
Abstract
Adipose-derived stem cells (ADSCs) are easy to harvest and have the ability for self-renewal and to differentiate into various cell types, including those of the hepatic lineage. Studies on the use of ADSCs for liver transplantation are, however, limited. The objective of this study was to investigate the feasibility of using human ADSCs and to better understand their mechanism of action for the repair of liver damage in a thioacetamide (TAA)-induced model of chronic liver damage in the rat. To induce liver damage, 200 mg/kg TAA was injected intraperitoneally into Wistar rats every 3 days for 60 days. For cell therapy, 1 × 106 human ADSCs suspended in 300 ml of phosphate-buffered saline were transplanted into each experimental rat by direct liver injection. Immunohistochemistry showed that the transplanted ADSCs differentiated into albumin- and α-fetoprotein-secreting liver-like cells 1 week after transplantation. In addition, liver function recovered significantly, as determined by biochemical analyses that analyzed total bilirubin, prothrombin time, and albumin levels. The Metavir score, derived from histopathological analysis, also showed a significant decrease in liver fibrosis and inflammatory activity after ADSC transplantation. Finally, we found a reduction in the expression of α-smooth muscle actin, a marker of hepatic stellate cells, which produce collagen fiber, and an increase in the expression of matrix metalloproteinase-9, which degrades collagen fiber, after ADSC transplantation. These findings are consistent with abrogation of liver fibrosis in the ADSC therapy group. Consequently, these results suggest that ADSC transplantation may facilitate recovery from chronic liver damage and thus may have clinical applications.
Keywords
Introduction
Liver fibrosis is a consequence of severe liver damage and follows many forms of chronic liver damage, including viral infection, autoimmune liver disease, and sustained alcohol abuse (1). It is a chronic progressive disorder with no definitive treatment.
The main goals of liver-directed cell therapy are the replacement of diseased hepatocytes and the stimulation of endogenous or exogenous stem cell regeneration. In a rat model of liver injury, Petersen et al. first demonstrated that liver stem cells are derived from the bone marrow (BM) (16). Subsequently, BM stem cells, including mesenchymal stem cells (MSCs), were proposed as a reservoir for liver repair. Although MSCs were first identified in the BM, they can also be isolated from human umbilical cord blood, synovium, placenta, periosteum, skeletal muscle, and adipose tissue (20). Adipose-derived stem cells (ADSCs) are characterized as being similar to BM-derived MSCs. They can differentiate into neurogenic, myogenic, osteogenic, and chondrogenic lineages (24, 25). More importantly, in comparison with other types of adult stem cells, ADSCs are easily accessible and abundant. As a consequence, many experiments with ADSCs are now under way.
In a model of liver fibrosis, Kiso and colleagues (8) demonstrated a decrease in hepatic fibrosis after transplantation of ADSCs that were pretreated with basic fibroblast growth factor (bFGF) into C57BL6/J mice injured by carbon tetrachloride. Furthermore, in the rat carbon tetrachloride liver injury model, intravenous injection of ADSCs into recipient livers significantly decreases serum levels of liver enzymes and improves serum levels of albumin (11). In addition, the phosphorylation of extracellular signal-related kinases 1/2 (ERK1/2) is significantly upregulated during the hepatogenic differentiation of ADSCs (11).
We recently demonstrated that BM-derived MSCs and Wharton's jelly stem cells contribute to improved liver function of chemical-induced liver injury with progressive fibrosis (3, 12). Stem cells derived from adipose tissue are an alternative source that can be obtained by a less invasive method and in larger quantities than stem cells derived from bone marrow (9). Moreover, our previous work involved the administration of stem cells into the portal vein, which is a tedious method that requires a well-trained surgeon. These considerations led us to investigate the possibility of using ADSC transplantation to abrogate liver fibrosis in a rat model of chemical-induced liver injury. In the current study, we used naive ADSCs and direct administration of ADSCs into the thioacetamide (TAA)-induced liver. We investigated the effects of the engrafted ADSCs on the function of recipient livers. We also analyzed the migration of the implanted ADSCs and the potential of transplanted ADSCs to differentiate into hepatic-like cells. In addition, we used immunohistochemistry to examine the expression of α-smooth muscle actin (α-SMA), a marker of stellate cells, and matrix metalloproteinase-9 (MMP-9), which degrades collagen fiber, to investigate the possible mechanisms through which the transplanted ADSCs are able to abrogate liver fibrosis.
