Glycolysis was an essential driver of chemo-resistance in colorectal cancer (CRC), albeit with limited molecular explanations.
OBJECTIVE:
We strived to elucidate the involvement of lncRNA XIST/miR-137/PKM axis in chemo-tolerance and glycolysis of CRC.
METHODS:
Altogether 212 pairs of tumor tissues and adjacent normal tissues were collected from CRC patients. Moreover, human CRC epithelial cell lines, including HT29, SW480, SW620 and LoVo, were purchased in advance, and their activity was estimated after transfection of si-XIST or miR-137 mimic. Furthermore, 5-FU/cisplatin-resistance of CRC cells was determined through MTT assay, and glycolytic potential of CRC cells was appraised based on oxygen consumption rate (OCR) and extracellular acidification rate (ECAR).
RESULTS:
Highly-expressed XIST were predictive of severe symptoms and unfavorable 3-year survival of CRC patients ( 0.05). Besides, silencing of XIST not only diminished proliferative, migratory and invasive power of CRC cells ( 0.05), but also enhanced sensitivity of CRC cells responding to 5-FU/cisplatin ( 0.05). Glycolytic potency of CRC cells was also undermined by si-XIST, with decreased maximal respiration and maximal glycolytic capacity in the si-XIST group as relative to NC group ( 0.05). Nevertheless, miR-137 mimic attenuated the facilitating effect of pcDNA3.1-XIST on proliferation, migration, invasion, 5-FU/cisplatin-resistance and glycolysis of CRC cells ( 0.05). Ultimately, ratio of PKM2 mRNA and PKM1 mRNA, despite being up-regulated by pcDNA3.1-XIST, was markedly lowered when miR-137 mimic was co-transfected ( 0.05).
CONCLUSIONS:
LncRNA XIST/miR-137 axis reinforced glycolysis and chemo-tolerance of CRC by elevating PKM2/PKM1 ratio, providing an alternative to boost chemo-therapeutic efficacy of CRC patients.
Colorectal cancer (CRC), a digestive tract malignancy, accounted for over 0.9 million deaths per year around the globe, and its annual prevalence was expected to arrive at 2.5 million by 2035 [1]. It was fortunate that 90.1% of early-stage patients were able to survive CRC for 5 years, however, merely one case out of ten patients with metastatic CRC lived longer than 5 years [2]. Despite chemotherapy-based strategies for dealing with metastatic CRC, emergence of drug resistance marginally restricted their effectiveness in hindering tumor metastasis, which ultimately led to treatment failure [3]. As proposed by the Warburg effect theory, tumor cells primarily relied on glycolysis, rather than oxidative phosphorylation, to perform metabolic activities, whether oxygen was present or not [4]. Intriguingly, this metabolic pattern was responsible for driving invasion and chemoresistance of CRC cells [5, 6], which implied that signaling pathways that disturbed glycolysis of CRC might also matter in decelerating tumor metastasis and alleviating chemoresistance in CRC.
Non-coding genes, which covered 70%–90% of the whole genome, have sparked huge interests of researchers from home and abroad, and considerable attentions were paid to their linkage with CRC etiology [7]. Taking long non-coding RNAs (lncRNAs) for instance, expressions of lncRNA MALAT1, lncRNA HOTAIR, lncRNA CCAT1 and lncRNA CRNDE were raised markedly with exacerbation of CRC, while high expressions of lncRNA MEG3 and lncRNA RP11-462C24.1 reduced the likelihood of CRC development [8, 9, 10, 11, 12, 13]. Besides that, lncRNA XIST was associated with unfavorable 6-year overall survival (OS) and progression-free survival (PFS) of CRC patients [14]. Experiments in vitro also disclosed that CRC cells were empowered to proliferate, migrate and invade under the force of XIST, and 5-FU/doxorubicin-induced resistance in CRC cells was relievable after silencing of XIST [14, 15, 16]. Nonetheless, little convincing evidence was available to explain the role of XIST in CRC glycolysis, an energy metabolism which was pivotal in chemoresistance and metastasis of tumors [5].
Based on ceRNA hypothesis, miRNAs potentially sponged by XIST were predictable through manipulation of ENCORI software [17]. It was noteworthy that a portion of the miRNAs, including miR-497, miR-195 and miR-137, were productive in hindering deterioration of CRC [18, 19]. For example, miR-137 was reported to hamper liver metastasis of CRC by prohibiting the function of IGF-1R signaling [20]. The miR-137 also showed huge potential to sensitize tumor cells (e.g. breast cancer and lung adenocarcinoma) against chemo-drugs, yet few studies were accomplished in the context of CRC [21, 22]. Furthermore, miR-137 was found to restrain glycolysis of CRC by blocking transformation of pyruvate kinase isozyme (PKM) from PKM1 to PKM2 [23, 24], a common phenomenon during tumorigenesis [25]. It might be speculated that XIST was implicated in glycolysis of CRC by acting upon miR-137 and PKM1/PKM2 transformation, yet direct proofs were in shortage.
Hence, this investigation was carried out to verify if XIST/miR-137 axis could reinforce drug resistance of CRC by hindering transformation from PKM2 to PKM1 in glycolysis, which might benefit CRC patients who received chemotherapies.
Materials and methods
Cell culture
Human CRC epithelial cell lines, including HT29, SW480, SW620 and LoVo, were supplied by cell bank of Chinese Academy of Sciences (Shanghai, China), and normal epithelial cell line (i.e. NCM460) was purchased from American Type Culture Collection (ATCC). The cell lines were cultivated in 10% FBS-containing RPMI-1640 medium (Gibco, USA) under circumstances of 5% CO, saturated humidity and constant temperature of 37C.
Cell transfection
After digestion by pancreatin (Beyotime Biotechnology, China), SW480 and LoVo cell lines were inoculated into 6-well plates at the concentration of 2 10 per well. The cell lines were, respectively, transfected by si-XIST (5’-GCUGACUACCUGAGAUUUATT-3’, Genepharma, China), si-NC (5’-UUCUCCGAACGUG UCACGUTT-3’, Genepharma, China), pcDNA-XIST (Genepharma, China), miR-137 mimic (sense: 5’-UUAUUGCUUAAGAAUACGCGUAG-3’, anti-sense: 5’-ACGCGUAAUUCUUAAGCAAUAAUU-3’, Sangon, China) and miR-NC (sense: 5’-UUCUCCGAAC GUGUCACGUTT-3’, anti-sense: 5’-ACGUAGCACG UUCGGAGAATT-3’, Sangon, China) for 48 h, according to the requirement of Lipofectamine RNAiMAX transfection kit (Invitrogen, USA).
Real-time polymerase chain reaction (PCR)
Total RNAs were harvested from CRC tissues and cell lines with the assistance of Trizol kit (Takara, Japan), and the RNAs were reversely transcribed into cDNAs utilizing reverse transcription kit (catalog number: RR047A, Takara, Japan). With primers designed by Primer 5.0 software(Supplementary Table 1), cDNAs were amplified on the real-time PCR instrument (model: CFX96, Bio-Rad, USA) in line with instructions of PCR kit (catalog number: RR820A, Takara, Japan), and relative expression of genes was calculated as per 2 method [26]. GAPDH was designated as the internal reference for XIST, PKM1 and PKM2, and U6 was set as the internal reference for miR-137.
MTT assay to assess 5-FU/cisplatin-resistance of CRC cells
SW480 and LoVo cell lines, incubated into 96-well plates at the density of 5 10/well, were firstly cultivated at 37C for 24 h, before treatment by 5-FU (Tianjin Kingyork group, China) and cisplatin (Shandong Qilu pharma, China) for 48 h. Subsequently, CRC cells of each well was incubated by 20 l MTT solution (5 mg/ml) (Sigma, USA) for 4 h, and were then mixed by 150 l dimethylsulfoxide (DMSO, Sigma, USA) in the darkness for 10 min. Optical density (OD) of each sample was measured with microplate reader (model: 680, Bio-Rad, USA) at the wavelength of 490 nm, and half maximal inhibitory concentration (IC50) of CRC cells in response to 5-FU and cisplatin were calculated employing online software of Quest Graph IC50 Calculator (https://www.aatbio.com/tools/ic50-calculator) (AAT Bioquest Inc., USA).
