
Abstract
Mealybugs (Hemiptera: Coccomorpha: Pseudococcidae) are pests in coffee, Coffea arabica, in different coffee-producing countries and affect both the roots and the aerial part; however, their biology and population dynamics remain poorly studied. This study aimed to evaluate the life cycle of Pseudococcus elisae and analyze the field dynamics of mealybugs species present in coffee roots. The life cycle was carried out under controlled conditions (25 °C ± 2 °C, 70% - 80% RH, and photoperiod 12:12 h [L: D]), on 100 four-month-old coffee plants and 100 fruits of Cucurbita moschata; additionally, fecundity and fertility parameters were recorded. In C. arabica the duration from egg to adult was 98.1 ± 1.3 days for females and 68 ± 0.4 days for males; each female lays an average of 231 ± 3.6 eggs from which 197.3 ± 6.2 nymphs emerged; the sexual ratio was 11♀: 6♂. In C. moschata the duration from egg to adult was 92.1 ± 0.3 days for females and 67.2 ± 0.3 days for males, each female lays an average of 201 ± 3.6 eggs, of which 182.7 ± 4.8 nymphs emerged, the sexual ratio was 8♀: 8♂. In the field, it was found that 70.4% of the plants had mealybugs, and they were observed from the first month of age; of these, 82.7% had P. elisae and Dysmicoccus texensis species-complex, which encysted the roots of coffee plants starting at eleven months of age.
Introducción
El café, Coffea arabica L. (Rubiaceae), es uno de los cultivos con mayor importancia económica en el mundo. Sin embargo, en los últimos años en Colombia algunas especies de cochinillas harinosas (Hemiptera: Coccomorpha: Pseudococcidae) han venido causando preocupación a los caficultores en las diferentes regiones productoras del grano (Gil et al., 2015). Estos insectos son considerados de gran importancia ya que son polífagos y afectan diversos cultivos (Dhaliwal et al., 2010; Williams & Granara de Willink, 1992). Estudios recientes reportaron para Colombia, al menos 65 especies de cochinillas harinosas presentes en las raíces de café, donde las familias de mayor importancia fueron Putoidae, con la especie Puto barberi (Cockerell, 1895) y Pseudococcidae con Dysmicoccus Ferris, 1950 y Pseudococcus Westwood, 1840, destacándose la presencia de Pseudococcus elisae Borchsenius, 1947 y Pseudococcus jackbeardsleyi Gimpel & Miller, 1996, y de especies del complejo Dysmicoccus texensis (Tinsley, 1900), (Caballero et al., 2019). Estas especies de Pseudococcidae se han reportado en asociación con hongos basidiomicetos, que colonizan desde la base de la raíz hasta las raíces primarias y secundarias, formando una estructura corchosa con cientos de cochinillas harinosas en su interior. El micelio de este hongo cubre todo el rizoma, generando una capa de protección para los insectos y los árboles afectados se debilitan hasta su muerte (Gil et al., 2015; Santa-Cecilia et al., 2002; Souza et al., 2008).
Los reportes sobre cochinillas harinosas asociadas con hongos han sido en su gran mayoría en café y uva; en café, en Brasil, Costa Lima (1930) reportó a Pseudococcus cryptus Hempel, 1918, asociado a Bornetina sp. Maublanc y Roger (1935) reportaron este mismo hongo en Camerún asociado con esta misma especie de cochinilla harinosa. De igual manera, Pfaltzer y Fluiter (1941) reportaron en Indonesia a Polyporus coffeae Wakef. En la India se reportó a Diacanthodes philippinensis (Pat.) (Singer) asociado con Planococcus lilacinus (Cockerell, 1905) (Chacko & Sreedharan, 1981; Sekhar, 1964). En África, Watson y Cox (1990) reportaron a Planococcus citri (Risso, 1813) asociada con el hongo Diacanthodes novoguineesis (Hennings) Fidalgo; y en Papua Nueva Guinea, Williams (1986) reportó a Paraputo leveri (Green, 1934) asociado con el hongo D. philippinensis (Pat.) Singer.
Dentro de las especies de cochinillas harinosas registradas en raíces de café, Caballero et al. (2019) destacan a Pseudococcus elisae como una especie importante para el cultivo del café, con base en datos de distribución y frecuencia de ocurrencia. Pseudococcus elisae es una cochinilla harinosa nativa de la región neotropical; es polífaga y está registrada en especies de Annonaceae, Apocynaceae, Araceae, Blechnaceae, Boraginaceae, Convolvulaceae, Euphorbiaceae, Fabaceae, Musaceae, Piperaceae, Rubiaceae, Rutaceae y Zingiberaceae; sin embargo, se encuentra con mayor frecuencia en Musa paradisiaca L. (Musaceae) y en C. arabica, Coffea canephora Pierre ex A.Froehner, Coffea liberica W.Bull e Ixora L. (García Morales et al., 2016). En las Américas, P. elisae está establecida en Brasil, Chile, Colombia, Costa Rica, Cuba, Ecuador (territorio continental e Islas Galápagos), Estados Unidos, Guadalupe, Guatemala, Haití, Hawái, Honduras, Martinica, México, Panamá, Perú y Surinam (García Morales et al., 2016; Watson et al., 2022).