Materials and Methods
Patient ADSC Donors
Female donors who underwent gynecological surgery were enrolled in this study. The ADSC03 cell line (AD03) was derived from a 65-year-old female, and the ADSC27 cell line (AD27) was derived from a 27-year-old female. ADSCs at passages 3–5 were used in this study. The Research and Ethics Committee of China Medical University Hospital approved all procedures used in this study, and informed consent was obtained from each subject before tissue collection.
Isolation of Human ADSCs
Human adipose tissue was harvested from the subcutaneous fat (1 cm3) of the abdominal wall during gynecologic surgery. Tissue samples were placed in Ca2+/Mg2+-free phosphate-buffered saline (PBS) and immediately transferred to the laboratory. Human adipose tissue was removed from the transport medium, placed in a Petri dish, and cut into small pieces (1–2 mm3) in the presence of Ca2+/Mg2+-free PBS. The tissues were dissociated with 0.1 mg of collagenase IA (Sigma-Aldrich, St. Louis, MO) and incubated for 60 min at 37°C. After enzymatic digestion, the resultant cells were collected and cultured in knockout serum-free medium (Invitrogen-Gibco, Carlsbad, CA) supplemented with 5% fetal bovine serum (FBS; HyClone, Logan, UT), L2 ascorbic acid, and phosphate. The supernatant and debris were removed from the culture dish on day 2 of culture. The resultant ADSC culture was denoted as passage 0. To prevent spontaneous differentiation, cultures were maintained at subconfluent levels (<80% confluency). Passage of ADSC cultures was performed using 2.5% trypsin in 0.23 mM ethylenediaminetetraacetic acid. Passaged cultures were deemed passage 1. The ADSCs used in this study were primarily prepared by Dr. P.-C. Lin.
The cells were maintained in Iscove's modified Dulbecco's medium (Invitrogen-Gibco) supplemented with 10% (v/v) FBS, 10 ng/ml bFGF (R&D Systems, Minneapolis, MN), 2 mM l-glutamine, and 100 U/L penicillin–streptomycin (Invitrogen-Gibco). The cells were incubated in a 37°C incubator with 5% CO2.
Surface Phenotype Characterization
Surface markers for ADSCs were characterized using a flow cytometer (FC500; Beckman Coulter, Brea, CA) after labeling with antibodies against human cluster of differentiation 14 (CD14), CD29, CD44, CD45, and human leukocyte antigen (HLA)-ABC (all from Dako, Carpinteria, CA); CD34, CD49b, CD73, and HLA-DR (all from Becton Dickinson, Franklin Lakes, NJ). Labeling was performed according to the manufacturer's instructions.
Establishment of the Rat Model of Chronic Liver Fibrosis
To establish the rat model of chronic liver fibrosis, 200 mg/ kg TAA (Sigma-Aldrich) was injected intraperitoneally into 8-week-old male Wistar rats every 3 days for 60 days (i.e., 20 injections). In the control group, 8-week-old male Wistar rats were injected with normal saline in the same volume as the TAA group. Four days after the last injection (day 64), cardiac blood samples were collected to measure the biochemical liver function index. Rats were then sacrificed, and liver samples were obtained, processed, and sectioned to evaluate the histopathologic score.
The biochemical liver function index included measurements of glutamate oxaloacetate transaminase (GOT), glutamate pyruvate transaminase (GPT), albumin, total bilirubin, and prothrombin time. The measurements were made using a biochemical analyzer (Roche Integra 800; Roche, Holliston, MA) and were used to evaluate the extent of hepatic damage.
For histopathological analysis, liver tissue samples were fixed in 3.7% formaldehyde for 2 days. Tissues were then dehydrated, cleared, and infiltrated by a histoprocessor (Tissue-Tek; Sakura, Tokyo, Japan) for 16 h. Serial 3-μm sections of processed tissue were stained with hematoxylin and eosin Y (H&E) and Masson's trichrome stain.
