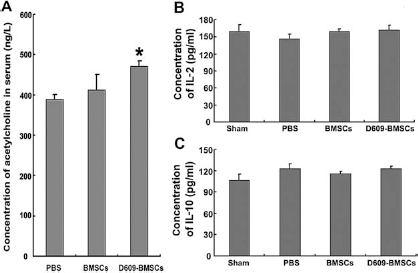
In addition, few NSE- or ChAT-positive cells were observed in the PBS group, indicating the neuronal loss after SCI. However, the number of neurons and cholinergic neurons in host cells (NSE+BrdU- and ChAT+BrdU-) was elevated in the D609-BMSCs group (Figs. 7A, B and 8A, B). Therefore, we assumed that transplantation of BMSC-derived neuron-like cells increased the number of neurons in the host spinal cords. To prove this hypothesis, cresyl violet staining and NeuN staining were performed to reveal neuronal preservation. As shown in Figure 9A–D, more neurons were present in the group that received D609-BMSCs. The results of NF staining showed NF+ neurons in host spinal cords were increased and the area and length of NF immunoreactivity were also increased in the D609-BMSCs group (Fig. 9E, green), indicating axonal regeneration promoted by these cholinergic neuron-like cells. The concentration of acetylcholine was also elevated in the serum of mice engrafted with D609-induced BMSCs (Fig. 10A). All of these results suggested that BMSC-derived cholinergic neuron-like cells could increase the number of host neural cells to promote neural recovery.
Xenotransplantation Had No Obvious Immune Rejection
Immune monitoring studies were performed to determine whether an inflammatory response to rat BMSCs was induced in the recipient mice. The levels of IL-2 and IL-10, two very important inflammatory factors, were detected in serum since they are involved in immune rejection and immune tolerance (30, 44). As shown in Figure 10B, there were no significant differences in the concentrations of IL-2 and IL-10 between the PBS and cell transplantation groups, indicating that the inflammatory response in mice was not sensitized after the transplantation.
Discussion
BMSCs represent an attractive source for cell transplantation therapies, and their curative effect has been confirmed in animal models of neurological disorders, including stroke, Parkinson's disease, spinal cord injury, and so on (10, 14). However, the mechanism by which BMSCs lead to functional recovery remains largely undefined. Some reports demonstrated that BMSCs could secrete trophic factors and cytokines to promote an endogenous repair of host neurons (23, 36), Others showed that engrafted BMSCs differentiated into neurons to improve neurological behavior (13, 31). Both of the viewpoints have been proved, and unfortunately, only a few BMSCs could differentiate into neurons in vivo. Therefore, it is necessary to develop methods to induce differentiation of BMSCs into functional neurons.
Previously, we demonstrated that inhibition of PC-PLC by D609 induced BMSCs to differentiate into neuron-like cells in the absence of serum (45–47). However, it was not clear whether the neuron-like cells were functional neurons. In the present study, we found that the neurons induced by D609 exhibited sharp responses in Ca2+ flux after the application of acetylcholine, the neurotransmitter of cholinergic neurons. It was reported that ChAT-positive cells could be harvested from BMSCs (20). However, little is known about the neuronal properties of these cells. In this study, we discovered that D609 induced BMSCs to differentiate into cholinergic neuron-like cells, which exhibited functional neuronal characteristics.
Two methods can be utilized to establish the neuronal properties of differentiated cells. The first one is the patch-clamp electrophysiology technique that reflects cellular resting and action potential, which has been adopted to test neuronal functions (24). Our previous study also showed that during the neuronal differentiation of BMSCs induced by D609, the resting membrane potential of the differentiated cells decreased, and this was similar to cultured neurons from mouse embryo forebrains (46). The second one is the intracellular calcium image method that is also widely applied to certify neuronal functions (35). Trzaska and colleagues demonstrated that during the specific differentiation of dopaminergic neurons from adult human BMSCs, low Ca2+ channels were detected and no Ca2+ image was excited after cell depolarization. These results were consistent with their inability to fire spontaneous or evoked action potentials and indicated these cells were at immature stage of dopamine neuronal progenitors (43). Also, a fast Ca2+ influx aroused by neurotransmitters also represented neuronal characteristics at physiological levels (28). Therefore, Ca2+ concentration changes stimulated by depolarization or neurotransmitters can illustrate the physiological function of neurons. In this article, we demonstrated that acetylcholine, rather than other neurotransmitters, evoked Ca2+ influx in D609-treated BMSCs, which indicated we got cholinergic neuron-like cells and eliminated the possibility of stress-initiated Ca2+ increase.
