
Abstract
The coffee berry borer Hipothenemus hampei (Ferrari) is the most serious insect pest of the coffee plantations. Among the biological control methods the fungus Beauveria bassiana (Bals.) Vuill. may play an important role in borer population reduction. In this research a bioassay method was standardized to test the pathogenicity of B. bassiana on H. hampei. Adults of the borer were obtained from a laboratory mass rearing using parchment coffee at 45% R.H. The borers were desinfested with sodium hypochlorite at 0,5% for 10 minutes and rinsing the excess in three washes with sterile distilled water. The experiment was organized in a complete randomized desing using three treatments formed by isolate Cenicafe Bb-9205 from Diatraea saccharalis (Bb-9205 DS), the same isolate passed through the borer (Bb-9205 BFC) and a control. Thirty adults of the borer were used per replication. These insects were exposed to the fungus using a concentration of 1 x 107 conidia/ml for two minutes and then transferred individually in glass vials (4 cc of capacity) with a piece of wet paper inside and capped with absorbent cotton. Twenty four hours later a berry of waxed-parchment coffee with 45% of humidity was added as a substrate. Infected insects showed a reduced activity, lost of legs and antennae and therefore inability to bore the coffee grain. Treated insects with the fungus resulted in lower grain attack (14.44%) compared with control insects (44.44%). There were significant differences (P≥0.05) among treatments, average mortality was 88.88% for isolate Bb-9205 DS and 100% for Bb-9205 BFC. Mean lethal time (TL50) was 92.4 hours with isolate Bb-9205 DS and 54.72 hours with Bb-9205 BFC. Five stages of development of B. bassiana on H. hampei were recognized: inoculation to death, death to initial mycelial growth, death to micelial coverage, death to conidiogenesis and death to discharge of conidia. These stages lasted 20.16; 31.92; 56.40; 83.28 and 198 hours for Bb-9205 DS and 20.88; 44.40; 66.88; 116 and 182 hours for Bb-9205 BFC. This biossay can be used as a model to evaluate different entomopathogens to be tested against H. hampei.
Introducción
En la actualidad, la broca del fruto del café, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae), es el mayor enemigo del cultivo del café en Colombia. La broca está considerada como la plaga insectil que mayor daño causa al cultivo del café, porque su ataque se dirige a la almendra o endospermo donde puede reproducirse (Decazy 1990). Los daños causados por la broca estan representados en la pérdida de peso de las almendras al ser atacadas por las larvas, caída de los frutos en estado acuoso y alteración en la calidad de la bebida al beneficiar conjuntamente granos sanos y brocados que además son contaminados por microorganismos (Cárdenas 1991). Esta plaga fué registrada por primera vez en Colombia en el año de 1988, en la frontera con el Ecuador, cerca a Tumaco (Nar.). En la actualidad se encuentra ampliamente distribuída en los departamentos de Antioquia, Caldas, Cauca, Cundinamarca, Huila, Putumayo, Quindío, Risaralda, Tolima y Valle, infestando cerca de 100.000 hectáreas.
La estrategia generalizada para el manejo de las poblaciones de la broca del café en los países donde a llegado, se basa en el control químico, el cual causa mortalidad inmediata sobre las poblaciones de broca, pero tiene efectos negativos sobre el agroecosistema y sobre la salud de la vida animal, lo que amerita la búsqueda de otras alternativas de control como pueden ser, el uso de parasitoides o de organismos entomopatógenos.
El hongo Beauveria bassiana (Bals.) Vuill (Hyphomycetos) es uno de los agentes de control biológico más importantes para la represión de muchas plagas (Ferron 1978; Evans 1982) y se considera que puede llegar a ser muy eficiente en programas de control de la broca del café (Bustillo 1991a). Sin embargo, no existe hasta el momento una metodología confiable para evaluar este hongo bajo condiciones de laboratorio.
Para la evaluación de un entomopatógeno sobre un huésped dado se requiere desarrollar una metodología de bioensayo confiable y reproducible que garantice el control del insecto plaga. La patogenicidad de un entomopatógeno puede evaluarse bajo condiciones de laboratorio empleando insectos susceptibles y un método de cría que permita la sobrevivencia y la expresión de los síntomas y signos de la enfermedad causada por el hongo a evaluar. El método de bioensayo a probar debe permitir estimar la mortalidad, la distribución de la mortalidad diaria, el tiempo letal medio (TL50) y el ciclo de desarrollo del patógeno sobre el insecto huésped. Esta investigación tuvo por objetivo determinar estos parámetros mediante la evaluación de la patogenicidad de B. bassiana cepa Bb-9205 aislada directamente de larvas de Diatraea saccharalis (Fabricius) (Lepidoptera: Pyralidae), la cual se comparó con la obtenida una vez se reactivó sobre la broca del café.