For H&E histopathological analysis, sectioned samples were stained in Mayer's hematoxylin solution (Sigma-Aldrich, Steinheim, Germany) for 5 min followed by eosin Y (J.T. Baker, Deventer, Holland) for 3 min. For Masson's trichrome stain, sectioned samples were placed in Bouin's solution at 56°C for 1 h and then stained sequentially in Mayer's hematoxylin solution for 5 min, Biebrich scarlet–acid fuchsin solution for 15 min, phosphomolybdic acid–phosphotungstic acid for 15 min, and aniline blue for 5 min (all reagents from Sigma-Aldrich, Steinheim, Germany). The samples were dried and mounted onto glass slides, and sections were examined with a microscope (IX70; Olympus Tokyo, Japan). An injury grading score based on the Metavir system (17) was used to evaluate the degree of fibrosis and inflammation in the liver parenchyma (Tables 1, 2).
Metavir-Based Injury Scoring System for Liver Fibrosis

Metavir-Based Activity Score for Measuring Inflammation in the Liver Parenchyma

Cell Transplantation Experiment
After the induction of liver fibrosis (i.e., 4 days after the 20th TAA injection), the cell transplantation experiment was performed. ADSCs that had been cultured in 75-cm2 T-flasks were detached by trypsinization and collected by centrifugation (300 × g for 5 min). The cells were then resuspended in normal saline to a concentration of 2 × 106 cells/600 μl. A 300-μl aliquot subsequently was withdrawn using a 29-gauge syringe.
To prepare the experimental rats with fibrotic livers for ADSC therapy, 54 rats were injected with TAA using the aforementioned methods. After the final injection on day 60, the rats were randomly divided into three groups: ADSC03 (n = 18), ADSC27 (n = 18), and sham (n = 18). Rats in the ADSC groups were anesthetized with ether and administered 1 × 106 cells in 300 μl of normal saline by liver injection. Rats in the sham group received 300 μl of normal saline alone by liver injection.
After transplantation, six rats per day per group were euthanized on days 7, 14, and 28. Cardiac blood samples were obtained for serum biochemical analysis, and liver tissue was collected for histopathological analysis and immunohistochemical staining for human α-fetoprotein and albumin.
Immunohistochemistry
Sectioned liver samples were deparaffinized and processed by the InnoGenex™ immunohistochemistry kit (HC-2409-06; InnoGenex, San Ramon, CA). Primary antibodies against human serum albumin (1:1,000 dilution; Sigma-Aldrich), α-fetoprotein (1:20 dilution; Lab Vision, Fremont, USA), α-smooth muscle actin (1:200 dilution; Sigma-Aldrich), and matrix metalloproteinase-9 (1:200 dilution; Millipore, Billerica, MA) were used. Bound primary antibody was detected by serial addition of a biotinylated secondary antibody, horseradish peroxidase–streptavidin conjugate, and diaminobenzidine substrate. The antigen-positive area appeared brown in color under microscopic observation.
Statistical Analysis
All data are shown as the mean and standard deviation. For the comparison of different treatments between two groups, the data were analyzed using Student's t test. Values of p < 0.05 were considered significant, and values of p < 0.01 were considered highly significant. The Bonferroni correction was used for multiple comparisons.
Results
Characterization of the ADSCs
The surface phenotype of the ADSCs used in our study was similar to that of ADSCs described previously (9, 19). Figure 1 shows the phenotype of ADSC27 cells as evaluated by flow cytometry, as an example. The ADSCs obtained in this study did not express the hematopoietic cell markers CD34 or CD45, but they did express the MSC markers CD29, CD44, and CD73.

Surface phenotype and typical morphology of the human adipose-derived stem cells (ADSC27) used in this study. Flow cytometry results using selective human antibodies against (A) cluster of differentiation 14 (CD14), (B) CD29, (C) CD34, (D) CD44, (E) CD45, (F) CD49b, (G) CD73, (H) human leukocyte antigen (HLA)-DR, and (I) HLA-ABC. The percentages indicate how much the markers (shaded regions) differ from that of the isotype control (open regions).