P. elisae tiene importancia económica debido a que en banano, junto con las especies del complejo D. texensis y otras especies de cochinillas harinosas transmiten el virus del estriado del banano (Banana Streak virus BSV), ocasionando pérdidas económicas (Armijos & Silva, 2010; Armijos et al., 2003). La biología de P. elisae ha sido poco estudiada, Armijos y Silva (2010) a una temperatura de 24 °C, reportaron una duración de huevo hasta adulto entre 54 y 93 días pasando por los estados de huevo con una duración de 12,4 días (rango entre 7 a 14 días) y una fertilidad del 75 %, ninfa (tres estados) con una duración de 8,95 días para la ninfa I; 9,98 días para la ninfa II y 12 días para la ninfa III. Por su parte, las hembras tienen un periodo de maduración de 8,9 días y el tiempo de oviposición es de 21 días durante el cual depositan 166 huevos en promedio; mientras que los machos pasan por los estados de ninfa I y II, prepupa y pupa y duran 2,5 días aproximadamente. De acuerdo con Beardsley (1986) las hembras de Pseudococcidae tienen en general cuatro estados y los machos cinco (incluidos los adultos). La hembra adulta pone los huevos en un saco ceroso llamado ovisaco adherido al hospedante y eclosionan en unas pocas horas o unos días y los primeros estados escapan del ovisaco y se arrastran sobre el hospedante para buscar un lugar adecuado para alimentarse, el primer estado es el más móvil. Los machos adultos no pueden alimentarse y normalmente no sobreviven más de un día y localizan a las hembras mediante feromonas sexuales (Beardsley, 1986).
Con base en lo anterior, para establecer las estrategias de manejo de una plaga agrícola, es necesario conocer la biología y su dinámica en campo. Por lo tanto, el objetivo de este trabajo fue evaluar el ciclo de vida de P. elisae y la dinámica en campo de especies de Pseudococcidae presentes en las raíces del café.
Materiales y métodos
Para la confirmación de la especie, se seleccionaron 15 hembras oviplenas y se montaron en láminas utilizando la metodología descrita por Sirisena et al. (2013), posteriormente se identificaron siguiendo las claves taxonómicas de Caballero et al. (2019) y Granara de Willink y González (2018).
Posteriormente se conformaron 100 unidades de observación, cada una, compuesta por una plántula de café de cuatro meses de edad, sembrada en un vaso desechable de 1000 ml (15,5 cm de alto × 11,5 cm de diámetro en la parte superior), con un disco de foami negro en la base del tallo que cubriera el suelo y permitiera ver la exuvia cuando la ninfa cambiara de estado, en la parte aérea de cada plántula con la ayuda de un pincel número cero, se colocaran cinco ninfas I. Las unidades de observación se ubicaron en bandejas metálicas y se mantuvieron en un cuarto climatizado. Diariamente, se evaluó el tiempo transcurrido entre las ninfas I y II, luego se seleccionó la primera ninfa que mudó en cada unidad de observación para continuar el ciclo, las restantes se eliminaron. Se realizaron observaciones diarias para determinar el momento que cambiaron de estado y se registró la duración en días. Como información complementaria se realizó la descripción morfológica de cada estado.
En C. moschata cada unidad de observación estuvo compuesta por una caja plástica igual a la descrita en el ítem anterior. En cada caja se dispuso un fruto de C. moschata en estado de maduración pintón con una hembra oviplena de P. elisae y en la base una servilleta humedecida con agua destilada. En ambos hospedantes, se realizaron observaciones diarias para determinar el momento de la oviposición; posteriormente, se realizó un conteo destructivo del número de huevos de cada ovisaco hasta la muerte de la hembra.
Para evaluar el parámetro de fecundidad se conformaron las mismas unidades de observación que en la evaluación de fertilidad. Por cada una se registró el número de ninfas emergidas por hembra yluego se realizaron observaciones hasta completar el ciclo de vida. Se contabilizó la proporción de machos y de hembras con base en la progenie total, duración de huevo a adulto (días), tiempo de pre-oviposición (días), tiempo de oviposición (días), número de huevos/hembra, número de ninfas emergidas/hembra y sobrevivencia del adulto.
Resultados
Ciclo de vida de Pseudococcus elisae sobre Coffea arabica en el laboratorio
La duración del ciclo de vida para las hembras desde huevo hasta adulto fue de 98,1 ± 1,3 días; se registraron tres estados ninfales con una duración de 61,4 ± 0,3 días. El tiempo de duración del estado de ninfa duró 9,3 veces más que el huevo. Las hembras pasaron por los estados huevo, ninfa (I, II y III) y adulto (Tabla 1). Para el caso de los machos la duración de huevo hasta adulto fue de 68 ± 0,4 días y pasó por los estados de huevo, ninfa (I y II), prepupa, pupa y adulto; la mayor duración se presentó en el estado de ninfa con 31,2 ± 0,5 días y el de menor duración el estado de huevo con 6,6 ± 0,3 días (Tabla 1).