CCK8 assay to evaluate proliferation of CRC cells
SW480 and LoVo cells growing at the logarithmic phase were digested and were then seeded into 96-well plates at the concentration of 5 10 per well. After cultivation for 24 h, 48 h and 72 h, cell samples of each time point were treated by 10 l CCK8 reagent (DOJINDO, Japan) for 2 h. Eventually, OD values of each sample were monitored on the microplate reader (model: 680, Bio-Rad, USA) at the wavelength of 450 nm.
Transwell assay
Cell migration
In the first place, SW480 and LoVo cells were diluted by serum-free RPMI 1640 medium to a concentration of 1 10/ml. Exactly 200 l cell suspension was inoculated into the upper transwell chamber (Costar, USA), and culture medium that included FBS (10%, v/v) was supplemented into the lower transwell chamber. Twenty-four or thirty-six hours later, SW480 and LoVo cells in the upper transwell chamber were abandoned, while cells in the lower chamber were fixated by alcohol and dyed by hematoxylin. Ultimately, five views were randomly selected, and number of SW480 and LoVo cells was counted manually under the inverted microscope ( 200, Olympus, Japan).
Cell invasion
Serum-free culture medium was firstly blended with Matrigel (BD, USA) at a ratio of 8:1 (v/v), and 100 l of the mixture was paved onto the polycarbonate film of Transwell chamber. After quiescent standing for 30 min, the Matrigel was solidified. The remaining procedures were in conformity with steps of migration assay, and SW480 and LoVo cells that penetrated through the microporous membrane were counted manually under the inverted microscope (200, Olympus, Japan).
Appraisal of cell metabolism
Energy metabolism of CRC cells
SW480 and LoVo cell lines, digested to a density of 4.5 10/well, were seeded in XF-24 culture plates (Seahorse Bioscience, USA), and were then placed in an incubator of 37C and 5% CO for 24 h. Around 1 hour before detection, cells were shifted into an incubator without CO, and culture medium was replaced by bicarbonate-exclusive XF Base Medium (Seahorse Bioscience, USA) which was made up of 25 mM glucose, 1 mM pyruvic acid and 1 mM glutamine. Subsequently, 1.0 M oligomycin was added into “A” well of Seahorse gauging plate, and 1.5 M carbonyl cyanide p-trifluoromethoxyphenylhydrazone (FCCP) was supplemented into “B” well. Then mixture of rotenone and antimycin A (Rot/AA, 0.5 M) was instilled into “C” well, and oxygen consumption rate (OCR) of the CRC cells was determined [27]. In addition, detection of extracellular acidification rate (ECAR) necessitated basal glycolysis rate and acidification rate of mitochondrial aspiration after addition of 11 mM glucose. Maximal glycolytic capacity of CRC cells was recorded after supplementation of 0.5 M Rot/AA, and glycolytic reserve was determined after supplementation of 2-deoxy-D-glucose (2-DG), based on which ECAR was drawn [28].
Glycolysis of CRC cells
SW480 and LoVo cell lines were seeded into 12-well plates at the concentration of 2 10/well, and they became adherent to the plate wall after overnight cultivation. Twenty-four hours later, glucose and lactate contents within cell medium were determined with usage of glucose detection kit and lactate detection kit (both from BioAssay Systems, USA), and number of CRC cells was counted utilizing hemocytometer (model: 1492, Hausser Scientific, USA).
Dual-luciferase reporter gene assay
Wide-type pmirGLO-XIST (i.e. pmirGLO-XIST Wt) was constructed by binding pmirGLO (Promega, USA) to XIST fragments which contained binding sites with miR-137 (Genechem, China). The mutant type of pmirGLO-XIST (i.e. pmirGLO-XIST Mut) was produced almost similarly, except that XIST fragments were mutated in their binding sites with miR-137. Afterwards, SW480 and LoVo cells at the concentration of 1 10/well were cultured until they anchored to the plate wall. Then pmirGLO-XIST Wt and pmirGLO-XIST Mut were, respectively, co-transfected with miR-137 mimic or miR-NC into CRC cell lines for 48 h, as guided by the Lipofectamine2000 transfection kit (TaKaRa, Japan). After rinse by PBS, the CRC cells were dissociated by 200 l lysis buffer for 15 min, and the products were monitored using GloMax luminometer (Promega, USA).
Statistical analyses
Data of this investigation were analyzed with the support of SPSS v.18.0 software (IBM Corporation, USA). The measurement data [mean standard deviation (SD)] were compared through student’s t test or one-way analysis of variance (ANOVA), and enumeration data (n, %) were contrasted utilizing chi-square test. The comparisons were statistically significant when value was less than 0.05.
Results
Association of lncRNA XIST expression with clinico-pathological features of CRC patients
XIST expression was up-regulated in CRC tissues than in para-carcinoma tissues ( 0.05) (Supplementary Fig. 1A), and CRC cell lines, including SW480, HT-29, SW620 and LoVo, revealed higher XIST expression than normal colon epithelial cell line (i.e. NCM460) ( 0.05)(Supplementary Fig. 1B). Since that XIST expression was augmented more significantly in SW480 and LoVo cell lines than in other CRC cell lines ( 0.05), the couple of cell lines were arranged for subsequent experiments. Furthermore, CRC patients with highly-expressed ( 3.86) XIST were associated with higher odds of large tumor size ( 5 cm), deep (T3 T4) infiltration, distant metastasis and advanced TNM stage (i.e. III IV) than patients carrying lowly-expressed ( 3.86) XIST (all 0.05)(Supplementary Table 2). And highly-expressed XIST, large tumor size, poor differentiation, deep infiltration and distant metastasis were independently predictive of poor 3-year survival of the Chinese CRC cohort (all 0.05)(Supplementary Table 3, Supplementary Fig. 1C).