Revisión de Literatura
La broca del café, H. hampei, es considerada, después de la enfermedad de la cereza del café (CBD), como la plaga más importante del cultivo, debido a su hábito de atacar directamente la cereza del café, lo cual hace difícil su control (Muller 1990). Se considera que el control de la broca se debe realizar a través de un programa de manejo integrado conformado por varios componentes de control biológico como son el hongo B. bassiana y los parasitoides Cephalonomia stephanoderis Betrem y Prorops nasuta Waterston (Hymenoptera: Bethylidae) (Moore y Prior 1988; Bustillo 1991b).
El B. bassina es uno de los entomopatógenos más estudiados para el control biológico de muchas plagas (Roberts y Yendol 1971). Este hongo se ha encontrado atacando al H. hmpei en los sitios de origen del café, en los países a donde este ha sido llevado y donde se ha introducido la broca (Bustillo 1991a). Los primeros registros de la incidencia del hongo sobre la broca fueron realizados en el Congo Belga en la década del 30 (Pascalet 1939; Steyaert 1935).
La incidencia del hongo B. bassiana sobre la broca del café, bajo condiciones de campo, varía de un país a otro; por ejemplo, en Costa de Marfil y en Kenya, las infecciones han sido raras (Ticheler 1963; Murphy y Moore 1990), pero se han registrado altos niveles en Camerún, Honduras, México y Ecuador (Pascalet 1939; Tronconi et al. 1986; Barrera et al. 1990; Klein et al. 1988). Estas diferencias se pueden deber a factores climáticos o a que la broca está mejor adaptada al hongo en su sitio de origen en el Africa, pero es susceptible a los aislamientos que encuentra en los nuevos sitios que coloniza (Prior 1987).
El B. bassiana causa epizootias en poblaciones de broca cuando las condiciones ambientales son favorables, pero estas infecciones naturales muchas veces no son suficientes para detener el avance de la plaga. Las condiciones de vida de la broca del café la hacen naturalmente susceptible a la infección por el hongo, ya que tanto este como la broca se desarrollan óptimamente bajo condiciones de alta humedad (Moore y Prior 1988).
Resultados de estudios de laboratorio y de campo indican que el hongo podría utilizarse eficientemente en la reducción de las poblaciones de broca al depositar suficiente inóculo para inducir o acelerar el proceso de infección en el campo (Jiménez 1992; Tronconi et al. 1986; Fernández et al. 1985; Bustillo et al. 1991). Sin embargo, el comportamiento del hongo puede ser variable según las condiciones de temperatura, humedad y radiación solar presentes en el agroecosistema (Ferron 1978; Ignoffo 1975). La temperatura óptima está entre 20 y 30°C, y las humedades superiores al 90% son más favorables. La precipitación debe mantener una humedad suficiente para una buena esporulación (Pascalet 1939; Alves 1986b).
El hongo B. bassiana puede infectar a la broca independientemente de la variedad de café. Villacorta (1984), en Brasil, encontró infecciones tanto en frutos de café robusta como de Coffea arabica L. var. Mundo novo. El efecto del hongo es más visible sobre frutos verdes, en los cuales se está iniciando la penetración de la broca, debido a que esta queda más expuesta a la infección (Pascalet 1939). La germinación de las conidias de B. bassiana ocurre en un período de 12 horas después de la inoculación. El hongo penetra a través del integumento por acción mecánica y efectos enzimáticos, lo cual toma otras 12 horas. Después de unas 72 horas de la inoculación, el insecto está totalmente colonizado. La duración de las diferentes fases de este ciclo depende de la especie atacada y de las condiciones ambientales presentes durante la infección (Alves 1986a).
La habilidad de un hongo entomopatógeno para sobreponerse a los mecanismos de defensa de sus huéspedes se debe en gran parte a la producción de toxinas, lo cual constituye uno de los componentes principales de la patogenicidad y uno de los más difíciles de establecer. La muerte causada por toxinas puede ser considerada cuando los insectos mueren rápidamente después de la infección y a menudo no muestran evidencia de una infección fungal extensiva.
Las enzimas extracelulares o metabolitos secundarios como las lecitinas, coagulasas, hialuronidasas son otro tipo de sustancias frecuentemente encontradas en ciertos microorganismos que contribuyen a la patogenicidad del hongo (Khachatourians 1991). El hongo B. bassiana presenta variaciones en su eficiencia para atacar un determinado insecto. Recientemente, Jiménez (1992) comprobó que existen diferencias en virulencia entre las razas de B. bassiana a la broca del café. Bajo condiciones de laboratorio, este autor evaluó 46 aislamientos provenientes de diferentes huéspedes y de diferentes sitios de origen, y encontró que el 35% de ellos causó mortalidades inferiores al 50% y cinco de ellos alcanzaron niveles de infección superiores al 80%. Igualmente, Paccola y Azevedo (1990) encontraron variabilidad natural del hongo B. bassiana cuando fue aislado de diferentes huéspedes. El aislamiento obtenido de larvas de D. saccharalis se caracterizó como potencialmente patogénico hacia insectos plagas debido a los patrones enzimáticos lipasa, amilasa y proteasa que le confieren mayor patogenicidad. Las diferencias en patogenicidad podrían deberse a respuestas internas del huésped, y por parte del hongo a características fisiológicas y genéticas (Ignoffo 1975).
