Testing carcinogenicity caused by chemicals requires a noninvasive tool that can be used before autopsy because autopsy takes a long time. We investigated whether non-small cell lung cancer related gene mutations could be detected in cell-free DNA in plasma by Insight Onco next-generation sequencing, which is a fast and sensitive method. Adenoma formation was confirmed in urethane-injected 17-week-old mice. Seven single nucleotide polymorphisms, such as Cdkn2a and Vegfa, were selected. Mutant-enriched Insight Onco NGS and normal NGS were performed on genomic DNA. The results demonstrate that Insight Onco NGS detected Cdkn2a and Vegfa SNPs at 0.05%. The sensitivity of Insight Onco NGS was twice higher than that of normal NGS. In this analysis, the Cdkn2a gene mutation was detected not only in two genomic DNA samples of lung tissue from the 11th week of urethane injection but also in two cell-free DNA samples. In addition, the Vegfa gene mutation was detected not only in three genomic DNA samples of lung tissue of injection but also in one cell-free DNA sample, showing 33% concordance. Our results confirm that Insight Onco NGS is a rapid and sensitive detection method that enables lung cancer-associated gene mutations to be detected in cell-free DNA before the end of the carcinogenicity test.
Lung cancer is the leading cause of cancer mortality worldwide [18]. At least 150 chemical and biological agents are known or probable causes of cancer [12]. Many of these are found in the workplace, although occupational cancers are almost entirely preventable through eliminating exposure, substituting safer materials, enclosing processes, and using ventilation. To assess and evaluate the potential carcinogenicity of a chemical, animal carcinogenicity studies have been conducted worldwide. The gold standard test under the Registration, Evaluation, Authorization and Restriction of Chemicals (REACH) for carcinogenicity is OECD 451 [22]. Animal carcinogenicity studies are large-scale studies, with a relatively long term. The use of 400 rats and mice is recommended for such studies that are normally terminated 24 months later [14]. However, neoplastic histopathological findings cannot be predicted before the gross necropsy or toxicological pathology studies at study termination. A wide variety of murine model systems have been developed with the aim to evaluate novel therapeutics and examine the mechanisms underlying transformation, invasion, and metastasis in human tumors to develop better prevention, screening, diagnostic, and treatment strategies [13]. Carcinogen inducible models are typically generated in strains of inbred mice, such as A/J or SWR, which are very susceptible to spontaneous tumorigenesis [6]. Of these models, the urethane-induced lung tumorigenesis model has several advantages. An intraperitoneal administration of urethane has been shown to be reliably reproducible [20]. Subsequent tumorigenesis develops in a time-dependent manner. Tumorigenesis progresses from hyperplasia to adenoma and eventually adenocarcinoma in response to sequential genetic changes. This is the characteristic of human non-small cell lung cancer (NSCLC) [23]. When cancer forms, genes have many mutations, causing genetic heterogeneity in cancer. A recent genomic profiling study suggests that urethane-induced tumors have far more single-nucleotide variants (SNVs) than tumors induced by genetic activation of Kras [34]. Direct sequencing for human cancer gene mutation analysis has been used in clinical practice as a gold standard method [19]. Recently, it was reported that Insight Onco (mutant-enriched) next-generation sequencing (NGS) technology could be used to perform cancer gene mutation analyses quickly and effectively [35]. In addition, it has been reported that the amount of cell-free DNA (cfDNA) in the plasma increases when tumors form. Therefore, monitoring cfDNA in such plasma, as a liquid biopsy has been suggested for lung cancer patient management [5]. In this study, we applied Insight Onco NGS technology to detect mutations in lung cancer-related genes of tumorized lung tissue in blood plasma of urethane-administered mice. The applicability of this technique for the early detection of lung cancer-related genes in carcinogenicity testing was also determined in this study.
Materials and methods
Chemicals and reagents
Urethane (CAS No. 51-79-6) was purchased from Sigma-Aldrich (94300, St Louis, MO, USA, Lot No. BCBP1319V). The hazard classification of urethane, according to the United Nations Globally Harmonized System of Classification and Labeling of Chemicals (UN GHS), is Category 1B for both germ cell mutagenicity and carcinogenicity.