Silencing of lncRNA XIST undermined proliferation, migration, invasion and chemo-resistance of CRC cells
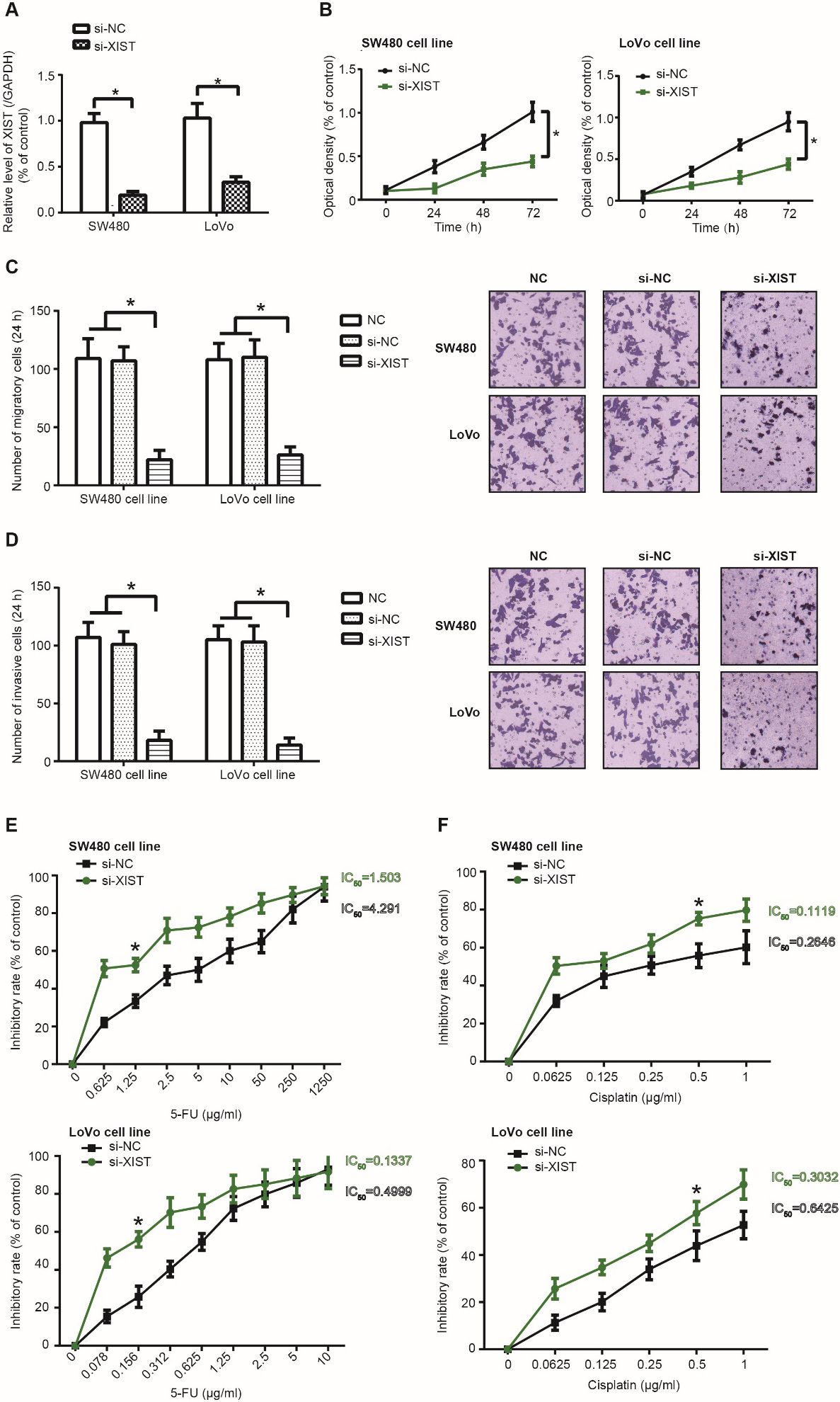
XIST expression in SW480 and LoVo cell lines was markedly reduced after transfection of si-XIST ( 0.05) (Fig. 1A), and CRC cells of si-XIST group became less capable of proliferating (Fig. 1B), migrating (Fig. 1C) and invading (Fig. 1D) than those of si-NC group (all 0.05). Furthermore, SW480 (IC50 1.503 g/ml) and LoVo cell lines (IC50 0.1337 g/ml) in the si-XIST group were vulnerable to the lethal effect of 5-FU as relative to CRC cells in the si-NC group (SW480: IC50 4.291 g/ml; LoVo: IC50 0.4999 g/ml) ( 0.05) (Fig. 1E). Cisplatin-tolerance of SW480 (IC50 0.1119 g/ml) and LoVo (IC50 0.3032 g/ml) cell lines was also abated in the si-XIST group, when compared with si-NC group (SW480: IC50 0.2646 g/ml; LoVo: IC50 0.6425 g/ml) ( 0.05) (Fig. 1F).
Activity of colorectal cancer (CRC) cells was affected by silencing of lncRNA XIST. (A) XIST expression was determined within SW480 and LoVo cell lines that were transfected by si-XIST and si-NC. *: 0.05 as relative to si-NC group. (B-D) Silencing of XIST undermined the proliferative (B), migratory (C) and invasive (D) capacity of SW480 and LoVo cell lines. *: 0.05 as relative to si-NC group. (E-F) Resistance of SW480 and LoVo cell lines against 5-FU (E) and cisplatin (F) were compared between si-XIST group and si-NC group. *: 0.05 as relative to si-NC group.
Silencing of XIST weakened glycolysis of CRC cells by increasing PKM1/PKM2 ratio
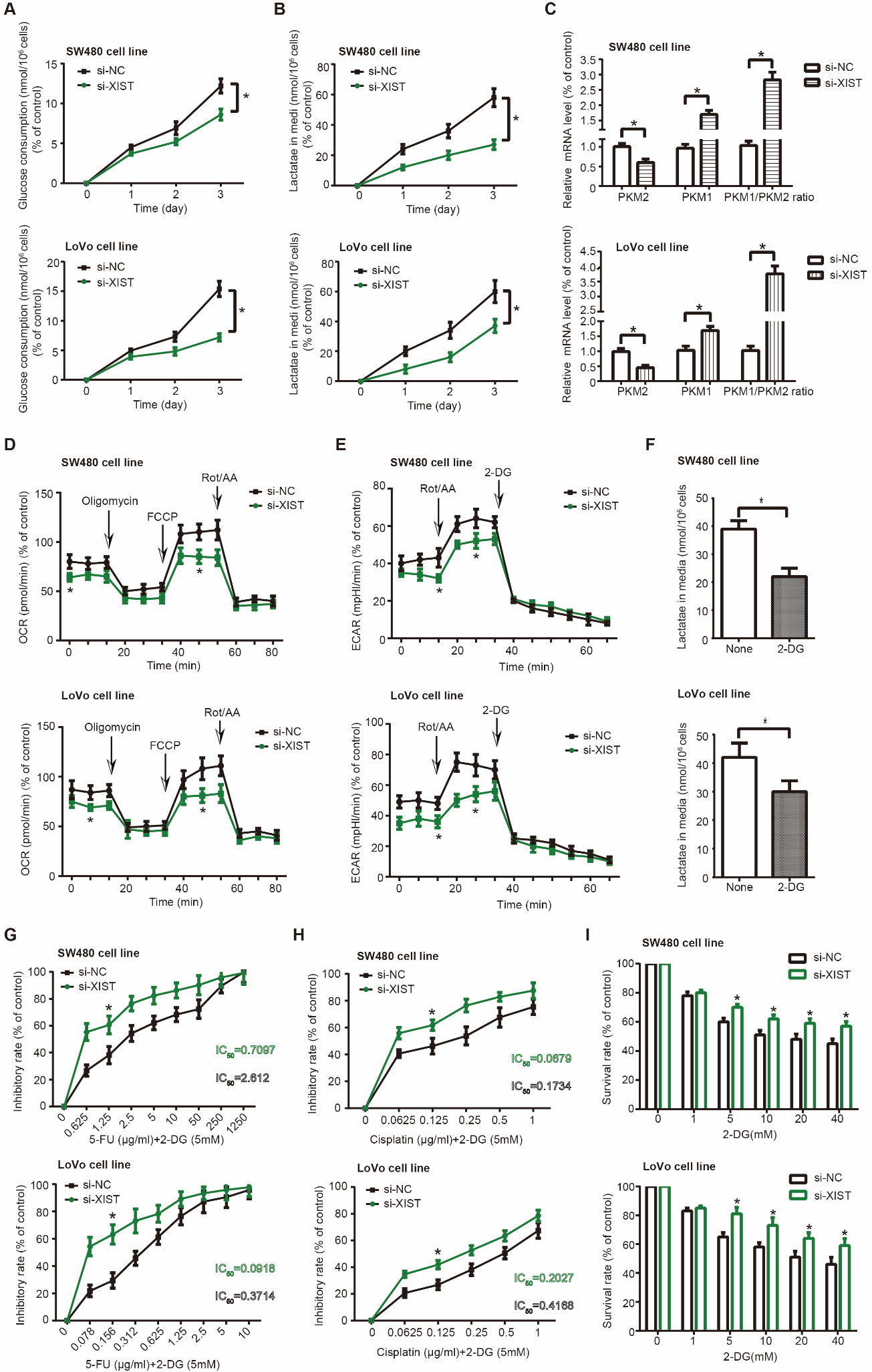
SW480 and LoVo cell lines in the si-XIST group consumed smaller amounts of glucose (Fig. 2A) and generated less lactic acid (Fig. 2B) than CRC cells of si-NC group ( 0.05). Meanwhile, mRNA level of PKM1 and PKM1/PKM2 ratio both went up significantly in the si-XIST group as compared with si-NC group ( 0.05), whereas mRNA level of PKM2 was remarkably declined after transfection of si-XIST ( 0.05) (Fig. 2C).