Promedio y error estándar para la duración en días de los estados de desarrollo de Pseudococcus elisae sobre plantas de Coffea arabica en el laboratorio.
Ciclo de vida de Pseudococcus elisae sobre frutos de Cucurbita moschata en el labortatorio
La duración del ciclo de vida para las hembras desde huevo hasta adulto fue de 92,1 ± 0,3 días; se registraron tres estados ninfales con una duración de 60,1 ± 0,3 días. El tiempo de duración de los tres estados ninfales duró 9,2 veces más que el huevo. Las hembras pasaron por los estados de huevo, ninfa (I, II y III) y adulto (Tabla 2). Para el caso de los machos la duración de huevo hasta adulto fue de 67,2 ± 0,3 días y pasó por los estados de huevo, ninfa (I y II), prepupa, pupa y adulto. La mayor duración se presentó en el estado de ninfa (dos estados ninfales) con 32,4 ± 0,4 días y el de menor duración el estado de huevo con 6,7 ± 0,1 días (Tabla 2).
Promedio y error estándar para la duración en días de los estados de desarrollo de Pseudococcus elisae sobre frutos de Cucurbita moschata en el laboratorio.
Al comparar la duración de cada estado de P. elisae entre C. arabica y C. moschata; se encontró que la duración de huevo hasta adulto fue mayor en C. arabica y se diferenció estadísticamente de la duración en frutos de C. moschata; de la misma manera para ninfa I, ninfa III y adulto (Tabla 3).
Comparación estadística entre Coffea arabica y Cucurbita moschata para la duración en días de cada estado de desarrollo de las hembras de P. elisae.
*E.E. = Error estándar
*Letras no comunes indican diferencia entre promedios según prueba de Duncan al 5%.
Descripción morfológica de cada estado de desarrollo de Pseudococcus elisae

Estados de desarrollo de la hembra de Pseudococcus elisae. a. Huevo, b. Ninfa I, c. Ninfa II, d. Ninfa III, E. Hembra adulta (lente 1,6 × 1X)

Estados de desarrollo del macho de Pseudococcus elisae. a. Prepupa, b. Pupa, c. Macho, vista dorsal, d. Macho, vista ventral, e. Macho, vista lateral (lente 1,6 × 1X).
Parámetros de preoviposición, oviposición, fertilidad, fecundidad, razón sexual y sobrevivencia de Pseudococcus elisae
En Coffea arabica, cada hembra depositó en promedio 231 ± 3,6 huevos (fertilidad), de los cuales emergieron 197,3 ± 6,2 ninfas (fecundidad) lo que corresponde al 85,4 % de huevos eclosionados, la razón sexual fue de 11:6 a favor de las hembras (Tabla 4).
Promedios y error estándar para la duración en días de la preoviposición, oviposición, fertilidad y fecundidad, así como la razón sexual de Pseudococcus elisae sobre plántulas de Coffea arabica.
La tasa de supervivencia de P. elisae en C. arabica, fue alta en todas las etapas de desarrollo, tanto para los machos como para las hembras, y se mantuvo a través de todo el ciclo de vida. La supervivencia de las hembras en ninfa II fue del 98,8% y en ninfa III del 96,5%, el porcentaje de hembras que alcanzaron la etapa adulta fue de 87,0% (Figura 3a). Para los machos la supervivencia fue del 100% bajo las condiciones evaluadas (Figura 3b).
La curva de supervivencia para P. elisae (Figuras 3a y b) se asemeja a la curva de supervivencia Tipo I: Alta supervivencia (Rabinovich, 1980), donde la mayor parte de los organismos alcanzaron su máximo desarrollo.
En frutos de C. moschata cada hembra ovipositó en promedio 201 ± 3,6 huevos (fertilidad), de los cuales emergieron 182,7 ± 4,9 ninfas (fecundidad) lo que corresponde al 90,9 % de huevos eclosionados, la razón sexual fue de 8:8 (Tabla 5).