Animals
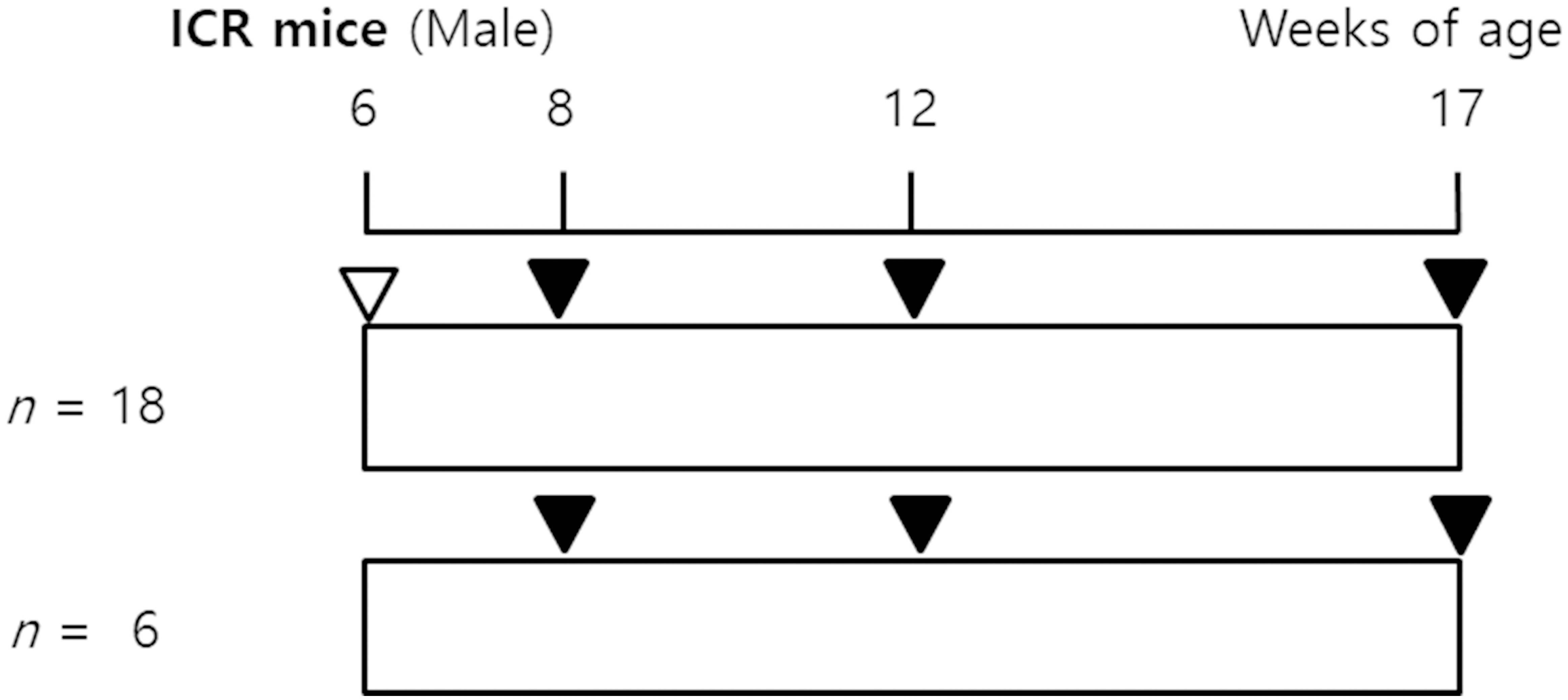
The experimental protocol is shown in Fig. 1. Male ICR (the Institute for Cancer Research in Philadelphia) mice, at 5-weeks-old, were purchased from Japan SLC, Inc. (Hamamatsu, Shizuoka, Japan). After acclimation for one week, six-week-old mice were used in the study. Food (a commercial diet PicoLab 5053 [PMI Nutrition International, MO, USA]) and water were given to mice ad libitum. The mice were housed in polysulfone cages (six individuals per cage) under controlled environmental conditions (temperature, 22 3C; humidity, 50 20%; lights on from 08:00 to 20:00 hr). The body weights (b.w.) of mice were monitored and recorded weekly. Eighteen male ICR mice were exposed to 1 mg/g b.w. urethane (CAS No. 51-79-6, Aldrich 94300) by i.p. injection. Six control mice were injected with vehicle control (0.9% NaCl saline) in the same manner. The mice exposed to urethane were studied at 2 weeks, 6 weeks, and 11 weeks after urethane exposure ( 6 at each time point). At 2 weeks, 6 weeks, and 11 weeks after an acute single exposure to urethane, the blood, and lung tissues were collected for a whole exome sequencing (WES) analysis, and histological examinations, respectively. The animal study protocol was reviewed and approved by the Institutional Animal Care and Use Committee/Animal Ethic Committee (IACUC/AEC) of the Occupational Safety and Health Research Institute (Approval No: IACUC1602).
Overview of the study design. Six-week-old male ICR mice were injected with 1000 mg/kg urethane and were killed 2 weeks, 6 weeks, and 11 weeks later. , intraperitoneal urethane; , sampling.
PNA probes used in this study
PNA probe
Probe sequence (N terminus to C terminus)
Size (bp)
Tm (C)
Probe location (base positions in gene)
Cdkn2a
cgtgtgcatgacgtgc
16
59
127–142
Vegfa
acaccccggggctc
14
56
570–583
Extraction of genomic DNA (gDNA) and cfDNA
The right lung lobes were collected from all the mice and were stored at 80C until the total gDNA extraction using the High Pure PCR Template Preparation Kit (Roche, Basel, Switzerland) according to the manufacturer’s instructions. Under CO anesthesia, the blood was collected from the abdominal aorta of mice into tubes, containing EDTA followed by centrifugation (3,000 rpm, 10 min, 4C). cfDNA was extracted from plasma using QIAamp circulating nucleic acid kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instruction. The quantity and quality of the gDNA were checked with a NanoDrop 2000 Spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA) and the PicoGreen assay. The extracted DNA samples were stored at 20C until use.
WES and analysis of gDNA
For the WES analysis, gDNA from 17-week-old mice was used. The sequencing libraries were prepared using the SureSelect Mouse All Exon Kit (Agilent Technologies, Inc., Santa Clara, CA, USA). The pooled libraries were enriched for the exome through hybridization and were pooled in 1:1 ratio for paired-end sequencing on a HiSeq2000 platform (Illumina, San Diego, CA, USA). The quality of the library obtained was assessed by Agilent 2100 Bioanalyzer on-chip electrophoresis (Agilent Technologies, Inc., Santa Clara, CA, USA). The sequencing reads were aligned to a mouse reference (mm10). The annotation for all the variants was made using dbSNP137 (NCBI, release 20120626). The Comparative Toxicogenomics Database (CTD; http://ctdbase.org) was used to identify known interactions of the genes with urethane.