Glycolysis of colorectal cancer (CRC) cells was regulated by lncRNA XIST. (A-E) Glucose consumption (A), lactate production (B), PKM1/PKM2 ratio (C), oxygen consumption rate (OCR) (D) and extracellular acidification rate (ECAR) (E) of SW480 and LoVo cell lines were measured under treatments of si-XIST and si-NC. *: 0.05 as relative to si-NC group. (F) Lactate produced by SW480 and LoVo cell lines was assessed after treatment of 2-deoxy-D-glucose (2-DG). *: 0.05 as relative to NC group. (G-H) Tolerance of si-XIST-transfected SW480 and LoVo cell lines was appraised about their response to 5-FU (G) and cisplatin (H). *: 0.05 as relative to si-NC group. (I) Inhibitory effect of si-XIST on growth of SW480 and LoVo cell lines was enhanced by addition of 2-DG. *: 0.05 as relative to si-NC group.
According to Supplementary Fig. 2A, basal OCR, indicating energy need of cells under normal conditions, was comprised of ATP-linked OCR, which reflected ATP synthesis of mitochondria, and proton leak OCR, which symbolized structural damage of mitochondria [29, 30]. We noticed that basal OCR and ATP-linked OCR of SW480 and LoVo cells were lower in the si-XIST group than in the si-NC group ( 0.05) (Fig. 2D). It was implied that si-XIST could reduce basal energy consumption of CRC cells mainly by impairing ATP-synthesis capability, rather than by injuring electron transport chain (ETC) and mitochondrial membrane. More than that, maximal respiration and spare respiratory capacity also descended significantly in SW480 and LoVo cells transfected by si-XIST as relative to those transfected by si-NC ( 0.05) (Supplementary Fig. 2A, D), which indicated that si-XIST not only deactivated mitochondria of CRC cells, but also made the cells vulnerable to oxidative stress [31]. In regard to ECAR (Supplementary Fig. 2B), we found that basal glycolysis, maximal glycolytic capacity and glycolytic reserve [29] of SW480 and LoVo cells were restrained after transfection of si-XIST ( 0.05) (Fig. 2E), which insinuated that glycolytic potency of CRC cells might be attenuated by si-XIST. Additionally, lactate release of SW480 and LoVo cells was remarkably depressed in the wake of 2-deoxy-D-glucose (2-DG) treatment ( 0.05) (Fig. 2F), which was aimed to block glycolysis [32]. Treatment of 2-DG also dramatically enhanced 5-FU/cisplatin-sensitivity of SW480 and LoVo cells in both si-NC group and si-XIST group ( 0.05) (Fig. 2G and H). It was noteworthy that 5-FU/cisplatin-resistance of SW480 and LoVo cells in the si-XIST group was diminished less significantly by 2-DG treatment than cells in the si-NC group, since that the difference of IC50 value between 5-FU/cisplatin treatment and 5-FU/cisplatin 2-DG treatment was smaller in the si-XIST group than in the si-NC group(Supplementary Table 4). Similarly, survival rate of SW480 and LoVo cells in the si-XIST group was decreased less pronouncedly by 2-DG treatment than cells in the si-NC group ( 0.05) (Fig. 2I). Taken together, SW480 and LoVo cells of si-XIST group were less dependent on glycolysis than those of si-NC group.
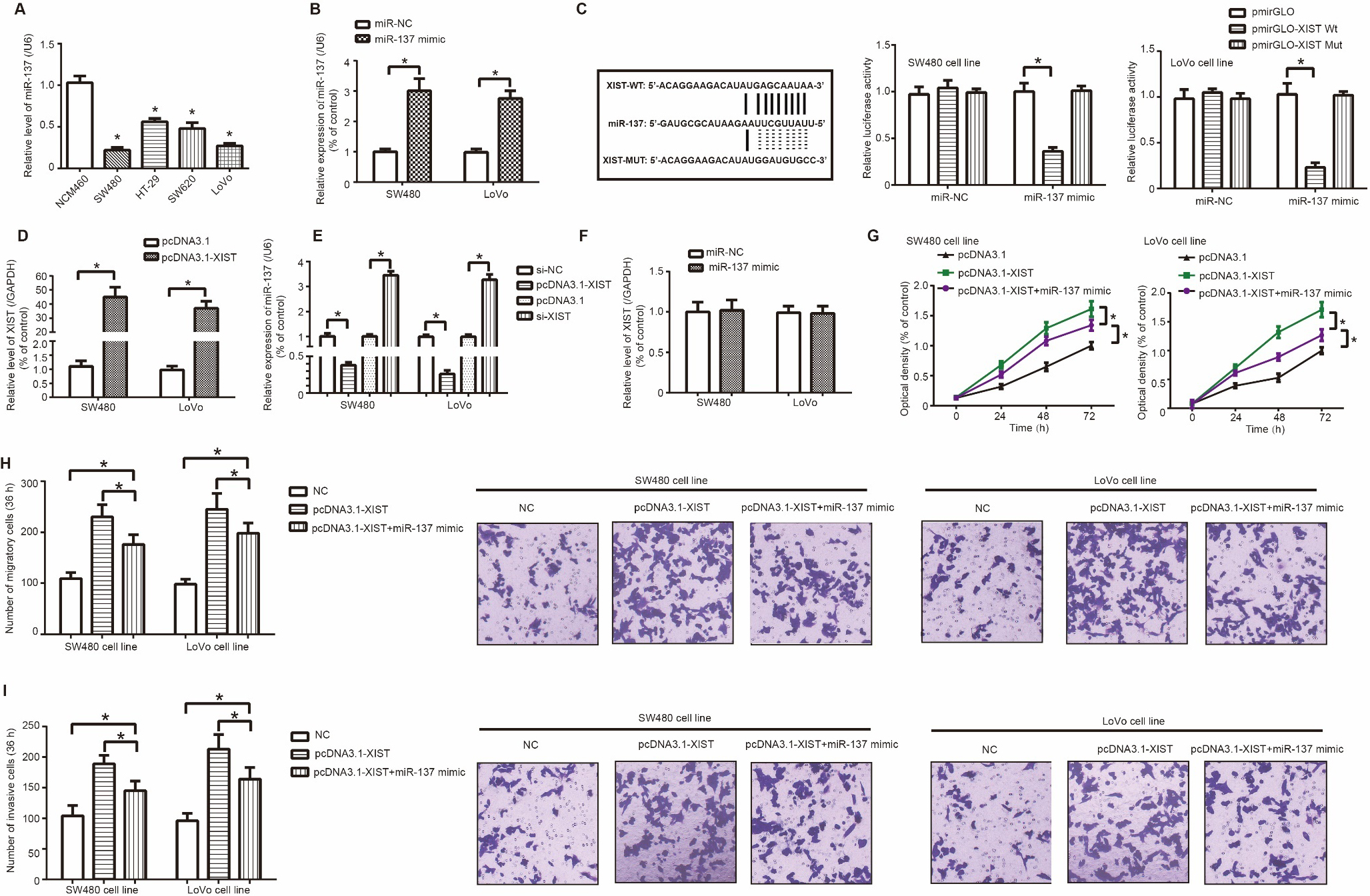
MiR-137 reversed the promoting effect of lncRNA XIST on proliferation, migration and invasion of colorectal cancer (CRC) cells. (A) Expression of miR-137 was determined among NCM460, SW480, HT-29, SW620 and LoVo cell lines. *: 0.05 as relative to NCM460 cell line. (B) MiR-137 expression within SW480 and LoVo cell lines was up-regulated after transfection of miR-137 mimic. *: 0.05 as relative to miR-NC group. (C) XIST sponged miR-137 in several sites, and luciferase activity of SW480 and LoVo cell lines was compared among XIST-WT miR-137 mimic group, XIST-MUT miR-137 mimic group and XIST-WT miR-NC group. *: 0.05 as relative to XIST-MUT miR-137 mimic group and XIST-WT miR-NC group. (D) XIST expression was promoted after transfection of pcDNA3.1-XIST. *: 0.05 as relative to pcDNA3.1 group; #: *: 0.05 as relative to si-NC group. (E) MiR-137 expression in SW480 and LoVo cell lines was altered after transfection of si-XIST and pcDNA3.1-XIST. *: 0.05 as relative to si-NC/pcDNA3.1 group. (F) XIST expression was determined when miR-137 mimic was transfected into SW480 and LoVo cell lines. *: 0.05 as relative to miR-NC group. (G-I) Proliferation (G), migration (H) and invasion (I) of SW480 and LoVo cell lines were compared among pcDNA3.1-XIST miR-137 mimic group, pcDNA3.1-XIST group and NC group. *: 0.05 as relative to NC group. #: P 0.05 as relative to pcDNA3.1-XIST group.