Establishment of the Liver Fibrosis Model and Evaluation of ADSC Transplantation
To confirm the establishment of the liver fibrosis model, 200 mg/kg TAA was injected intraperitoneally into six Wistar rats every 3 days for 60 days (fibrosis group). An additional six rats received intraperitoneal injections of normal saline at the same volume as that received by the TAA group. On day 64, rats in both groups were euthanized to examine their liver function index and histopathological score. To evaluate the effects of ADSC transplantation, 54 Wistar rats were induced to develop liver fibrosis by the same method as that described above. On day 64, these rats were randomly divided into three groups of 18 rats each: ADSC03, ADSC27, and sham. The ADSC03 and ADSC27 groups received ADSC transplantation, whereas the sham group received normal saline. Six rats from each transplant group were evaluated on days 7, 14, and 28 after transplantation.
The biochemical index showed that after the TAA induction procedure, blood levels of GOT and GPT rose to relatively high levels (from 136 ± 39 to 348 ± 198 U/L and from 49 ± 6 to 101 ± 29 U/L, respectively) (Table 3), indicating that the liver cells were damaged. The prothrombin time increased significantly (from 10.97 ± 0.26 to 12.28 ± 0.78 s) (Table 3), and blood levels of albumin decreased significantly (from 4.11 ± 0.4 to 3.62 ± 0.34 g/dl) (Table 3). Decreases in serum albumin levels and increases in the prothrombin time are associated with a decrease in liver function (6); therefore, our findings indicated that the liver was injured in this model of liver fibrosis.
Biochemical Analysis of Liver Cell Function in TAA-Treated and Control Rats

p < 0.05 as compared with the normal group; six rats were analyzed for each group.
p < 0.01 as compared with the normal group; six rats were analyzed for each group. TAA, thioacetamide; GOT, glutamate oxaloacetate transaminase; GPT, glutamate pyruvate transaminase.
A histopathological analysis was also conducted using H&E staining and Masson's trichrome staining. In the liver fibrosis model (Fig. 2A), macrophage tissue infiltration was observed with H&E staining, showing that chronic TAA injections damaged the liver tissue (activity grade 4). In rats that did not receive chronic TAA injections (Fig. 2B), the liver sections appeared normal (activity grade 0). The Masson's trichrome stain showed a significant accumulation of collagen in TAA-injured liver sections, and in most areas the accumulated collagen “connected” two blood vessels, a phenomenon called bridging (15) (fibrosis grade 4) (Fig. 3A). This accumulation was not found in rats that did not receive chronic TAA injections (Fig. 3B).
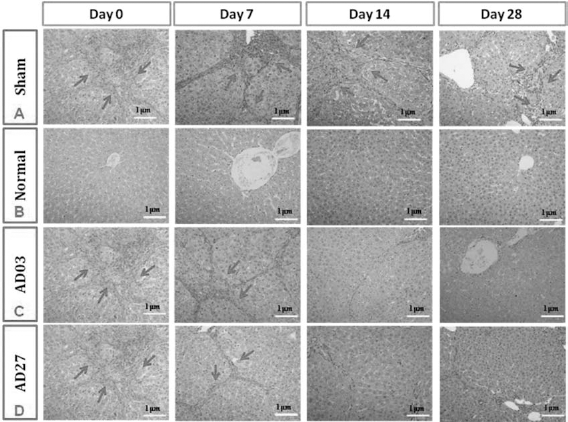
H&E staining of rat liver sections in the absence of (day 0) and 7, 14, and 28 days after ADSC transplantation. (A) Stained liver sections from a sham rat with thioacetamide (TAA)-induced liver fibrosis that received saline injections (Sham). (B) Stained liver sections from a normal rat. (C) Stained liver sections from a rat that received ADSC03 transplantation. (D) Stained liver sections from a rat that received ADSC27 transplantation. The arrows indicate that macrophage tissue infiltration was observed. Scale bars: 1 μm.