Peptide nucleic acid (PNA) – mediated clamping PCR of Cdkn2a and Vegfa
The PNA-mediated clamping method was used to detect the Cdkn2a and Vegfa gene mutations by real-time PCR. The PNA probe sequences (Table 1) were obtained using PNA Probe Designer software (Applied Biosystems, Waltham, MA, USA) (http://www.appliedbiosystems.com/cgi-bin/calculator/ab_configured/oligodesigner/designer.cgi) and were synthesized by Panagene (Daejeon, South Korea). For the NGS library preparation, the following primer pairs were used for each site: Cdkn2a-forward, 5’-GCTGTGACTGATGT ATGACGtggagcagcatggagtccgc-3’; Cdkn2a-reverse, 5’-GCGATGCGAGATAGTAGATGtccttgcctacctgaatc gg-3’; Vegfa-forward, 5’-GCTGTGACTGATGTATGA CGgatctgtagagggagggagc-3’; Vegfa-reverse, 5’-GCGA TGCGAGATAGTAGATGctggtcctcctatctccacc-3’. All the reactions were performed in a total of 20 , including 15 ng of template DNA, a primer and PNA probe set, and a fluorescence dye PCR master mix. Real-time PCR was carried out on a CFX 96 instrument (Bio-Rad, Philadelphia, PA, USA). The thermal cycling conditions were as follows: 10 minute hold at 94C followed by 40 cycles of 94C for 30 seconds; 70C for 20 seconds; 58C for 40 seconds; and 72C for 30 seconds. To determine the efficiency of the PCR, the automatic threshold cycle (Ct) value was measured. The delta Ct (Ct-1) was calculated according to the criteria reported by Yoon et al. [35]. Briefly, the Ct-1 was calculated as the tested sample Ct value minus the wild-type control sample Ct value. When the Ct-1 value was higher than 2.0, the sample was considered to have a mutant status. When the Ct-1 value was between 0 and 2.0, the Ct-2 value was calculated as the sample Ct value minus the non-PNA Ct value. The sample was judged to have a mutant status if the Ct-2 value was less than 3.0.
List of lung cancer related target genes selected in this study
Gene
Protein
Single nucleotide polymorphism
Allele change
Amino acid
Residue change
database (dbSNP)
position
Apba1
Amyloid beta (A4) precursor protein
rs31235157
GTC GCC
142
V[Val] A[Ala]
binding, family A, member 1
Apc
Adenomatous polyposis coli gene
rs47505115
GAT GGT
2086
D[Asp] G[Gly]
Cdkn2a
Cyclin-dependent kinase inhibitor 2A
rs224423625
GTA ATA
51
V[Val] I[Ile]
Cdkn2a
Cyclin-dependent kinase inhibitor 2A
rs244465024
CAT CCT
18
H[His] P[Pro]
Erbb2
Erb-b2 receptor tyrosine kinase 2
rs29390172
GCC ACC
130
A[Ala] T[Thr]
Sema3b
Semaphorin 3B
rs29789812
ACC GCC
122
T[Thr] A[Ala]
Sftpc
Surfactant associated protein C
rs8266234
ACT GCT
163
T[Thr] A[Ala]
Vegfa
Vascular endothelial growth factor A
rs16821352
CCG CTG
5
P[Pro] L[Leu]
Enrichment PCR for mutant-enriched NGS and sequencing library preparation
To maximize the resolution for the detection of low levels of the Cdkn2a or Vegfa mutants, an Insight Onco Panel for Cdkn2a or Vegfa (SeaSun Biomaterials, Daejeon, Korea) was designed for mutant enrichment PCR according to the manufacturer’s instructions. The mutant specific enrichment PCR was run using 10 ng of gDNA. We analyzed the samples according to the manufacturer’s instructions. Nonspecific PCR products were eliminated by Agencourt AMPure XP beads (Beckman Coulter, Brea, CA, USA), using a 1:1 DNA-to-bead ratio. For sequencing the Library Preparation PCR, 2 of purified PCR products (from enrichment PCR amplification) was used as template with the Insight Onco 2x Seq Lib Pep Premix (SeaSun Biomaterials, Daejeon, Korea) and each barcoded primer pair. A sequencing adaptor with a multiplex identifier (MID) was conjugated using the manufacturer’s instructions. The unwanted short fragments were eliminated by Agencourt AMPure XP beads (Beckman Coulter, Brea, CA, USA), using a 1:1 DNA-to-bead ratio.
Quantification and normalization of the sequencing amplicons
The isolated amplicons were read on an external Infinite F200Pro fluorometer (Tecan, Männedorf, Zürich, Switzerland) using Magellan v7.0 Software (Tecan, Männedorf, Zürich, Switzerland). Based on the standard curves, the signals were calculated to ng/l. The coefficient of determination (validation criteria R 0.99) was obtained using eight DNA standards at concentrations from 0 to 100 ng/l. To perform the emulsion PCR amplification, the concentrations of amplicons were calculated to molecules/l using the associated amplicon length. The manufactured DNA pools were stored at 20C until further use.
Pyrosequencing
Pyrosequencing was conducted according to the manufacturer’s protocol for the amplicons using GS Junior System (Roche Diagnostics, Basel, Switzerland). The GS Junior Titanium emPCR Kit Lib-L, emPCR Reagents Lib-L kit, Oil and Breaking Kit, and the Bead Recovery Reagents Kit (Roche Diagnostics, Basel, Switzerland) were used for the emulsion PCR, breaking, and bead enrichment according to the supplier’s instructions. For the emulsion PCR, a copy-per-bead ratio of 0.5 was adopted. The enrichment of DNA carrying beads was obtained using magnetic beads and a magnetic particle collector (Invitrogen, Waltham, MA, USA). To examine the number of enriched beads, a reading was conducted using the GS Junior Bead Counter (Roche Diagnostics, Basel, Switzerland). After approximately 100,000 500,000 beads were transferred onto a PicoTiterPlate (PTP) (Roche Diagnostics, Basel, Switzerland), sequencing was performed with a GS Titanium Sequencing Kit (Roche Diagnostics, Basel, Switzerland) and a GS Junior device according to standard Roche/454 protocols.