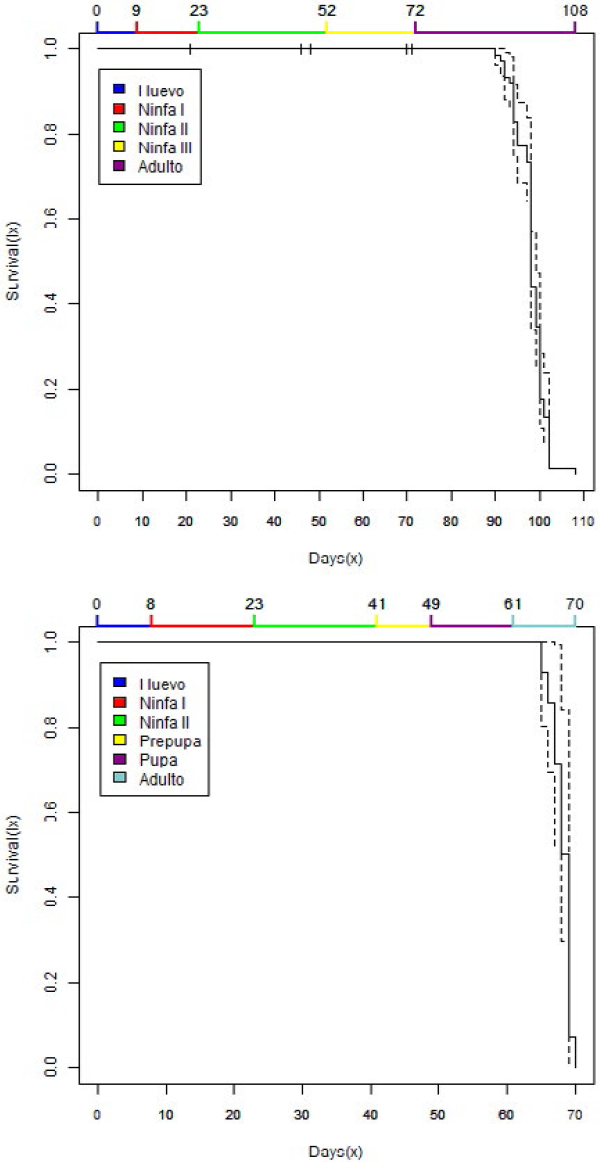
Análisis de supervivencia Kaplan - Meier para hembras (3a) y machos (3b) de Pseudococcus elisae sobre plantas de Coffea arabica.
Promedios y error estándar para la duración en días de la preoviposición, oviposición, fertilidad y fecundidad, así como la razón sexual de Pseudococcus elisae sobre frutos de Cucurbita moschata.
La tasa de supervivencia de P. elisae en frutos de C. moschata fue alta en todas las etapas de desarrollo tanto para los machos como para las hembras y se mantuvo a través de todo el ciclo de vida. La supervivencia de las hembras en ninfa II fue del 98,6% y en ninfa III del 95,8%, el porcentaje de hembras que alcanzaron la etapa adulta fue de 88,6% (Figura 4a). Para los machos la supervivencia fue del 90,9% bajo las condiciones evaluadas (Figura 4b). La curva de supervivencia para P. elisae en frutos de C. moschata (Figuras 4a y 4b) se asemeja a la curva de supervivencia Tipo I: Alta supervivencia (Rabinovich, 1980), donde la mayor parte de los organismos alcanzaron su máximo desarrollo.
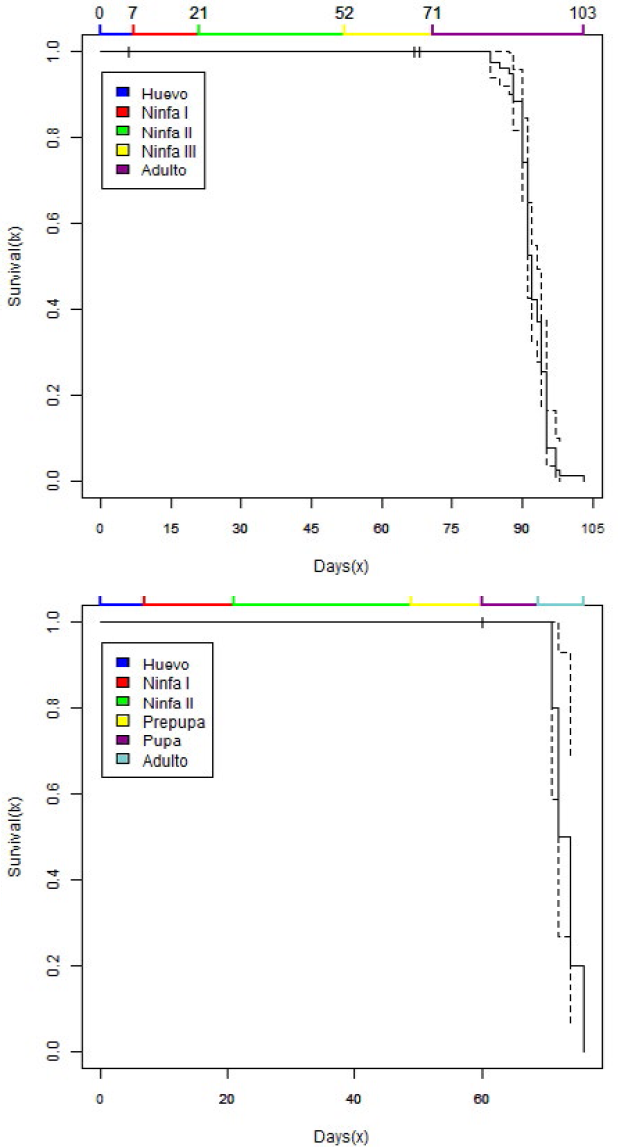
Análisis de supervivencia Kaplan - Meier para hembras (4a) y machos (4b) de Pseudococcus elisae sobre plantas de Cucurbita moschata.