LncRNA XIST sponged miR-137 and depressed its expression in CRC cells
MiR-137 expression fell significantly in SW480, HT-29, SW620 and LoVo cell lines as relative to NCM460 cell line ( 0.05) (Fig. 3A), and miR-137 expression ascended significantly in SW480 and LoVo cells transfected by miR-137 mimic in comparison to miR-NC group ( 0.05) (Fig. 3B). Moreover, luciferase activity of SW480 and LoVo cells in the pmirGLO-XIST Wt miR-137 mimic group was reduced as relative to pmirGLO-XIST Mut miR-137 mimic group and pmirGLO-XIST Wt miR-NC group ( 0.05) (Fig. 3C), which verified a sponging relationship between XIST and miR-137. In addition, XIST expression in SW480 and LoVo cells was heightened after transfection of pcDNA3.1-XIST ( 0.05) (Fig. 3D). MiR-137 expression was decreased after transfection of pcDNA3.1-XIST ( 0.05), and was up-regulated when XIST was silenced ( 0.05) (Fig. 3E). Nonetheless, no statistical difference was discerned in XIST expression of SW480 and LoVo cells between miR-137 mimic group and miR-NC group ( 0.05) (Fig. 3F).
MiR-137 disturbed contribution of lncRNA XIST to proliferation, migration, invasion and chemo-resistance of CRC cells
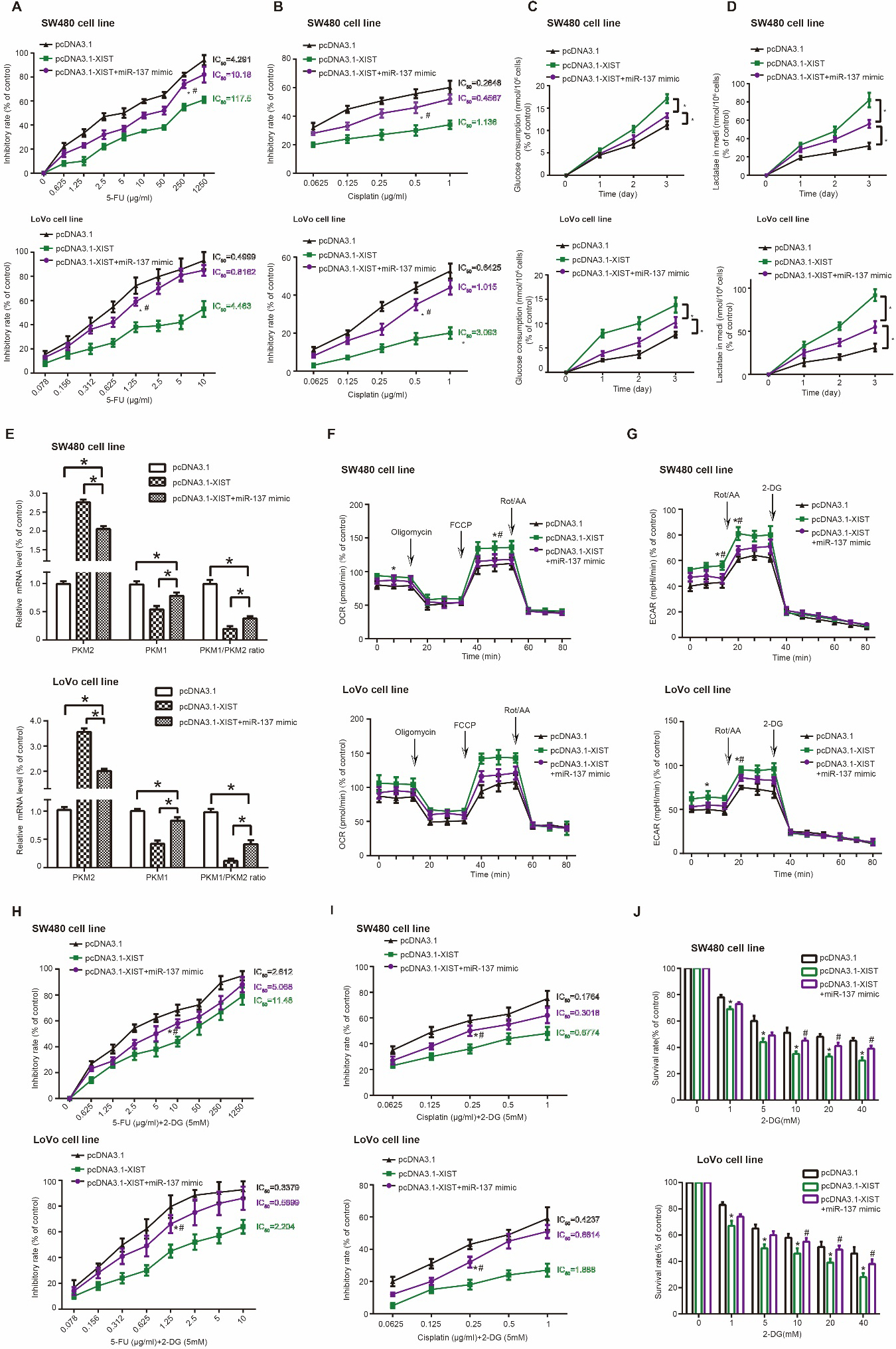
Proliferation of SW480 and LoVo cells was restrained in the pcDNA3.1-XIST miR-137 mimic group, when compared with pcDNA3.1-XIST group ( 0.05) (Fig. 3G). Analogously, migration and invasion of SW480 and LoVo cells were impaired after co-transfection of pcDNA3.1-XIST and miR-137 mimic, as relative to pcDNA3.1-XIST group ( 0.05) (Fig. 3H and I). Furthermore, SW480 (5-FU: IC50 10.18 g/ml; cisplatin: 0.4567 g/ml) and LoVo (5-FU: IC50 0.8162 g/ml; cisplatin: 1.015 g/ml) cells in the pcDNA3.1-XIST miR-137 mimic group were less tolerant to 5-FU (Fig. 4A) and cisplatin (Fig. 4B) than SW480 (5-FU: IC50 117.5 g/ml; cisplatin: 1.136 g/ml) and LoVo (5-FU: IC50 4.463 g/ml; cisplatin: 3.093 g/ml) cells of pcDNA3.1-XIST group ( 0.05).
MiR-137 obstructed XIST-motivated chemo-resistance and glycolysis in colorectal cancer (CRC) cells. (A-B) Resistance of SW480 and LoVo cell lines against 5-FU (A) and cisplatin (B) were compared among pcDNA3.1-XIST miR-137 mimic group, pcDNA3.1-XIST group and NC group. *: 0.05 as relative to NC group. #: 0.05 as relative to pcDNA3.1-XIST group. (C-G) Glucose consumption (C), lactate production (D), PKM1/PKM2 ratio (E), oxygen consumption rate (OCR) (F) and extracellular acidification rate (ECAR) (G) were monitored among SW480 and LoVo cell lines transfected by pcDNA3.1-XIST miR-137 mimic, pcDNA3.1-XIST and none. *: 0.05 as relative to NC group. #: 0.05 as relative to pcDNA3.1-XIST miR-137 mimic group. (H-J) Impacts of cisplatin 2-deoxyglucose (DG) (H), 5-FU 2-DG (I) and 2-deoxy-D-glucose (2-DG) (J) on growth of SW480 and LoVo cell lines were weighed among pcDNA3.1-XIST miR-137 mimic group, pcDNA3.1-XIST group and NC group. *: 0.05 as relative to NC group. #: 0.05 as relative to pcDNA3.1-XIST group.