Data analysis
Processed and quality-filtered reads were assessed using a GS Amplicon Variant Analyzer (Roche Diagnostics, Basel, Switzerland). The sequencing reads were visualized using a GS Amplicon Variant Analyzer. The Cdkn2a and Vegfa amplicons (excluding the adaptors and MIDs) were used as references to align the amplicon reads. We defined known mutations of the samples selected as substitutions relative to the reference sequence.
Results
Survival and body weight
As shown in Fig. 1, all the animals survived to 17 weeks. There were no overt signs of toxicity due to urethane (data not shown). Significant differences in b.w. were noted between the 1,000 mg/kg urethane treatment group and the control group of animals starting at 8 weeks (data not shown).
Growth of lung tumors
The incidence of adenoma at 6 and 11 weeks was 33% and 66%, respectively (data not shown). Papillary-type adenomas were observed. The tumor cells showed a columnar epithelium with a basophilic cytoplasm. Pleomorphic nuclei, mitotic cells, or invasion of surrounding tissue was not observed.
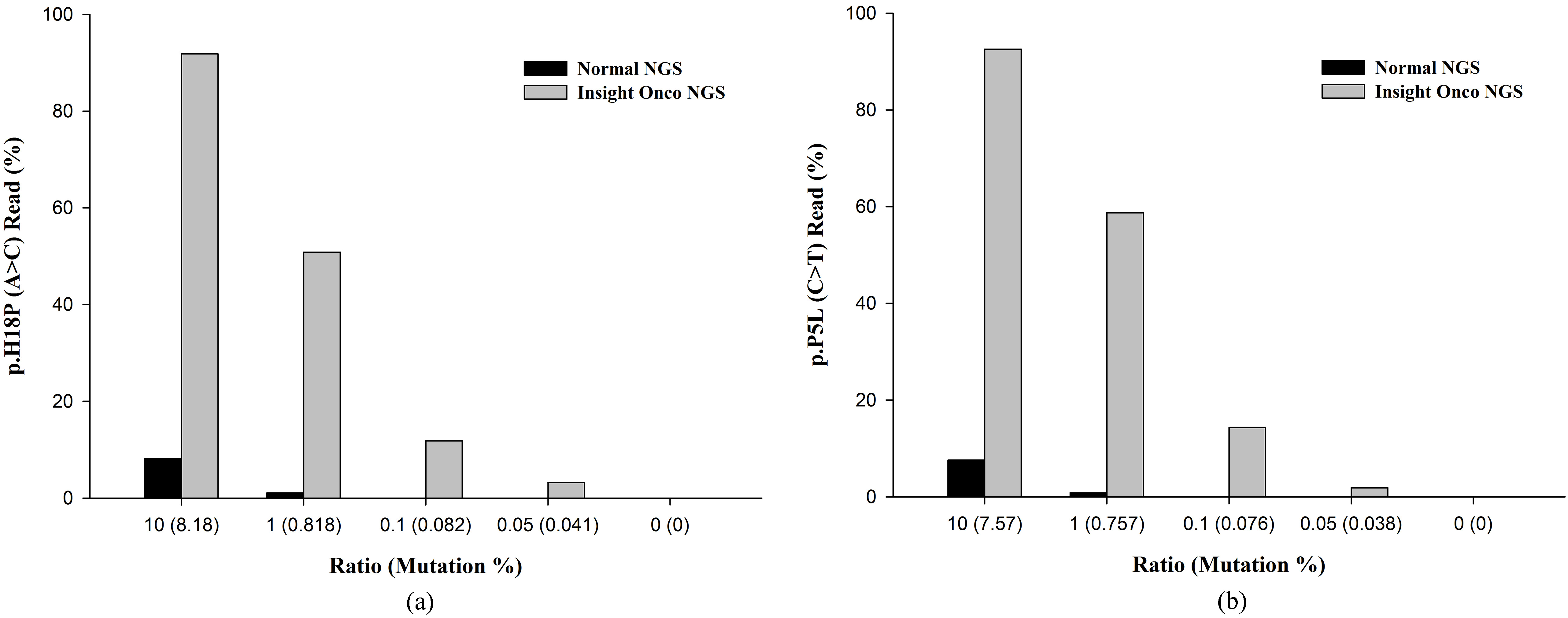
Comparison of conventional next-generation sequencing (NGS) and mutant-enriched NGS for detecting the Cdkn2a (a) and Vegfa (b) mutations in gDNA. More than 1% of mutant DNA is needed for conventional NGS to detect mutations, and in contrast, mutant-enriched NGS can detect mutant DNA with as low as 0.05%.