Al comparar la duración en días de la preoviposición, oviposición, fertilidad, fecundidad y sobrevivencia de P. elisae (Tabla 6), las hembras de P. elisae ovipositaron mayor número de huevos en C. arabica que en frutos de C. moschata siendo diferentes estadísticamente; también se presentó mayor emergencia de ninfas en C. arabica.
Comparación estadística entre Coffea arabica y frutos de Cucurbita moschata para los parámetros preoviposición, oviposición, fertilidad, y fecundidad de P. elisae.
*E.E. = Error estándar
*Letras no comunes indican diferencia entre promedios según prueba de Duncan al 5%.
Dinámica en campo de especies de Pseudococcidae presentes en las raíces del café
Se evaluaron 395 plantas de café durante 14 evaluaciones, de las cuales 278 (70,4%) tenían cochinillas harinosas; de estas, el 82,7% presentaron P. elisae y especies del complejo D. texensis. Se encontraron individuos de P. elisae desde la primera evaluación, la presencia de cochinillas harinosas se mantuvo a lo largo de todas las evaluaciones y el número de individuos fue variable en cada evaluación (Figura 5). En las 278 plantas se encontraron en total 11.323 individuos.
Las especies P. elisae y complejo D. texensis se encontraron distribuidas aleatoriamente en el lote; sin embargo, se observaron sitios de mayor concentración (Figura 6). Fue frecuente encontrar en una misma raíz dos especies de Coccomorpha: complejo D. texensis y P. elisae; complejo D. texensis y Puto barberi; P. elisae y P. barberi. Hasta el momento se desconoce que P. barberi se asocie con hongos basidiomicetos y enquiste raíces del café; sin embargo, se encontró frecuentemente en el lote evaluado.
El análisis descriptivo para la variable número de individuos por planta, indicó que P. elisae presentó mayor población por planta, con una mediana de 24,5 individuos frente a 20,5 para el complejo D. texensis (Figura 7).
A partir de los 11 meses de edad se encontraron plantas con presencia de micelio de los hongos que se asocian con las cochinillas harinosas que enquistan raíces del café, es decir, enquistadas (Figura 8). En total se registraron 10 plantas con presencia de cochinillas harinosas y nódulos del hongo que cubrían las raíces principales y secundarias (Tabla 7) y contenían en su interior ninfas y adultos de P. elisae y el complejo D. texensis. Con base en el índice de disponibilidad hídrica (IDH) de la estación meteorológica cercana al sitio de estudio se encontró que, para la primera década del mes de agosto hubo déficit hídrico en la zona (Figura 9) que coincidió con elmomento en el cual se registraron las primeras raíces enquistadas con nódulos recién formados.
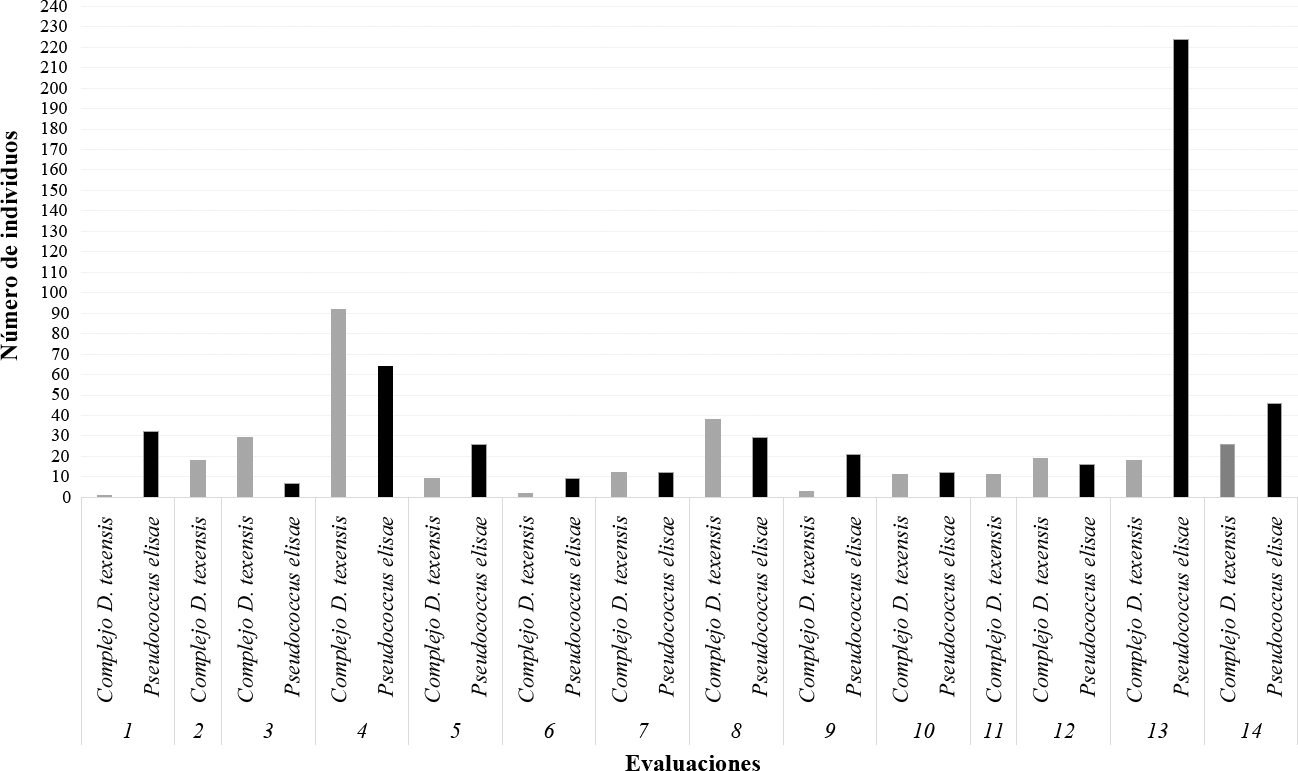
Mediana del número de individuos por especie y evaluación.