La patogenicidad de un microorganismo y el desarrollo de la infección son el resultado de un número de componentes tanto del patógeno como del huésped, tales como la rápida germinación, la velocidad de reproducción, la tasa de esporulación, el tiempo de exposición, la producción de toxinas y el estado fisiológico del huésped (Kachatourians 1991).
Los antecedentes expuestos permiten inferir que el hongo B. bassiana puede ser considerado como un componente promisorio en el manejo integrado de la broca del café debido a las ventajas que ofrece, como son: su fácil manejo, la rapidez con que se colonizan sus huéspedes y, principalmente, por la preservación del ecosistema cafetero.
El desarrollo de un bioplaguicida requiere, inicialmente, la selección de razas o biotipos altamente patogénicos hacia el insecto a controlar, para lo cual se debe establecer un sistema de bioensayo adecuado (Hall y Papierok 1982). Por lo tanto se hace necesario evaluar mediante bioensayos el potencial patogénico de los aislamientos de B. bassiana con que cuenta CENICAFE, para asegurar el uso del mejor material en el control de la broca del café bajo condiciones de campo.
Materiales Y Métodos
Los especímenes de H.hampei utilizados en el experimento fueron obtenidos de la unidad de cría de parasitoides que tiene CENICAFE, en Ansermanuevo (Valle). La broca se crió sobre café pergamino con un 45% de humedad, previamente tratado con fungicidas y recubierto con cera natural. Posteriormente, los insectos se desinfectaron por inmersión en una solución de hipoclorito de sodio al 0,5% durante 10 minutos y tres enjuagues con agua destilada estéril. Se seleccionaron los especímenes más activos, completamente sanos, negros o melaminizados y con una edad inferior a 8 días, en promedio, para garantizar la homogeneidad del material biológico utilizado.
Del cepario de B. bassiana del laboratorio de entomopatógenos de CENICAFE se tomó, como inóculo para este trabajo, el denominado Bb-9205 aislado de larvas de D. saccharalis colectadas en caña de azúcar en Candelaria (Valle).
El experimento se organizó en un diseño completamente al azar. Se evaluaron tres tratamientos: el primero con el aislamiento Cenicafé Bb-9205 aislado directamente de D. saccharalis que en adelante se denominará Bb-9205 DS, el segundo tratamiento fue la misma cepa pasada por broca que se denominará Bb-9205 BFC y el tercero el testigo. Cada tratamiento tuvo tres repeticiones y se emplearon 30 insectos por repetición.
Los aislamientos se purificaron en el medio de cultivo Sabouraud Dextrosa Agar (SDA), en cajas de petri de 9 cm de diámetro. La edad de los cultivos fue inferior a 30 días.
El inóculo se preparó suspendiendo el hongo, proveniente de tubos de ensayo, en 10 ml de agua destilada estéril y dos gotas de Tween 80 al 0,1%. La concentración del inóculo se determinó con un hemocitómetro, ajustándola posteriormente a 1 x 107 conidias/ml. La viabilidad del inóculo se determinó antes de los bioensayos, evaluando la germinación de las conidias a las 24 y 48 horas en cajas de petri con SDA acidificado e incubadas a 25 ±1°C.
Para realizar el experimento, las brocas se desinfectaron previamente. Primero se tomaron las brocas del testigo, separando un grupo de 90 individuos al azar, los cuales se sumergieron por dos minutos en agua destilada estéril y se dispusieron en forma individual dentro de frascos de vidrio transparente de 2 cm de diámetro y 4 cm de altura que contenían un disco de papel de filtro húmedo. Los frascos se taparon con un tapón de algodón, bien ajustado, para impedir la salida del insecto y mantener las condiciones de humedad.
La metodología empleada para la inoculación de los tratamientos consistió en colocar en inmersión el número total de brocas (tratamientos) en una caja de petri que contenía la suspensión 1 x 107 conidias/ml y se agitó por dos minutos. Cumplido el tiempo de exposición se retiró el exceso de inóculo de las brocas invirtiendo la caja de petri sobre un papel toalla seco. Posteriormente, con la ayuda de un pincel, los insectos se colocaron individualmente en los frascos y se distribuyeron en tres repeticiones por tratamiento, de 30 individuos cada una. Cada repetición fue confinada dentro de cajas de galletas con una mota de algodón húmedo, para crear una atmósfera saturada de humedad propicia para desencadenar el proceso de la patogénesis. Venticuatro horas más tarde se adicionó a cada uno de los frascos un grano de café pergamino seco, como se describió inicialmente, el cual sirvió como sustrato alimenticio.