Masson's trichrome staining in liver sections in the absence of (day 0) and 7, 14, and 28 days after ADSC transplantation. (A) Stained liver sections from a sham rat with TAA-induced liver fibrosis that was injected with saline. (B) Stained liver sections from a normal rat. (C) Stained liver sections from a rat that received ADSC03 transplantation. (D) Stained liver sections from a rat that received ADSC27 transplantation. The arrows indicate where collagen has accumulated to produce the bridging phenomenon. Scale bars: 0.5 mm.
After confirmation that the liver fibrosis model was established successfully, the ADSC transplantation experiment was performed. First, the biochemical liver function indexes were compared between the cell therapy (ADSC-transplanted) groups and the sham group (Table 4). We found that the high blood levels of GOT and GPT observed after the induction of liver fibrosis decreased to near-normal values 7 days after ADSC transplantation. There were, however, no significant differences in GOT or GPT levels among the ADSC transplantation groups and the sham group at day 7. The overall low levels of GOT and GPT might reflect in part the short half-lives of GOT and GPT [i.e., 17 and 47 h, respectively (2)].
Biochemical Analysis of Liver Cell Function in TAA-Treated Rats After ADSC Transplantation

p < 0.05 as compared with the sham group; six rats were analyzed for each group.
p < 0.01 as compared with the sham group; six rats were analyzed for each group. ADSC, adipose-derived stem cells.
After 28 days of cell therapy using ADSCs, the liver function indexes showed some recovery when compared with those from the sham group (Table 4). The prothrombin time was significantly lower in the ADSC03 and ADSC27 transplantation groups as compared with that in the sham group 14 days after transplantation [10.72 ± 0.20 vs. 11.73 ± 0.67 s (p < 0.01) and 10.73 ± 0.35 vs. 11.73 ± 0.67 s (p < 0.01), respectively]. Twenty-eight days after transplantation, both of the cell transplantation groups were significantly also better than the sham group for prothrombin time. In contrast, there were no differences detected between groups in prothrombin time 7 days after transplantation. Serum albumin levels were significantly higher in the ADSC27 group than in the sham group 7 days after transplantation (3.75 ± 0.32 vs. 3.31 ± 0.33 g/dl, p < 0.05). Fourteen days after transplantation, both ADSC groups (ADSC03 and ADSC27) showed a greater recovery in serum albumin levels than did the sham group [3.67 ± 0.31 vs. 3.27 ± 0.22 g/dl (p < 0.05) and 4.05 ± 0.33 vs. 3.27 ± 0.22 g/dl (p < 0.01), respectively]. The restoration of prothrombin time and serum albumin levels suggested that 2 or 3 weeks after ADSC transplantation, functional liver mass had regenerated.
The fibrosis score that was generated (Table 1) was lower in the ADSC03 and ADSC27 transplantation groups than in the sham group 14 and 28 days after transplantation (Fig. 4). Similar findings were found when activity scores (Table 2) were examined (Fig. 5). These findings suggested that an improvement in the recovery of the damaged liver occurs 2 weeks after ADSC transplantation.

Fibrosis scores on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD scores are shown for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. ***p < 0.01, ADSC treatment group compared to the sham group.

Activity scores on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD scores are shown for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. ***p < 0.01, ADSC treatment group compared to the sham group.
The histopathological data provide further support for this suggestion (Fig. 2). Macrophage tissue infiltration was decreased in the ADSC transplantation groups on days 14 and 28 (activity score 0–1) (Fig. 2C, D), whereas the sham group still showed significant inflammation (activity score 3–4) (Figs. 2A, 5) at the same time points. Masson's trichrome stain for collagen showed almost no collagen accumulation in the ADSC transplantation groups on days 14 and 28 (fibrosis grades 1–2) (Fig. 3C, D), whereas the sham group still showed a significant amount of collagen accumulation on days 14 and 28 (fibrosis grades 3–4) (Figs. 3A, 4).