Immunohistochemical analysis of injured spinal cords in mice. (A) Neuronal differentiation of grafted cells in contused spinal cord. Grafted cells were detected by immunoreactivity against BrdU (red); PI staining indicates cells in spinal cord (blue), and NSE-positive neurons were shown in green. The merged cells of PI and BrdU (i.e., NSE negative) were shown in purple (*), while merged cells of PI, BrdU and NSE were shown in white (indicated by arrow). Scale bars: 40 μm. (B) Quantification of NSE+BrdU- cells (green only) shown in (A). *p < 0.05; **p < 0.01; n = 5; (C) Rate of neuronal differentiation from grafted cells shown in (A). **p < 0.01; n = 5. Cholinergic neuronal differentiation of injected BMSCs in spinal cord. (A) Cholinergic neuronal staining of injected BMSCs and D609-BMSCs. Injected BMSCs were labeled with BrdU (red); PI staining indicates cells in spinal cord (blue), and ChAT-positive neurons were shown in green. *, ChAT-negative BMSCs; white arrows, ChAT+BrdU+ neurons differentiated from BMSCs; black arrows, the typical dendritic morphology of injected cells. Scale bars: 20 μm. (B, C) Proportion of ChAT+BrdU- cells (green only) and cholinergic differentiation rate of grafted cells shown in (A). *p < 0.05; **p < 0.01; n = 5. Assessment of neuronal protection and axonal regeneration. (A) Results of cresyl violet staining that is specific for neurons. The neurons were dyed purple. Scale bar: 150 μm (upper panels). The magnification in different groups is shown in the lower panels. Scale bar: 75 μm. (B) The relative number of neurons was quantified in the same field of different groups. *p < 0.05; **p < 0.01; n = 4 mice, and the most representative one is presented. (C) NeuN staining of the spinal cords (upper panels, scale bar: 200 μm) and at higher magnification (lower panels, scale bar: 40 μm). (D) Relative quantification of NeuN+ cells in (C). (E) Neurofilament (NF) staining of the host spinal cords. Green: NF; red: PI. Scale bar: 40 μm. (F) Relative quantification of NF+ cells in (A). Acetylcholine release and the level of inflammatory factors in serum of spinal cord-injured mice with cell transplantation. (A) Quantification of acetylcholine in serum of different SCI mice. *p < 0.05 versus the PBS group; n = 4. (B, C) Concentration of interleukin 2 (IL-2) and IL-10, respectively, in serum of SCI mice 4 weeks after cell transplantation; n = 6.

Since the reason why BMSCs can cure neurodegenerative diseases is still an issue, in this study, we investigated the roles and neuronal differentiation of BMSCs in SCI mice. Our results showed that BMSCs could promote the recovery of SCI mice, and a few of them could differentiate into neurons including cholinergic neurons (NSE+BrdU+ or ChAT+BrdU+ cells). The data proved the low neuronal differentiation rate of BMSCs in vivo.
It has been demonstrated that delivery of BMSC-derived neurons or Schwann cells promotes functional recovery in animals with traumatized spinal tissues (21, 29, 41). These BMSC-derived neurons were induced by transgenic technology or a cocktail of induction medium such as retinoic acid/growth factors (2), β-mercaptoethanol/butylated hydroxyanisole (48), or 5-azacytidine (5-azaC)/growth factors (1). In this study, we established a more simple and effective method for producing cholinergic neuron-like cells from BMSCs rapidly, which allowed us to investigate the mechanism of BMSC neuronal differentiation by a single small molecule and to develop a new therapy method for SCI.
It was reported that BMSCs could differentiate and develop the phenotype of cholinergic neurons, and the authors predicted that these cells might be a potential source for cell therapy in spinal cord injuries (32). Our results provide direct evidence that the BMSC-derived cholinergic neuron-like cells could ameliorate symptoms in spinal cord-injured mice.
It is well known that neurons undergo atrophy in spinal cords after SCI in humans, rats, and mice, accompanied with changes in the density and morphology of neurons (7, 22, 36, 49). Delivery of BMSCs can protect against neuronal cell death to improve neuronal disorders (36, 38). In the present study, few neurons were observed after spinal cord injury, and BMSC-derived cholinergic neuron-like cell transplantation exhibited the preservation and increase of neuronal number as well as the promotion of axonal regeneration. The release of acetylcholine (neurotransmitter of cholinergic neurons) in spinal cord-injured mice was also increased by delivery of BMSC-derived cholinergic neuron-like cells, indicating the survival and maintenance of functional cholinergic neurons. We can deduce from these results that the neuronal protection of injected cells may be responsible for neurological repair. BMSCs expressed neural trophic factors and supporting substances such as nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), collagen, fibronectin, and laminin (8), and these factors activated signal transduction pathways involved in neurogenesis, angiogenesis, synaptic plasticity, and neuron restoration (9). In fact, the regenerative process after BMSC implantation was accompanied by elevated expression of neurotrophic factors at both early and later phases (8). Therefore, the effect of neuronal protection in our study might be attributed to the secretion of neurotrophic factors of BMSCs after cell transplantation.
It is inevitable that xenotransplantation can lead to death of donor cells and immune response in recipient animals. However, as one kind of poor antigen-presenting cells, BMSCs possess immune-modulatory effects and do not express major histocompatibility complex (MHC) class II, which result in their low immunogenicity and benefit their application in allografts or xenografts (4). In addition, it has been confirmed that xenotransplantation of mouse BMSCs into adult rats is feasible, and these grafted cells could survive and migrate without immunosuppression in vivo (19). Our results in this article showed that no inflammatory response was observed, and living BMSCs or BMSC-derived neurons were detected 4 weeks after transplantation, suggesting the availability of BMSC xenotransplantation.
In summary, D609 could induce BMSCs to differentiate into cholinergic neuron-like cells, and delivery of these cells promotes functional recovery and protection of neurons in spinal cord-injured mice. Our findings provide a novel strategy for obtaining cholinergic neuron-like cells from BMSCs and new possibilities for treating neurodegenerative diseases.
Footnotes
Acknowledgments
The research was financially supported by the National 973 Research Project (No. 2011CB503906), the National Natural Science Foundation of China (Nos. 90813022 and 31070735), the Natural Science Foundation of Shandong Province (ZR2010CZ004), and the Independent Innovation Foundation of Shandong University (2009JC007, 2009GN033, and 2009TS083). The authors declare no conflict of interest.