Somatic DNA alterations
To understand the operative processes involved in the progression to adenoma, the WES was performed using the ICR strain with urethane-induced and histologically confirmed lung adenomas. Of the 38,572 coding variants observed, 13,047 variants were predicted to cause amino acid changes (nonsynonymous), while 440 were called as coding indels. In this analysis, a total of 1,844 single nucleotide polymorphisms (SNPs) and indels passed the pipeline filters. Based on the annotations and the literature, seven target genes were identified, including Apc, Cdkn2a, Erbb2, and Sema3b, which were previously determined as mutated in lung cancer, and others, such as Apba1, Sftpc, and Vegfa, had roles in tumorigenesis but were not previously implicated in lung cancer [7]. Determining which of these genes might be true driver mutations in lung cancer requires functional characterization. Mutations in tumor suppressor genes, including Apc, Cdkn2a, and Sema3b, were observed in this study. Mutations in oncogenes, including Erbb2, were also observed (Table 2). In this study, we could not detect KRAS and TP53 mutations. We presumed that this difference was probably due to the use of the outbred ICR mice, unlike many other studies using inbred mice (e.g., A/J).
Analysis of plasma DNA using PNA-clamping real-time PCR and pyrosequencing by NGS
As a source of genetic material, the use of plasma cfDNA (a non-invasive specimen) coupled with NGS offers a great option for cancer screening, early detection, and disease monitoring because plasma may reflect the disease status instead of performing a tumor biopsy. First, we compared the sensitivity of conventional NGS and mutant-enriched NGS by diluting the control DNA. The Cdkn2a and Vegfa mutations were detected by conventional NGS when the mutant allele comprised more than 1% of the sample. However, with the mutant-enriched NGS, the mutation was detected when the mutant allele comprised only 0.05% of the sample (Fig. 2).
Comparison of normal NGS and mutant-enriched NGS (Insight Onco NGS) in detecting the Cdkn2a mutation in lung tissue samples and plasma samples
Cdkn2a p.H18P (A C)
Period (week)
Animal ID
Lung tissue samples (gDNA)
Plasma samples (cfDNA)
Normal NGS
Insight Onco NGS
Normal NGS
Insight Onco NGS
(%)
Depth
(%)
Depth
(%)
Depth
(%)
Depth
2
1003
0
3204
0
3465
0
1295
0
1105
(Control)
1004
0
2980
0
4021
0
1648
0
1956
2
1101
0
2074
0
2989
0
1963
0.14
1200
(Urethane-
1103
0
2674
0
2678
0
1756
0
986
treated)
1104
0
2871
0
2956
0
1944
0
979
6
1002
0
2094
0
3495
0
1238
0
1016
(Control)
1005
0
3109
0
4025
0
1956
0.16
1198
6
1201
0
3100
0
4951
0
986
0
2004
(Urethane-
1202
0
2730
0
2945
0.1
1297
0
1920
treated)
1204
0
2105
0
3756
0
1193
0
1946
11
1001
0
2978
0
3469
0.14
1935
0
987
(Control)
1006
0
3661
0
3946
0
2103
0
1624
11
1302
49.54
2287
99.22(2)
4852
0.89
1869
14.92 (16.76)
2630
(Urethane-
1305
0
3529
0
5461
0
1679
0
1932
treated)
1306
100
2947
100 (1)
4612
0
1975
3.2 (3.2)
2193
The numbers in parentheses denote the fold change between the Normal NGS and the Insight Onco NGS samples.
Comparison of the normal NGS and the mutant-enriched NGS (Insight Onco NGS) when detecting the Vegfa mutation in lung tissue samples and plasma samples
Vegfa p.P5L(CT)
Period (week)
Animal ID
Lung tissue samples (gDNA)
Plasma samples (cfDNA)
Normal NGS
Insight Onco NGS
Normal NGS
Insight Onco NGS
(%)
Depth
(%)
Depth
(%)
Depth
(%)
Depth
2
1003
0
2495
0
2953
0
1209
0
2953
(Control)
1004
0
2149
0
2898
0.12
983
0
2898
2
1101
0
2658
0
2963
0
976
2963
(Urethane-
1103
0
1953
0
2754
0
891
0
2754
treated)
1104
0
1993
0
3479
0.17
1185
0
3479
6
1002
0
2015
0
2965
0
892
0
2965
(Control)
1005
0
2953
0
2378
0
836
2378
6
1201
0
3268
0
2496
0
1203
0
2496
(Urethane-
1202
0
2956
0
2298
0.13
1098
0
2298
treated)
1204
0
2985
0
3298
0
1327
0
3298
11
1001
0
2127
0
4512
0
980
0
4512
(Control)
1006
0
2643
0
2986
0
1034
0.19
2986
11
1302
98.39
2186
99.92 (1.02)
4145
0.11
968
3.29 (29.9)
4145
(Urethane-
1305
52.96
2270
91.26 (1.72)
2562
0.09
1408
0.42
2562
treated)
1306
94.18
2189
95.73 (10.2)
3844
0
1763
0.92
3844
The numbers in parentheses denote the fold change between the Normal NGS and the Insight Onco NGS samples.
Comparative study of the NGS application for tissue and plasma detection
Neoplastic tissue remains the standard specimen for molecular analysis [11]. However, the potential to obtain a noninvasive sample compared to a tissue biopsy is very attractive. Blood collection is less invasive than tissue sampling. It can be used when tissue specimens are limited or unavailable. Moreover, it allows for sampling at several time points to monitor the genetic evolution of the tumor. Both Cdkn2a and Vegfa are related to the AKT/PI3K pathways in lung cancer [28]. A recent study demonstrates that the mean tumor diameters in animals correlate with tumor DNA detection in the plasma [4]. We performed a comparative study of the NGS application for tissue and plasma detection of these genes. However, we did not find mutated DNA sequences in plasma or lung tissue samples from mice at 6 weeks after the urethane treatment. cfDNA was extracted from the plasma of 3 animals per group and was analyzed. Mutated Cdkn2a sequences were detected in two of the three plasma samples obtained from the mice (2/3; 66%) at 11 weeks after the urethane treatment, using Insight Onco (mutant-enriched) NGS (Table 3). However, no mutated DNA in the plasma was detected using conventional NGS. We detected mutated Cdkn2a sequences in the nodules from 2 of 3 (66%) animals that had lung adenoma. Mutated Vegfa sequences were detected in one of three (1/3; 33%) plasma samples from the mice at 11 weeks after the urethane treatment using mutant-enriched NGS (Table 4). However, no mutated DNA in the plasma was detected using conventional NGS. We detected mutated Vegfa sequences in nodules from 3 of 3 (100%) animals that had lung adenoma. The difference in the incidence of the Cdkn2a and Vegfa mutation detection might be related to the copy number variations of these genes.