MiR-137 inhibited XIST-activated glycolysis in CRC cells by decreasing PKM2/PKM1 ratio
SW480 and LoVo cells in the pcDNA3.1-XIST+miR-137 mimic group were refrained from intaking glucose (Fig. 4C) and releasing lactate (Fig. 4D), as compared with pcDNA3.1-XIST group ( 0.05). Besides, CRC cells transfected by pcDNA3.1-XIST demonstrated higher mRNA level of PKM2, lower mRNA level of PKM1 and a larger ratio of PKM2 mRNA and PKM1 mRNA than untreated CRC cells ( 0.05) (Fig. 4E). However, mRNA level of PKM2 and PKM2/PKM1 ratio were reduced, and mRNA level of PKM1 was raised obviously in the pcDNA3.1-XIST group miR-137 mimic group, when compared with pcDNA3.1-XIST group ( 0.05).
With respect to OCR and ECAR, maximal respiration and maximal glycolytic capacity of SW480 and LoVo cells were strengthened in the pcDNA3.1-XIST group as relative to NC group ( 0.05) (Fig. 4F and G, Supplementary Fig. 2). On the contrary, SW480 and LoVo cells in the pcDNA3.1-XIST miR-137 mimic group were associated with weaker maximal respiration capability and lower glycolysis-dependent ECAR than CRC cells in the pcDNA3.1-XIST group ( 0.05). Moreover, 5-FU/cisplatin-tolerance of SW480 and LoVo cells dropped significantly after 2-DG treatment (Fig. 4H and I) in comparison to without 2-DG treatment (Fig. 4A and B), and decreases in the IC50 value were less prominent in the pcDNA3.1-XIST miR-137 mimic group than in the pcDNA3.1-XIST group ( 0.05)(Supplementary Table 4). Not only that, after treatment of 2-DG, survival of SW480 and LoVo cells was depressed less significantly in the pcDNA3.1-XIST miR-137 mimic group than in the pcDNA3.1-XIST group ( 0.05) (Fig. 4J). Collectively, pcDNA3.1-XIST-enhanced reliant on glycolysis and 5-FU/cisplatin-resistance in CRC cells could be ameliorated by miR-137 mimic.
Discussion
Even if early-stage CRC was curable by surgery, massive CRC patients already exacerbated to the advanced stage at diagnosis. Huge endeavors have been made to develop chemotherapies for late-stage CRC, such as 5-FU/leucovorin combined with oxaliplatin/irinotecan [33], nevertheless, growing drug-resistance readily gave rise to failure in CRC treatment [34]. It was noteworthy that tumor cells were equipped with incremental glycolytic power [35], so elucidation of energy metabolism in CRC might be conducive to improving chemotherapeutic regimens.
LncRNAs played irreplaceable roles in a plethora of biological activities, including dynamic change of chromosome structure, genomic imprinting and dosage compensation, and their abnormal expression imposed sizable effects on malignant transformation of normal cells [36, 37]. LncRNA XIST, formed during transcriptional silencing of X chromosome, not only served as an oncogene in CRC [38, 39], but also spurred development of gastric cancer [40], nasopharyngeal carcinoma [41], non-small cell lung cancer [42] and pancreatic cancer [43]. Consistent with former documentations, we noticed that XIST encouraged proliferation, migration and invasion of CRC cells (Fig. 1B–D), which molecularly explained why high XIST expression was suggestive of CRC patients’ undesirable prognosis(Supplementary Fig. 1C). Furthermore, glycolysis of CRC cells was also explored under the influence of si-XIST (Fig. 2B–E), which was a novelty of this investigation. It was reported that glycolysis produced great deals of energy [44], and created an acidic environment made up of high-concentration lactate, which altogether geared up metastasis of tumor cells [45]. Our investigation revealed that maximal respiration (Fig. 2D), maximal glycolytic capacity (Fig. 2E), glucose uptake (Fig. 2A) and lactate release (Fig. 2B) of CRC cells were all decreased by si-XIST, which suggested that si-XIST depressed CRC progression partly by obstructing glycolysis of tumor cells [46]. Moreover, 5-FU/cisplatin-tolerance of CRC cells was also relevant to glycolysis (Fig. 2G and H), owing to that CRC cells with incremental 5-FU/cisplatin-sensitivity, triggered by si-XIST (Fig. 1E and F), depended less on glycolysis to maintain survival. Summing up the above, glycolysis blocked by si-XIST might reverse malignant phenotype and promote drug-sensitivity of CRC cells.
In addition, a sponging relationship was found to exist between XIST and miR-137 in CRC (Fig. 3C), and miR-137 could antagonize the oncogenic impact of XIST in CRC (Fig. 3H and I), which was consistent with the finding of Liu et al. [47]. Apart from CRC, miR-137 also prevented development of glioma [48], oral squamous cell carcinoma [49], gastric cancer [50] and ovarian cancer [51], implying that miR-137 might impose identical effects in diverse neoplasms. Furthermore, PKM2/PKM1 ratio, a pivotal index of glycolysis in CRC [52], was subjected to regulation of miR-137 (Fig. 4E) and XIST (Fig. 2C). There were multiple proofs that measuring PKM2 level in blood and feces was a promising approach to diagnose cancer and to track tumor progression [25]. For example, PKM2 expression was dramatically elevated with increasing severity of glioma patients [53], and patients with esophageal squamous cell carcinoma were inclined to enjoy longer life span if they carried lower PKM2 expression [54]. Not only that, phosphorylation of PKM2 could activate -catenin signaling [55] and drive expression of oncogenes in CRC (e.g. cyclin D1) [56], which persuasively expounded the CRC-promoting role of PKM2. Hence, it seemed tenable that XIST motivated glycolysis of CRC cells by suppressing the inhibitory effect of miR-137 on PKM2/PKM1 ratio (Fig. 4F–G). Last but not the least, miR-137 could diminish XIST-enhanced 5-FU/cisplatin-resistance of CRC cells (Fig. 4A and B), and miR-137 also ameliorated glycolysis-dependence of 5-FU/cisplatin-resistant CRC cells which was intensified by XIST (Fig. 4H and I). Allowing for that PKM2 was a contributor to CRC chemo-resistance [57], XIST/miR-137 axis was likely to implicate in energy metabolism and chemo-resistance of CRC by reshaping PKM2/PKM1 ratio.
In conclusion, XIST undermined the function of miR-137 and then led to biased ratio of PKM1 and PKM2, which eventually drove glycolysis and reinforced chemo-tolerance of CRC. However, several pitfalls were present in the experimental design of this investigation. Firstly, we failed to incorporate populations of different ethnicities or with distinct living habits, so whether conclusions of this study could be applied to other populations was uncertain. Secondly, rat models were not established because of technical obstacles, so that the in-vivo connection of XIST/miR-137/PKM axis with CRC glycolysis and chemo-resistance could not be validated. Finally, other glycolysis-relevant non-coding RNAs, including lncRNAs and circRNAs, should also be probed into, through which the molecular network that explained CRC etiology could be enriched.
Footnotes
Acknowledgments
This work was supported by Anhui Provincial Natural Science Foundation (No: 1808085MH240) and the Key Project of Natural Science Research of Universities of Anhui Province (No: KJ2019A0336, No: KJ2015A177).
Conflict of interest
We declare that we have no conflict of interest.
Supplementary data
The supplementary files are available to download from http://dx.doi.org/10.3233/CBM-201740.