Distribución en el lote de las especies de cochinillas harinosas que enquistan raíces del café Pseudococcus elisae y complejo Dysmicoccus texensis, así como P. barberi.
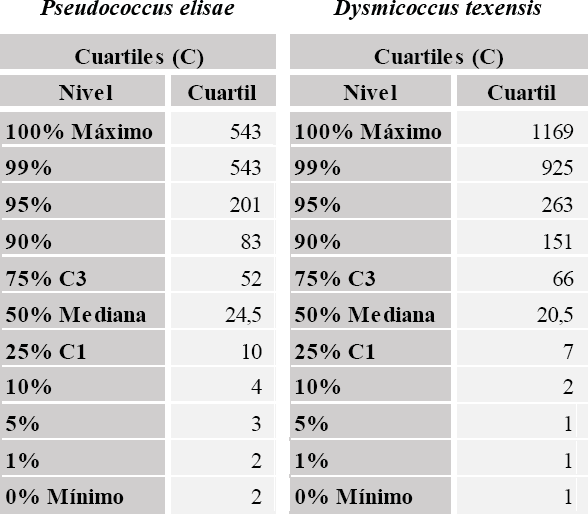
Análisis descriptivo para el número de individuos por planta de Pseudococcus elisae y complejo Dysmicoccus texensis.

Raíz de planta de Coffea arabica enquistada, con presencia de una especie del complejo Dysmicoccus texensis.
Número de plantas enquistadas en cada evaluación, especie de cochinilla harinosa, número de individuos, número de nódulos y promedio de individuos encontrados en cada nódulo.
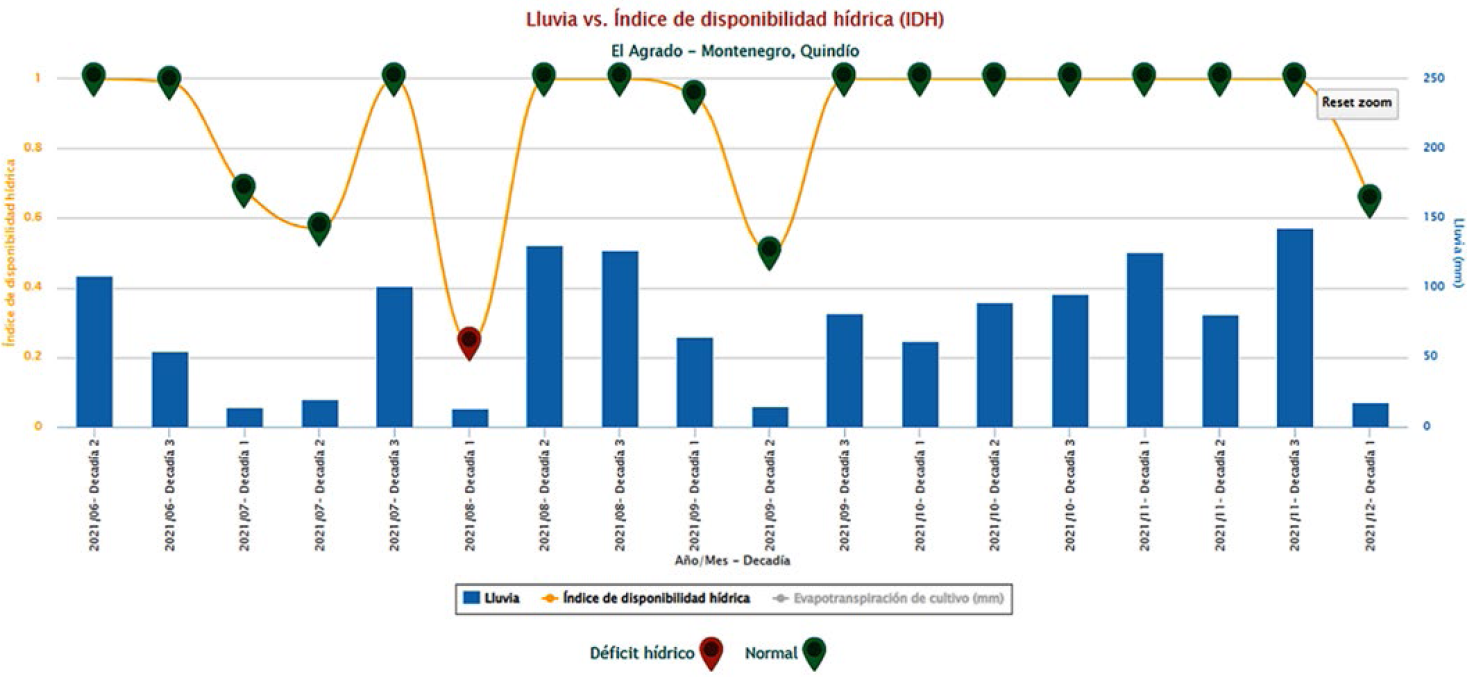
Índice de disponibilidad hídrica para la estación meteorológica el Agrado, Montenegro, Quindío.