Discussion
Worldwide, occupational exposures account for an estimated 8% of lung cancer, the most frequent form of occupational cancer [24]. Phenomena of intra-cancer heterogeneity (genetic, cellular, and epigenetic), cross-cancer similarity, frequent point mutations, and frequent gene copy number changes have been observed in human cancers, making lung cancer an intractable disease [3]. Blood sampling is minimally invasive. In addition, blood is a source of fresh DNA with good quality for molecular analyses [35]. Considering that cfDNA contains the same genetic defects as tumor tissue, cfDNA sampling is often compared to standard tissue biopsy and is called a liquid biopsy [32]. In this study, Insight Onco NGS revealed that single nucleotide variations (SNVs) of cfDNA in the plasma and SNVs in adenoma coincided with each other in the urethane model, a well-known lung cancer model. In mice, urethane induces lung adenocarcinomas (ACs). The time frames for the development of hyperplasias, adenomas, and ACs is well characterized in mice. Urethane-induced lung adenocarcinoma resembles the adenocarcinoma subtype of human non-small cell lung carcinoma [27], and also, urethane induces lung ACs. Histopathologically, murine ACs are indistinguishable from human ACs [33]. ACs are associated with mutations in the Kras oncogene and tumor suppressor genes, such as Tp53 in humans and mice [9]. Urethane is carcinogenic, mutagenic, and clastogenic in vivo. However, it is generally considered non-genotoxic in vitro due to its unique metabolism requirements [29]. Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) is a ubiquitous key modulator of cellular defense against oxidative stress and inflammation. It plays a cell survival role in urethane-induced pulmonary neoplasia [1]. In Friend leukemia virus B (FVB) mice, urethane-induced NF-B activation is identified in airway epithelium, type II alveolar epithelial cells, and macrophages [30]. In this study, an examination of paired exomes from the normal lung specimens and lung specimens obtained from the mice with adenoma led to the discovery of 38,727 somatic mutations. Of these 38,727 somatic mutations, 1,844 were detected in coding regions, and seven target genes related to lung cancer were identified based on the annotation and literature search. Some homozygous alleles have a high frequency of gene variations [2]. Among the seven driver genes (Apba1, Apc, Cdkn2a, Erbb2, Sema3b, Sftpc, and Vegfa) found in the WES of lung adenoma, we selected two genes (Cdkn2a and Vegfa) and confirmed that two mutations found in lung adenoma were also found in plasma cfDNA (Tables 3 and 4). In NSCLC, the p16/RB pathway is mostly switched off through alterations of cyclin D1, CDK4, and cyclin dependent kinase inhibitor p16 (CDKN2A) [31]. p16 is inactivated in approximately 80% of NSCLC. It is also altered in 72% of lung squamous cell carcinomas (SCCs) examined by The Cancer Genome Atlas (TCGA) mostly through homozygous deletion, methylation, or inactivating mutagens [10]. Human vascular endothelial growth factor A (VEGF-A) is a key regulator of angiogenesis under Wnt control [25]. Recent studies have revealed that VEGF SNPs are associated with VEGF expression and that high VEGF expression is associated with high vascular density [16]. PNA is a DNA mimic that binds strongly to its complementary DNA sequence [17]. PNA clamping enables the detection of Cdkn2a and Vegfa gene mutations in samples containing as few as 1% mutant alleles. As expected, the detection rate by PNA clamping was significantly higher than that by deep sequencing. In the plasma samples from a 6-week-old and an 11-week-old mouse of the control group, the Insight Onco NGS values for the Cdkn2a and Vegfa mutations were 0.16% and 0.19%, respectively. We estimated that these values were due to the background noise generated by the PCR. In this study, the incidence rates for the detection of the Cdkn2a and Vegfa mutations using cfDNA and the tissue samples were compared. The cfDNA test concordance rates were 66% and 33% for assessing the Cdkn2a and Vegfa mutations, respectively. There were 34% and 67% discordance between the tissue and plasma samples for the detection of the Cdkn2a and Vegfa mutations in this study. A possible reason for this discrepancy might be due to the time-points for collecting the blood samples, which might be earlier than those for collecting the tissue samples. In the present study, we failed to detect mutated Cdkn2a or Vegfa sequences in tumor-bearing mice examined at 6 weeks after urethane exposure. Both the frequency of such detections and the levels of mutated DNA increase with time and tumor diameter [8]. Once data from the WES are accumulated according to group of chemicals, it will be possible to trace back the target organ early through the SNVs analysis of cfDNA when performing carcinogenicity tests for certain chemicals. There are over 100,000 synthetic chemicals in commerce today. Most of these chemicals are not known to be harmful. Carcinogenicity tests are being conducted mainly for chemicals suspected of causing cancer or that have a large amount of circulation and handling. Although mouse models clearly have different aspects from that of humans [21], such as the lack of metastasis, mutations in both genes (Cdkn2a and Vegfa) are common in many cancers, including lung cancer. Therefore, mutations in Cdkn2a and Vegfa can be used as indicators of tumorigenesis in mice. There are enough data showing that urethane causes cancer in the lung [15]. Therefore, mice treated with urethane were used in this study. Ultimately, we conducted this study to investigate the possibility of using plasma cfDNA as an early diagnostic marker for predicting the carcinogenicity of chemicals unknown to the target organ. Since cfDNA is an important biomarker derived from cells that are detached from cancer tissues, its profile might be different according to the origin of the cancer tissue. Sheikine et al. [26] recently reported the approximate lower limit of detection of mutant allelic frequencies (AF) of allele-specific ddPCR and NGS test methods at 0.1%. Further studies are needed to compare the performances among ddPCR, NGS, and Insight Onco (mutant-enriched) NGS techniques. In addition, further studies on copy number alterations in cfDNA are needed to unravel its potential as a biomarker in lung cancer.