References
1.
DekkerE.TanisP.J.VleugelsJ.L.A.KasiP.M. and WallaceM.B., Colorectal cancer, Lancet394 (2019), 1467–1480.
2.
SiegelR.DeSantisC.VirgoK.SteinK.MariottoA.SmithT.CooperD.GanslerT.LerroC.FedewaS.LinC.LeachC.CannadyR.S.ChoH.ScoppaS.HacheyM.KirchR.JemalA. and WardE., Cancer treatment and survivorship statistics, 2012, CA Cancer J Clin62 (2012), 220–241.
3.
SkarkovaV.KralovaV.VitovcovaB. and RudolfE., Selected aspects of chemoresistance mechanisms in colorectal carcinoma-a focus on epithelial-to-mesenchymal transition, autophagy, and apoptosis, Cells8 (2019).
4.
VaupelP.SchmidbergerH. and MayerA., The Warburg effect: essential part of metabolic reprogramming and central contributor to cancer progression, Int J Radiat Biol95 (2019), 912–919.
5.
WangT.NingK.SunX.ZhangC.JinL.F. and HuaD., Glycolysis is essential for chemoresistance induced by transient receptor potential channel C5 in colorectal cancer, BMC Cancer18 (2018), 207.
6.
XuH.ZengY.LiuL.GaoQ.JinS.LanQ.LaiW.LuoX.WuH.HuangY. and ChuZ., PRL-3 improves colorectal cancer cell proliferation and invasion through IL-8 mediated glycolysis metabolism, Int J Oncol51 (2017), 1271–1279.
7.
KondoY.ShinjoK. and KatsushimaK., Long non-coding RNAs as an epigenetic regulator in human cancers, Cancer Sci108 (2017), 1927–1933.
8.
WangW.XieY.ChenF.LiuX.ZhongL.L.WangH.Q. and LiQ.C., LncRNA MEG3 acts a biomarker and regulates cell functions by targeting ADAR1 in colorectal cancer, World J Gastroenterol25 (2019), 3972–3984.
9.
PanS.LiuY.LiuQ.XiaoY.LiuB.RenX.QiX.ZhouH.ZengC. and JiaL., HOTAIR/miR-326/FUT6 axis facilitates colorectal cancer progression through regulating fucosylation of CD44 via PI3K/AKT/mTOR pathway, Biochim Biophys Acta Mol Cell Res1866 (2019), 750–760.
10.
TangD.YangZ.LongF.LuoL.YangB.ZhuR.SangX. and CaoG., Inhibition of MALAT1 reduces tumor growth and metastasis and promotes drug sensitivity in colorectal cancer, Cell Signal57 (2019), 21–28.
11.
DingJ.LiJ.WangH.TianY.XieM.HeX.JiH.MaZ.HuiB.WangK. and JiG., Long noncoding RNA CRNDE promotes colorectal cancer cell proliferation via epigenetically silencing DUSP5/CDKN1A expression, Cell Death Dis8 (2017), e2997.
12.
LvL.JiaJ.Q. and ChenJ., The lncRNA CCAT1 Upregulates Proliferation and Invasion in Melanoma Cells via Suppressing miR-33a, Oncol Res26 (2018), 201–208.
13.
WangC.YuJ.HanY.LiL.LiJ.LiT. and QiP., Long non-coding RNAs LOC285194, RP11-462C24.1 and Nbla12061 in serum provide a new approach for distinguishing patients with colorectal cancer from healthy controls, Oncotarget7 (2016), 70769–70778.
14.
XiaoY.YurievichU.A. and YosypovychS.V., Long noncoding RNA XIST is a prognostic factor in colorectal cancer and inhibits 5-fluorouracil-induced cell cytotoxicity through promoting thymidylate synthase expression, Oncotarget8 (2017), 83171–83182.
15.
YangL.G.CaoM.Z.ZhangJ.LiX.Y. and SunQ.L., LncRNA XIST modulates HIF-1A/AXL signaling pathway by inhibiting miR-93-5p in colorectal cancer, Mol Genet Genomic Med8 (2020), e1112.
16.
ZhuJ.ZhangR.YangD.LiJ.YanX.JinK.LiW.LiuX.ZhaoJ.ShangW. and YuT., Knockdown of long non-coding RNA XIST Inhibited doxorubicin resistance in colorectal cancer by upregulation of miR-124 and downregulation of SGK1, Cell Physiol Biochem51 (2018), 113–128.
17.
LiJ.H.LiuS.ZhouH.QuL.H. and YangJ.H., starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data, Nucleic Acids Res42 (2014), D92–97.
18.
PoelD.BoydL.N.C.BeekhofR.SchelfhorstT.PhamT.V.PiersmaS.R.KnolJ.C.JimenezC.R.VerheulH.M.W. and BuffartT.E., Proteomic Analysis of miR-195 and miR-497 Replacement Reveals Potential Candidates that Increase Sensitivity to Oxaliplatin in MSI/P53wt Colorectal Cancer Cells, Cells8 (2019),
19.
ZouG.WangR. and WangM., Clinical response and prognostic significance of serum miR-497 expression in colorectal cancer, Cancer Biomark25 (2019), 11–18.
20.
ChuP.C.LinP.C.WuH.Y.LinK.T.WuC.Bekaii-SaabT.LinY.J.LeeC.T.LeeJ.C. and ChenC.S., Mutant KRAS promotes liver metastasis of colorectal cancer, in part, by upregulating the MEK-Sp1-DNMT1-miR-137-YB-1-IGF-IR signaling pathway, Oncogene37 (2018), 3440–3455.
21.
ChengS.HuangY.LouC.HeY.ZhangY. and ZhangQ., FSTL1 enhances chemoresistance and maintains stemness in breast cancer cells via integrin beta3/Wnt signaling under miR-137 regulation, Cancer Biol Ther20 (2019), 328–337.
22.
SuT.J.KuW.H.ChenH.Y.HsuY.C.HongQ.S.ChangG.C.YuS.L. and ChenJ.J., Oncogenic miR-137 contributes to cisplatin resistance via repressing CASP3 in lung adenocarcinoma, Am J Cancer Res6 (2016), 1317–1330.
23.
SunY.ZhaoX.ZhouY. and HuY., miR-124, miR-137 and miR-340 regulate colorectal cancer growth via inhibition of the Warburg effect, Oncol Rep28 (2012), 1346–1352.
24.
GuZ.XiaJ.XuH.FrechI.TricotG. and ZhanF., NEK2 promotes aerobic glycolysis in multiple myeloma through regulating splicing of pyruvate kinase, J Hematol Oncol10 (2017), 17.
25.
LiY.H.LiX.F.LiuJ.T.WangH.FanL.L.LiJ. and SunG.P., PKM2, a potential target for regulating cancer, Gene668 (2018), 48–53.
26.
LivakK.J. and SchmittgenT.D., Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method, Methods25 (2001), 402–408.
27.
ZaytsevaY.Y.HarrisJ.W.MitovM.I.KimJ.T.ButterfieldD.A.LeeE.Y.WeissH.L.GaoT. and EversB.M., Increased expression of fatty acid synthase provides a survival advantage to colorectal cancer cells via upregulation of cellular respiration, Oncotarget6 (2015), 18891–18904.
28.
LiQ.TangH.HuF. and QinC., Silencing of FOXO6 inhibits the proliferation, invasion, and glycolysis in colorectal cancer cells, J Cell Biochem120 (2019), 3853–3860.
29.
YangM.ChadwickA.E.DartC.KamishimaT. and QuayleJ.M., Bioenergetic profile of human coronary artery smooth muscle cells and effect of metabolic intervention, PLoS One12 (2017), e0177951.
30.
BaddingM.A.FixN.R.AntoniniJ.M. and LeonardS.S., A comparison of cytotoxicity and oxidative stress from welding fumes generated with a new nickel-, copper-based consumable versus mild and stainless steel-based welding in RAW 264.7 mouse macrophages, PLoS One9 (2014), e101310.