Immunostaining of Sectioned Liver Tissue
Antibodies against human α-fetoprotein and albumin were used to recognize transplanted ADSCs in sectioned liver samples and, moreover, to indicate the presence of functional differentiated hepato-lineage cells derived from ADSCs of human origin. Transplanted cells were detected up to 14 days after transplantation (Figs. 6, 7). Transplanted ADSCs were dispersed around blood vessels, and reactive sites overlapped with immunostaining for albumin. Together, these results suggested that the transplanted cells differentiate into albumin-secreting hepatocyte-like cells in the damaged livers of the experimental rats.

Immunohistochemistry using human antibodies specific for α-fetoprotein and albumin in liver sections 7 days after ADSC03 or ADSC27 transplantation. (A, C) Immunostaining for α-fetoprotein. (B, D) Immunostaining for albumin. Staining within the boxes indicates cells positive for α-fetoprotein or albumin. Scale bars: 1 mm.

Immunohistochemistry using human antibodies specific for α-fetoprotein and albumin in liver sections 14 days after ADSC03 or ADSC27 transplantation. (A, C) Immunostaining for α-fetoprotein. (B, D) Immunostaining for albumin. Staining within the boxes indicates cells positive for α-fetoprotein or albumin. Scale bars: 1 mm.
To investigate the possible mechanism through which transplanted ADSCs reduced liver fibrosis in the rat model, antibodies against α-SMA, a marker of activated stellate cells, which produce collagen, and antibodies against MMP-9, which degrades collagen, were used to stain sectioned liver samples. The expression of α-SMA was decreased in the ADSC transplantation groups when compared with the sham group at 14 days after transplantation (Fig. 8). In addition, the expression of MMP-9 was increased in the ADSC groups when compared with the sham group at 7 and 14 days after transplantation (Fig. 9). The immunostaining data suggested that ADSC transplantation reduces collagen accumulation in the liver parenchyma of rats in the liver fibrosis model.
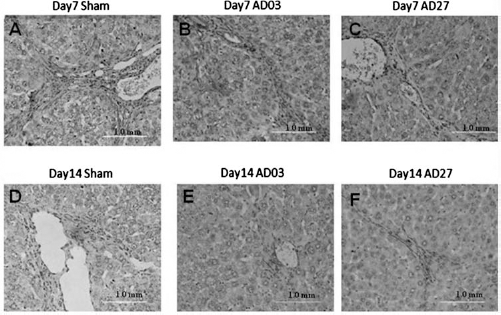
Immunohistochemistry using antibodies specific for α-smooth muscle actin (α-SMA) in liver sections 7 and 14 days after ADSC transplantation. (A, D) Immunostaining for α-SMA in liver sections from a sham rat with liver fibrosis that did not receive ADSC transplantation. (B, E) Immunostaining for α-SMA in liver sections from a rat that received ADSC03 transplantation. (C, F) Immunostaining for α-SMA in liver sections from a rat that received ADSC27 transplantation. Staining indicates cells positive for α-SMA. Scale bar: 1 mm.

Immunohistochemistry using antibodies specific for matrix metalloproteinase-9 (MMP-9) in liver sections 7 and 14 days after ADSC transplantation. (A, D) Immunostaining for MMP-9 in liver sections from a sham rat with liver fibrosis that did not receive ADSC transplantation. (B, E) Immunostaining for MMP-9 in liver sections from a rat that received ADSC03 transplantation. (C, F) Immunostaining for MMP-9 in liver sections from a rat that received ADSC27 transplantation. Staining indicates cells positive for MMP-9. Scale bar: 1 mm.