This study has a number of strengths. It is the first trial that uses cfDNA for mutation detection in carcinogenicity studies. However, this study also has several limitations, including its small sample size and low number of recurrences. Despite these limitations, our study provides a proof-of-concept and attests the feasibility of performing cfDNA analysis for mice with early-stage NSCLC.
In conclusion, Insight Onco (mutant-enriched) NGS technology can be used for tumor detection in animal carcinogenesis as a sensitive assay. Our approach will allow researchers to reduce the cost of identifying genetic mutations in cancer development due to exposure to chemicals.
Conflict of interest
The authors have no conflicts of interest to declare.
Footnotes
Acknowledgments
This research was supported by a research fund from the Occupational Safety and Health Research Institute, Korea Occupational Safety and Health Agency (OSHRI, KOSHA). We thank Dr. Jin-Wook Chung and Dr. Hanwoo Lee (SeaSun Biomaterials, Daejeon, Korea) for their assistance with the mutant-enriched NGS.
References
1.
BauerA.K.ChoH.Y.Miller-DegraffL.WalkerC.HelmsK.FostelJ.YamamotoM. and KleebergerS.R., Targeted deletion of Nrf2 reduces urethane-induced lung tumor development in mice, PLoS One6 (2011), e26590.
2.
BunzF., Principles of cancer genetics, Springer, Dordrecht, 2016.
3.
CirielloG.MillerM.L.AksoyB.A.SenbabaogluY.SchultzN. and SanderC., Emerging landscape of oncogenic signatures across human cancers, Nat Genet45 (2013), 1127–1133.
4.
CzeigerD.ShakedG.EiniH.VeredI.BelochitskiO.AvrielA.AriadS. and DouvdevaniA., Measurement of circulating cell-free DNA levels by a new simple fluorescent test in patients with primary colorectal cancer, Am J Clin Pathol135 (2011), 264–270.
5.
DakuboG.D., Cancer biomarkers in body fluids: principles, Springer, Cham, Switzerland, 2016.
6.
de SerannoS. and MeuwissenR., Progress and applications of mouse models for human lung cancer, Eur Respir J35 (2010), 426–443.
7.
MairingerF., Pulmonary Neuroendocrine Tumours – Different Biological Entities? Duisburg, Essen, Universität Duisburg-Essen, Diss., 2014.
8.
Garcia-OlmoD.C.PicazoM.G.TobosoI.AsensioA.I. and Garcia-OlmoD., Quantitation of cell-free DNA and RNA in plasma during tumor progression in rats, Mol Cancer12 (2013), 8.
9.
GibbonsD.L.ByersL.A. and KurieJ.M., Smoking, p53 mutation, and lung cancer, Mol Cancer Res12 (2014), 3–13.
10.
GreulichH., The Genomics of Lung Adenocarcinoma Opportunities for Targeted Therapies, Genes & Cancer1 (2010), 1200–1210.
11.
HassaneinM.CallisonJ.C.Callaway-LaneC.AldrichM.C.GroganE.L. and MassionP.P., The state of molecular biomarkers for the early detection of lung cancer, Cancer Prev Res (Phila)5 (2012), 992–1006.
12.
IARC Working Group on the Evaluation of Carcinogenic Risks to Humans., World Health Organization. and International Agency for Research on Cancer., Man-made vitreous fibres, International Agency for Research on Cancer; World Health Organization, Lyon, France, 2002.
13.
KellarA.EganC. and MorrisD., Preclinical Murine Models for Lung Cancer: Clinical Trial Applications, Biomed Res Int2015 (2015), 621324.
14.
KlaassenC.D.WatkinsJ.B. and CasarettL.J., Casarett & Doull’s essentials of toxicology, McGraw-Hill Medical Pub. Division, New York, 2010.
15.
LiX.WuJ.ZhengJ.LiY.YangT.HuG.DaiJ.YangQ.DaiL. and JiangY., Altered miRNA expression profiles and miR-1a associated with urethane-induced pulmonary carcinogenesis, Toxicol Sci135 (2013), 63–71.
16.
MaedaA.NakataM.YasudaK.YukawaT.SaishoS.OkitaR.HiramiY. and ShimizuK., Influence of vascular endothelial growth factor single nucleotide polymorphisms on non-small cell lung cancer tumor angiogenesis, Oncol Rep29 (2013), 39–44.
17.
MiyazawaH.TanakaT.NagaiY.MatsuokaM., HuqunSutaniA.UdagawaK.ZhangJ.HiramaT.MurayamaY.KoyamaN.IkebuchiK.NagataM.KanazawaM.NukiwaT.TakenoshitaS.KobayashiK. and HagiwaraK., Peptide nucleic acid-locked nucleic acid polymerase chain reaction clamp-based detection test for gefitinib-refractory T790M epidermal growth factor receptor mutation, Cancer Sci99 (2008), 595–600.