31.
HillB.G.BenavidesG.A.LancasterJ.R., Jr.BallingerS.Dell’ItaliaL.JianhuaZ. and Darley-UsmarV.M., Integration of cellular bioenergetics with mitochondrial quality control and autophagy, Biol Chem393 (2012), 1485–1512.
32.
O’NeillS.PorterR.K.McNameeN.MartinezV.G. and O’DriscollL., 2-Deoxy-D-Glucose inhibits aggressive triple-negative breast cancer cells by targeting glycolysis and the cancer stem cell phenotype, Sci Rep9 (2019), 3788.
33.
Garcia-AlfonsoP.TorresG.GarciaG.GallegoI.OrtegaL.SandovalC.MunozA. and LloansiA., FOLFOXIRI plus biologics in advanced colorectal cancer, Expert Opin Biol Ther19 (2019), 411–422.
34.
FongW. and ToK.K.W., Drug repurposing to overcome resistance to various therapies for colorectal cancer, Cell Mol Life Sci76 (2019), 3383–3406.
35.
ChakrabortyP.K.MustafiS.B.XiongX.DwivediS.K.D.NesinV.SahaS.ZhangM.DhanasekaranD.JayaramanM.MannelR.MooreK.McMeekinS.YangD.ZunaR.DingK.TsiokasL.BhattacharyaR. and MukherjeeP., MICU1 drives glycolysis and chemoresistance in ovarian cancer, Nat Commun8 (2017), 14634.
36.
SunJ.DingC.YangZ.LiuT.ZhangX.ZhaoC. and WangJ., The long non-coding RNA TUG1 indicates a poor prognosis for colorectal cancer and promotes metastasis by affecting epithelial-mesenchymal transition, J Transl Med14 (2016), 42.
37.
YangL.QiuM.XuY.WangJ.ZhengY.LiM.XuL. and YinR., Upregulation of long non-coding RNA PRNCR1 in colorectal cancer promotes cell proliferation and cell cycle progression, Oncol Rep35 (2016), 318–324.
38.
LiuA.LiuL. and LuH., LncRNA XIST facilitates proliferation and epithelial-mesenchymal transition of colorectal cancer cells through targeting miR-486-5p and promoting neuropilin-2, J Cell Physiol234 (2019), 13747–13761.
39.
SongH.HeP.ShaoT.LiY.LiJ. and ZhangY., Long non-coding RNA XIST functions as an oncogene in human colorectal cancer by targeting miR-132-3p, J BUON22 (2017), 696–703.
40.
ChenD.L.JuH.Q.LuY.X.ChenL.Z.ZengZ.L.ZhangD.S.LuoH.Y.WangF.QiuM.Z.WangD.S.XuD.Z.ZhouZ.W.PelicanoH.HuangP.XieD.WangF.H.LiY.H. and XuR.H., Long non-coding RNA XIST regulates gastric cancer progression by acting as a molecular sponge of miR-101 to modulate EZH2 expression, J Exp Clin Cancer Res35 (2016), 142.
41.
HanQ.LiL.LiangH.LiY. and XieJ.WangZ., Downregulation of lncRNA X Inactive Specific Transcript (XIST) suppresses cell proliferation and enhances radiosensitivity by upregulating mir-29c in nasopharyngeal carcinoma cells, Med Sci Monit23 (2017), 4798–4807.
42.
WangH.ShenQ.ZhangX.YangC.CuiS.SunY.WangL.FanX. and XuS., The Long Non-Coding RNA XIST Controls Non-Small Cell Lung Cancer Proliferation and Invasion by Modulating miR-186-5p, Cell Physiol Biochem41 (2017), 2221–2229.
43.
WeiW.LiuY.LuY.YangB. and TangL., LncRNA XIST Promotes Pancreatic Cancer Proliferation Through miR-133a/EGFR, J Cell Biochem118 (2017), 3349–3358.
44.
LuJ., The Warburg metabolism fuels tumor metastasis, Cancer Metastasis Rev38 (2019), 157–164.
45.
SimoesR.V.SerganovaI.S.KruchevskyN.LeftinA.ShestovA.A.ThalerH.T.SukenickG.LocasaleJ.W.BlasbergR.G.KoutcherJ.A. and AckerstaffE., Metabolic plasticity of metastatic breast cancer cells: adaptation to changes in the microenvironment, Neoplasia17 (2015), 671–684.
46.
LiuY.HuoY.WangD.TaiY.LiJ.PangD.ZhangY.ZhaoW.DuN. and HuangY., MiR-216a-5p/Hexokinase 2 axis regulates uveal melanoma growth through modulation of Warburg effect, Biochem Biophys Res Commun501 (2018), 885–892.
47.
LiuX.CuiL. and HuaD., Long noncoding RNA XIST regulates miR-137-EZH2 axis to promote tumor metastasis in colorectal cancer, Oncol Res27 (2018), 99–106.
48.
JiZ.G.JiangH.T. and ZhangP.S., FOXK1 promotes cell growth through activating wnt/beta-catenin pathway and emerges as a novel target of miR-137 in glioma, Am J Transl Res10 (2018), 1784–1792.
49.
DingF.ZhangS.GaoS.ShangJ.LiY.CuiN. and ZhaoQ., MiR-137 functions as a tumor suppressor in pancreatic cancer by targeting MRGBP, J Cell Biochem119 (2018), 4799–4807.
50.
DengJ.LeiW.XiangX.ZhangL.LeiJ.GongY.SongM.WangY.FangZ.YuF.FengM.SunZ.ChenJ.ZhanZ. and XiongJ., Cullin 4A (CUL4A), a direct target of miR-9 and miR-137, promotes gastric cancer proliferation and invasion by regulating the Hippo signaling pathway, Oncotarget7 (2016), 10037–10050.
51.
SunJ.CaiX.YungM.M.ZhouW.LiJ.ZhangY.LiZ.LiuS.S.CheungA.N.Y.NganH.Y.S.LiY.DaiZ.KaiY.TzatsosA.PengW.ChanD.W. and ZhuW., miR-137 mediates the functional link between c-Myc and EZH2 that regulates cisplatin resistance in ovarian cancer, Oncogene38 (2019), 564–580.
52.
TaniguchiK.SugitoN.KumazakiM.ShinoharaH.YamadaN.NakagawaY.ItoY.OtsukiY.UnoB.UchiyamaK. and AkaoY., MicroRNA-124 inhibits cancer cell growth through PTB1/PKM1/PKM2 feedback cascade in colorectal cancer, Cancer Lett363 (2015), 17–27.
53.
LiangJ.CaoR.WangX.ZhangY.WangP.GaoH.LiC.YangF.ZengR.WeiP.LiD.LiW. and YangW., Mitochondrial PKM2 regulates oxidative stress-induced apoptosis by stabilizing Bcl2, Cell Res27 (2017), 329–351.
54.
MaR.LiuQ.ZhengS.LiuT.TanD. and LuX., PKM2-regulated STAT3 promotes esophageal squamous cell carcinoma progression via TGF-beta1-induced EMT, J Cell Biochem, 2019.
55.
LinY.MengF.LuZ.ChenK.TaoY.OuyangY. and CaoX., Knockdown of PKM2 suppresses tumor progression in human cervical cancer by modulating epithelial-mesenchymal transition via Wnt/beta-catenin signaling, Cancer Manag Res10 (2018), 4191–4202.
56.
TangJ.C.RenY.G.ZhaoJ.LongF.ChenJ.Y. and JiangZ., Shikonin enhances sensitization of gefitinib against wild-type EGFR non-small cell lung cancer via inhibition PKM2/stat3/cyclinD1 signal pathway, Life Sci204 (2018), 71–77.
57.
LuW.Q.HuY.Y.LinX.P. and FanW., Knockdown of PKM2 and GLS1 expression can significantly reverse oxaliplatin-resistance in colorectal cancer cells, Oncotarget8 (2017), 44171–44185.