Discussion
In this study, a rat model of hepatic damage was successfully established by the periodic injection of TAA for 60 days. ADSCs isolated from the subcutaneous fat of humans differentiated into hepatocyte-like cells when injected directly into the chemically damaged liver tissue in vivo. The biochemical liver function index revealed that transplantation of human ADSCs led to the regeneration of hepatocyte-like cells, the production of albumin, a recovery of prothrombin time, a decrease in bilirubin, and a general restoration of liver function. In addition, our histopathological data and Metavir score analysis showed a substantial improvement in liver fibrosis in the ADSC transplantation groups in comparison with the sham group (Figs. 4, 5). A previous report by Liang et al. (11) on ADSC transplantation using a rat model of liver injury showed an improvement in liver function, which reached a peak 2 weeks after transplantation. In that study, the ADSCs were, however, induced before liver transplantation using a hepatogenic medium. Another study by Kamada et al. (8) showed an improvement in liver fibrosis after ADSC transplantation, but that study also involved pretreatment of ADSCs with bFGF for 1 week before transplantation. In our study, the ADSCs were naive before transplantation, and recovery of liver function was evident within 1 week after transplantation. Furthermore, in our study, ADSCs were transplanted directly into the liver, in contrast to the study by Liang et al., in which cells were injected into the hepatic portal vein, and the study by Kamada et al., in which cells were injected into the spleen. Our study involved the direct in vivo injection of naive human ADSCs into the TAA-induced fibrotic liver. Our results are the first to demonstrate that nonmanipulated transplanted ADSCs can differentiate into hepatocyte-like cells in damaged liver tissue and can ameliorate liver fibrosis by secreting metalloproteinase and decreasing the activity of stellate cells, which produce collagen fiber and thus cause liver fibrosis.
After transplantation of ADSCs into the rat liver, we estimated an average of five to eight albumin-positive cells on each high-magnification section, using 10 sections per case. We found an even distribution of albumin-positive cells throughout the liver parenchyma, not just near the injection site. Because we used a human-specific antibody against albumin, we were able to track the fate of the differentiated hepatocyte-like cells that developed from the transplanted human ADSCs. The volume of each section was calculated as 1 × 10−3 cm3 (2 cm x 1 cm x 5 × 10−4 cm), and the volume of the rat liver was calculated as 621 cm3. Therefore, the total number of albumin-positive cells in the rat liver after ADSC transplantation was estimated at 6.2 × 106 cells per rat. We transplanted 1 × 106 ADSCs per rat. Thus, the number of albumin-positive cells was six times more than our expected number. Because the immunohistochemistry was performed 1–2 weeks after ADSC transplantation, it is possible that the ADSCs proliferated. Alternatively, endogenous oval stem cells might have been induced to proliferate by the exogenous ADSCs, although this possibility is unlikely because we used human-specific antibodies against albumin and α-fetoprotein. Taken together, our study confirmed that naive ADSCs differentiate into hepatocyte-like cells and that the graft rate after direct injection into the liver is extremely efficient and is associated with no side effects.
Two human ADSC stem cell lines were used in our study: ADSC03 was derived from an older woman (65 years old), and ADSC27 was derived from a younger woman (27 years old). A comparison of the two ADSC cell lines, in terms of recovery of liver function and reduction in liver fibrosis, yielded interesting results. The two cell lines produced similar effects on serum GOT, GPT, total bilirubin, and prothrombin time (Figs. 10–12). Albumin, however, recovered earlier and persisted longer in the livers of rats that received the line that originated from the younger individual (ADSC27) than it did in those that received the line that originated from the older individual (ADSC03) (Fig. 13). Both ADSC lines produced a significant improvement in the Metavir score, a measurement of inflammation and fibrosis (Figs. 4, 5). The ADSC03 line did, however, reduce liver fibrosis to a greater extent than did the ADSC27 line (Fig. 4). In addition, the younger age-derived ADSC27 line attenuated α-SMA-positive cell generation more than the older age-derived ADSC03 line (Fig. 8), whereas the ADSC03 line increased MMP-9 activity more than the ADSC27 line (Fig. 9). It remains to be determined whether ADSCs derived from donors of different ages reduce liver fibrosis using different mechanisms.

Serum levels of glutamate oxaloacetate transaminase (GOT) (A) and glutamate pyruvate transaminase (GPT) (B) on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD levels are shown for normal rats, for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. ***p < 0.01, compared to the normal group.
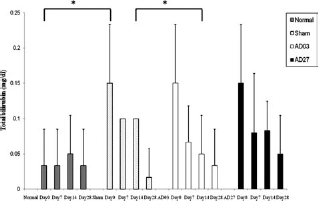
Serum levels of total bilirubin on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD levels are shown for normal rats, for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. *p < 0.05, compared to the normal or sham group.

Serum prothrombin time on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD prothrombin times are shown for normal rats, for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. *p < 0.05, compared to the sham group; ***p < 0.01, compared to the normal or sham group.