18.
MolinaJ.R.YangP.CassiviS.D.SchildS.E. and AdjeiA.A., Non-small cell lung cancer: epidemiology, risk factors, treatment, and survivorship, Mayo Clin Proc83 (2008), 584–594.
19.
MonzonF.A.OginoS.HammondM.E.HallingK.C.BloomK.J. and NikiforovaM.N., The role of KRAS mutation testing in the management of patients with metastatic colorectal cancer, Arch Pathol Lab Med133 (2009), 1600–1606.
20.
NemmarA.YuvarajuP.BeegamS. and AliB.H., Betaine (N, N, N-trimethylglycine) averts photochemically-induced thrombosis in pial microvessels in vivo and platelet aggregation in vitro, Exp Biol Med (Maywood)240 (2015), 955–960.
21.
NikitinA.Y.AlcarazA.AnverM.R.BronsonR.T.CardiffR.D.DixonD.FraireA.E.GabrielsonE.W.GunningW.T.HainesD.C.KaufmanM.H.LinnoilaR.I.MaronpotR.R.RabsonA.S.ReddickR.L.RehmS.RozengurtN.SchullerH.M.ShmidtE.N.TravisW.D.WardJ.M. and JacksT., Classification of proliferative pulmonary lesions of the mouse: recommendations of the mouse models of human cancers consortium, Cancer Res64 (2004), 2307–2316.
22.
OECD, Test No. 451: Carcinogenicity Studies, OECD Publishing, 2009.
23.
OhnoJ.HorioY.SekidoY.HasegawaY.TakahashiM.NishizawaJ.SaitoH.IshikawaF. and ShimokataK., Telomerase activation and p53 mutations in urethane-induced A/J mouse lung tumor development, Carcinogenesis22 (2001), 751–756.
24.
Pruss-UstunA.VickersC.HaefligerP. and BertolliniR., Knowns and unknowns on burden of disease due to chemicals: a systematic review, Environ Health10 (2011), 9.
25.
RappJ.KissE.MeggyesM.Szabo-MelegE.FellerD.SmukG.LaszloT.SarosiV.MolnarT.F.KvellK. and PongraczJ.E., Increased Wnt5a in squamous cell lung carcinoma inhibits endothelial cell motility, BMC Cancer16 (2016), 915.
26.
SheikineY.RangachariD.McDonaldD.C.HubermanM.S.FolchE.S.VanderLaanP.A. and CostaD.B., EGFR Testing in Advanced Non-Small-Cell Lung Cancer, A Mini-Review, Clin Lung Cancer17 (2016), 483–492.
27.
SongJ.S.JungS.H.YiS.Y.OhH.E.ChoM.Y. and ParkK.H., Telomerase Activity in Urethane-Induced Mouse Lung Tumorigenesis, Korean J Pathol45 (2011), 261–270.
28.
SparaneoA.FabrizioF.P. and MuscarellaL.A., Nrf2 and Notch Signaling in Lung Cancer: Near the Crossroad, Oxid Med Cell Longev2016 (2016), 7316492.
29.
StankowskiL.F., Jr.AardemaM.J.LawlorT.E.PantK.RoyS.XuY. and ElbekaiR., Integration of Pig-a, micronucleus, chromosome aberration and comet assay endpoints in a 28-day rodent toxicity study with urethane, Mutagenesis30 (2015), 335–342.
30.
StathopoulosG.T.SherrillT.P.ChengD.S.ScogginsR.M.HanW.PolosukhinV.V.ConnellyL.YullF.E.FingletonB. and BlackwellT.S., Epithelial NF-kappaB activation promotes urethane-induced lung carcinogenesis, Proc Natl Acad Sci U S A104 (2007), 18514–18519.
31.
TamK.W.ZhangW.SohJ.StastnyV.ChenM.SunH.ThuK.RiosJ.J.YangC.MarconettC.N.SelamatS.A.Laird-OffringaI.A.TaguchiA.HanashS.ShamesD.MaX.ZhangM.Q.LamW.L. and GazdarA., CDKN2A/p16 inactivation mechanisms and their relationship to smoke exposure and molecular features in non-small-cell lung cancer, J Thorac Oncol8 (2013), 1378–1388.
32.
VendrellJ.A.Mau-ThemF.T.BegantonB.GodreuilS.CoopmanP. and SolassolJ., Circulating Cell Free Tumor DNA Detection as a Routine Tool forLung Cancer Patient Management, Int J Mol Sci18 (2017).
33.
WakamatsuN.DevereuxT.R.HongH.H. and SillsR.C., Overview of the molecular carcinogenesis of mouse lung tumor models of human lung cancer, Toxicol Pathol35 (2007), 75–80.
34.
WestcottP.M.HalliwillK.D.ToM.D.RashidM.RustA.G.KeaneT.M.DelrosarioR.JenK.Y.GurleyK.E.KempC.J.FredlundE.QuigleyD.A.AdamsD.J. and BalmainA., The mutational landscapes of genetic and chemical models of Kras-driven lung cancer, Nature517 (2015), 489–492.
35.
YoonS.H.ChoiY.D.OhI.J.KimK.S.ChoiH.ChangJ.ShinH.J.ParkC.K. and KimY.C., Peptide Nucleic Acid Clamping Versus Direct Sequencing for the Detection of EGFR Gene Mutation in Patients with Non-small Cell Lung Cancer, Cancer Res Treat47 (2015), 661–669.