En cuanto al efecto de la presencia de cochinillas harinosas en el número de hojas por planta y número de raíces secundarias, se encontró que, en las plantas con presencia de cochinillas harinosas y quistes el número de raíces secundarias disminuyó con respecto a las plantas sanas (Figura 10). Sin embargo, no se diferenciaron estadísticamente y para la variable número de hojas no se encontraron diferencias.
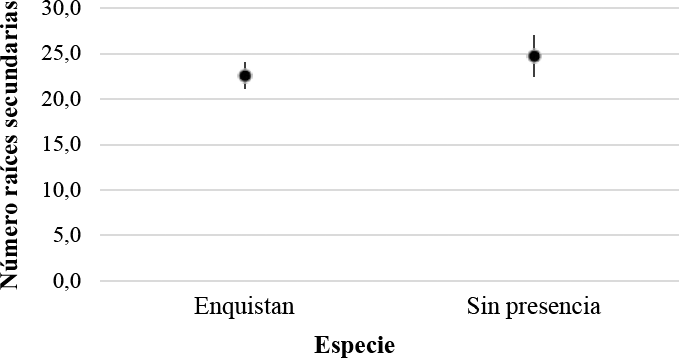
Análisis descriptivo de la variable número de raíces secundarias en plantas con raíces enquistadas y plantas sanas, es decir sin presencia de cochinillas harinosas ni quistes.
Discusión
Sobre el ciclo de vida de P. elisae, los pocos estudios existentes reportaron a una temperatura de 24 °C, una duración de huevo hasta adulto entre 54 y 93 días pasando por los estados de huevo, ninfa (tres estados), adulto macho y hembra. Los machos pasan por los estados de prepupa y pupa y este estado presentó una duración entre cinco y ocho días (Armijos & Silva, 2010). En el presente estudio bajo condiciones controladas de temperatura de 25 °C ± 2 °C, y humedad relativa (HR) entre 70% y 80%, encontramos que una hembra de P. elisae completa su ciclo de vida entre 92 y 98 días dependiendo del hospedante, lo que concuerda con Armijos y Silva (2010); sin embargo, difiere de la duración registrada para otras especies de Pseudococcus como Pseudococcus viburni (Signoret, 1875) sobre hojas de manzana [Malus domestica (Suckow) Borkh (Rosaceae)], hojas de uva [Vitis vinifera L. (Vitaceae)] y caqui [Diospyros kaki Thunb. (Ebenaceae)] en donde reportaron desde 45 hasta 47 días a 25 °C (Mudavanhu, 2009). Lo anterior se puede deber a las variaciones en las características morfológicas, químicas y fisiológicas de los hospedantes, así como los valores nutricionales y los niveles de nitrógeno de las plantas, los cuales están relacionados con el desarrollo de los insectos (Cocco et al., 2014; Schoonhoven et al., 2005).
El ciclo de vida de las especies de Pseudococcus puede ser más largo que las especies de Dysmicoccus; por ejemplo, Alves (2006) reportó para D. texensis a 27 °C un ciclo total de 60 ±13,2 días para las hembras pasando por tres estados ninfales; mientras que Garay y Mora (2018), registraron que el ciclo de vida total a 25 °C ± 1 °C y 80% HR fue de 56,3 ± 2,2 días, lo que puede conllevar a menos generaciones por año. Nuestros resultados indican que a 25 °C ± 2°C, 70% - 80% HR y fotoperiodo 12:12 h [L:D], P. elisae podría completar de 3 a 4 generaciones en un año, pero se deben realizar más estudios para evaluar el efecto de la temperatura y humedad relativa en la biología de P. elisae.