Serum levels of albumin on days 0, 7, 14, and 28 after ADSC transplantation. The mean ± SD levels are shown for normal rats, for sham rats with liver fibrosis that did not receive ADSC transplantation, for rats that received ADSC03 transplantation, and for rats that received ADSC27 transplantation. *p < 0.05, compared to the normal or sham group; ***p < 0.01, compared to the sham group.
We reported previously that transplantation of human BM stem cells and Wharton's jelly-derived stem cells promotes liver recovery after chronic liver damage (3, 12). The effect of these stem cells on the recovery of liver function was similar to that of ADSCs in terms of serum GOT, GPT, and prothrombin time. Liver recovery measured in terms of albumin was, however, more gradual after transplantation of ADSCs than after transplantation of BM- or Wharton's jelly-derived stem cells. Restoration of serum albumin levels to 3.8 g/dl required only 7 days using Wharton's jelly-derived stem cells (12), but it required 14 days using ADSC03, although both types of stem cells result in albumin recovery to the same level by 28 days after transplantation. ADSCs have several advantages over other types of stem cells, in that they are conveniently extracted from resected adipose tissue or from elective cosmetic liposuction. ADSCs also can be extracted repeatedly in large quantities in a manner that is safer and less painful than for BM stem cells (22). Therefore, ADSCs hold advantages as a source of autologous stem cells for transplantation in the treatment of liver disease.
The key cellular and molecular events involved in the pathogenesis of liver fibrosis include activation of hepatic stellate cells (HSCs) to a myofibroblast-like phenotype, production of excess matrix proteins, and increased cell proliferation (10, 23). Overproduction of extracellular matrix components, particularly collagen, is a characteristic of activated HSCs (21), and activation and proliferation of HSCs have been implicated in the pathogenesis of liver fibrosis (1, 5). Therefore, suppression of HSC activation has been proposed as a therapeutic target for the treatment of hepatic fibrosis (13). In our study, we examined the expression of α-SMA, which is a well-recognized marker of HSC activation (4), in the recipient liver after ADSC transplantation. We found that there was an obvious reduction in activated HSCs after transplantation of ADSCs into the injured liver (Fig. 8), especially with ADSC03. This result indicated that the transplanted ADSCs had a fibrolytic effect in the fibrotic liver. In contrast, a recent report showed that transplantation of ADSCs into the livers of fgf -/- mice, which do not express FGF, significantly increased liver fibrosis and that this effect was accompanied by a high expression of α-SMA in the liver parenchyma (8). In contrast, pretreatment of ADSCs with FGF before transplantation attenuated liver fibrosis (8). In our study, we used a wild-type rat, which expresses FGF. Therefore, the role of FGF in liver fibrosis cannot be determined from our study. We will examine this issue in the future.
Several reports have shown an attenuation of liver fibrosis by stem cell transplantation. Moriya et al. (14) transplanted embryonic stem cells into the fibrotic liver and found MMP-9-immunopositive cells derived from the transplanted embryonic stem cells in the recipient liver. Hardjo et al. (7) demonstrated that transplantation of MSCs improves liver fibrosis and is associated with high levels of expression of MMP-2 and MMP-9. In contrast, Roderfeld et al. (18) found an elevation in MMP-9 protein in the fibrotic liver up to 20 weeks after BM transplantation, but they identified neutrophils as the major source of MMP-9, not stem cells. In our study, there were many inflammatory cells around the central vein and portal vein of the recipient liver, concurrent with the expression of MMP-9, after ADSC transplantation, but MMP-9 expression was not increased in the absence of ADSC transplantation (Fig. 9). It remains to be determined whether ADSCs can secrete cytokines to attract inflammatory cells and thereby reduce fibrosis.
In conclusion, from the results of this study, we propose using naive ADSCs for direct administration into liver parenchymal tissue to restore liver function and ameliorate liver fibrosis. Transplantation of autologous ADSCs into the diseased liver provides a less invasive method of transplantation than do those that rely on other types of stem cells.
Footnotes
Acknowledgment
The authors declare no conflict of interest.