En el presente trabajo, los resultados en cuanto al número de huevos por hembra (fertilidad) de P. elisae, son consistentes con lo reportado para hembras de la familia Pseudococcidae por Pacheco da Silva et al. (2017) en la especie P. viburni a 25 °C ± 1 °C y 70% ± 10% HR que encontraron que el número medio de huevos por hembra fue de aproximadamente 88, alcanzando un máximo de 201. Así mismo, Bettencourt y Simões (2008) y Oyarzun (2004), para la misma especie, reportaron desde 70 hasta 300 huevos en papas germinadas; mientras tanto, Mudavanhu (2009), a 25 °C reportó 240 huevos en plantas de manzana. En el caso de Phenacoccus manihoti Matile-Ferrero, 1977 (Pseudococcidae) sobre plantas de yuca [Manihot esculenta Crantz (Euphorbiaceae)], Barilli et al. (2014), a 25 °C ± 2 °C y un fotoperiodo de 14 h obtuvieron desde 37 hasta 497 huevos por hembra, con un promedio de 247,1. Lo anterior parece indicar que la capacidad de oviposición de las hembras de diferentes especies de la familia Pseudococcidae se encuentra estrechamente relacionada con las condiciones de temperatura y plantas hospedantes. En cuanto a la fecundidad, en este caso, sobre ambos hospedantes, P. elisae tuvo una tasa alta con más del 90% de huevos eclosionados por hembra.
La supervivencia de las hembras de P. elisae registrada en este trabajo fue alta y comenzó a disminuir drásticamente después de las etapas reproductivas. En el caso de los machos, la longevidad fue mayor que lo registrado por otros autores como Costa et al. (2016) para Planococcus citri (Risso, 1813), estos autores indicaron longevidades de 36,5 + 1,2 días a 25 C° ± 1 C°, 70 % ± 10% de HR y 12 horas de fotoperiodo y para Planococcus minor (Maskell, 1897) de 37,1+1,8 días bajo las mismas condiciones. Así mismo, Garay y Mora (2018) registraron promedios altos de longevidad a 25 °C en las hembras de D. texensis desarrolladas sobre tubérculos de papa.
Bajo las condiciones de este estudio, se encontró que P. elisae y especies del complejo D. texensis comenzaron a presentar la asociación con los hongos basidiomicetos a partir del mes 11 de establecido el lote; el hongo se observa como una capa corchosa (micelio) que crece y se extiende sobre la raíz principal y las raíces secundarias, cubriendo a los individuos de los pseudocóccidos que se están alimentando de las raíces; de esta manera, el hongo les proporciona protección de los enemigos naturales y de condiciones ambientales adversas (Fang et al., 2020).
Conclusiones
Pseudococcus elisae es una especie con estrategia r por su tamaño pequeño, vida corta, respuesta muy rápida, y la estrategia de supervivencia es multiplicarse y crecer muy rápidamente. Con base en la literatura esta plaga se desarrolla y reproduce exitosamente sobre varios hospedantes; en este estudio se encontró que en C. arabica y C. moschata presentó altas tasas de supervivencia y longevidad, esto sugiere que es una amenaza para los cultivos de café del país, ya que, si las condiciones ecológicas son adecuadas, una hembra alcanza la adultez y madurez reproductiva en 90 días aproximadamente y puede producir más de 200 huevos, que eclosionan en un período de seis días. El conocimiento del ciclo de vida de P. elisae es útil para planificar estrategias de manejo, como el número y el período de las aplicaciones de productos de síntesis química.
Origen y financiamiento
Este estudio hizo parte del proyecto de Investigación ENT103004, financiado con recursos de la Federación Nacional de Cafeteros de Colombia (FNC)-Cenicafé.
Contribución de los autores
Luz Andrea Carmona-Valencia, realizó el trabajo de campo y laboratorio, procesamiento de datos, análisis de la información y redacción del manuscrito. Pablo Benavides-Machado, contribuyó en la construcción del proyecto de investigación, realizó ajustes a la metodología, seguimiento al registro de la información y revisión del manuscrito. Rubén Darío Medina contribuyó en los análisis estadísticos. Zulma Nancy Gil-Palacio. Fue responsable de la construcción del proyecto de investigación, la dirección, diseño y supervisión del trabajo experimental, recolección de las muestras en el campo y revisión y ajustes del manuscrito.
Conflicto de intereses
Los autores declaran no tener ningún conflicto de intereses.
Footnotes
Agradecimientos
Agradecemos la colaboración de Fáber de los Ríos, Mauricio Jiménez, Robín García, Luis Eduardo Escobar, Daniel Antonio Franco y el personal de la Estación Experimental Paraguaicito por el apoyo en la fase campo. Así mismo, extender nuestro agradecimiento a Carlos Augusto Ramírez, Luisa Fernanda Monsalve y Nancy Arciniegas por su colaboración con los procedimientos de laboratorio.