Para proveer la humedad adecuada en el interior de cada uno de los frascos, diariamente, y con una jeringa estéril, se adicionaron 0,2 ml de agua destilada estéril. Los tratamientos y el testigo fueron mantenidos bajo condiciones de luz alterna, a una temperatura de 25 ±1°C y una humedad relativa superior al 90%.
La mortalidad de las brocas se evaluó diariamente durante 10 días después de la inoculación, utilizando para ello un estereoscopio que permitía realizar la disección de los granos sin causar daño a la broca. Se registró la mortalidad total y la mortalidad causada por el hongo. Con los datos obtenidos se estimó el porcentaje de mortalidad, la distribución de la mortalidad diaria, el tiempo letal medio (TL50) y las etapas de desarrollo del hongo sobre la broca.
Resultados Y Discusión
Las pruebas de viabilidad realizadas a las suspensiones de conidias dieron porcentajes de germinación superiores al 80%, lo cual indica la buena calidad biológica del patógeno evaluado en el experimento. La calidad del inóculo se relaciona directamente con la viabilidad de las conidias y con el tiempo de sobrevivencia del hongo en el medio. Diferencias en la viabilidad de las conidias generan variabilidad en las respuestas, las cuales podrían ser atribuídas a diferencias de patogenicidad (Lazo 1990).
La práctica de desinfección superficial de las brocas mediante el tratamiento con hipoclorito de sodio al 0,5% durante 10 minutos, no ocasionó mortalidad al insecto ni inhibió el proceso de germinación de las conidlas en los tratamientos. Esto concuerda con lo registrado por Jiménez (1992), quien obtuvo una sobrevivencia del 80% en el testigo al efectuar desinfestación de los granos con hipoclorito de sodio al 5% durante 5 minutos. El bioensayo permitió la sobrevivencia de las brocas por un tiempo superior a 10 días, suficiente para la evaluación del experimento y para la expresión de los síntomas y signos de la enfermedad en los tratamientos.
Al cabo de 24 horas se empezaron a observar los primeros síntomas de la enfermedad en las brocas tratadas con el hongo B. bassiana. Se observó una menor actividad y pérdida de apéndices como patas y antenas, así como una disminución en la capacidad de brocado. En los tratamientos con B. bassiana, el porcentaje de brocado en los granos pergamino fue de 14,44%, mientras que en el testigo alcanzó un nivel de 44,44%, lo que significa que las brocas infectadas por el hongo tienen una menor capacidad de causar daño y de penetrar en los granos.
En la patogenicidad del aislamiento Bb-9205 se encontraron diferencias significativas (P≥0,05) antes y después de pasar el hongo por la broca al igual que en el testigo. El porcentaje de mortalidad causado por el aislamiento Bb-9205 DS fue de 88,88%, y para el aislamiento Bb-9205 BFC fue del 100%. En el testigo no se presentó mortalidad por el hongo, ni por contaminantes. La mortalidad atribuída a factores de manipulación fue de 3,33% en el testigo y de 11,11% para el aislamiento Bb-9205 DS (Tabla 1). Entre las posibles causas de mortalidad por otros factores se identificó la manipulación de las brocas, el escape de los recipientes y causas fisiológicas como la inanición.
Mortalidad promedia de H. hampei causada por B. bassiana
Promedios seguidos por la misma letra no presentan diferencia significativa. (Prueba de Duncan P≤0,05)
El porcentaje de mortalidad con el aislamiento Bb-9205 BFC fue superior en 11,12%, comparado con el Bb-9205 DS, lo cual puede atribuirse a que el patógeno adquirió mecanismos de agresividad al pasar por el huésped susceptible (Khachatourians 1991).
La mortalidad diaria (Fig. 1) presentó una distribución variable en la población sometida a los tratamientos en estudio. Para el aislamiento Bb-9205 DS, la mortalidad de las brocas se inició después de las 12 horas de la inoculación, el mayor porcentaje (36,25%) se presentó a las 108 horas. La mortalidad se registró aún 228 horas después de la inoculación. Para el Bb-9205 BFC, la mortalidad se inició antes de 12 horas y el mayor porcentaje de mortalidad (56,66%) se obtuvo a las 60 horas y esta se extendió hasta las 132 horas después de la inoculación. El aislamiento Bb-9205 BFC mató más rápídamente a H. hampei como se observa en la Figura 1.
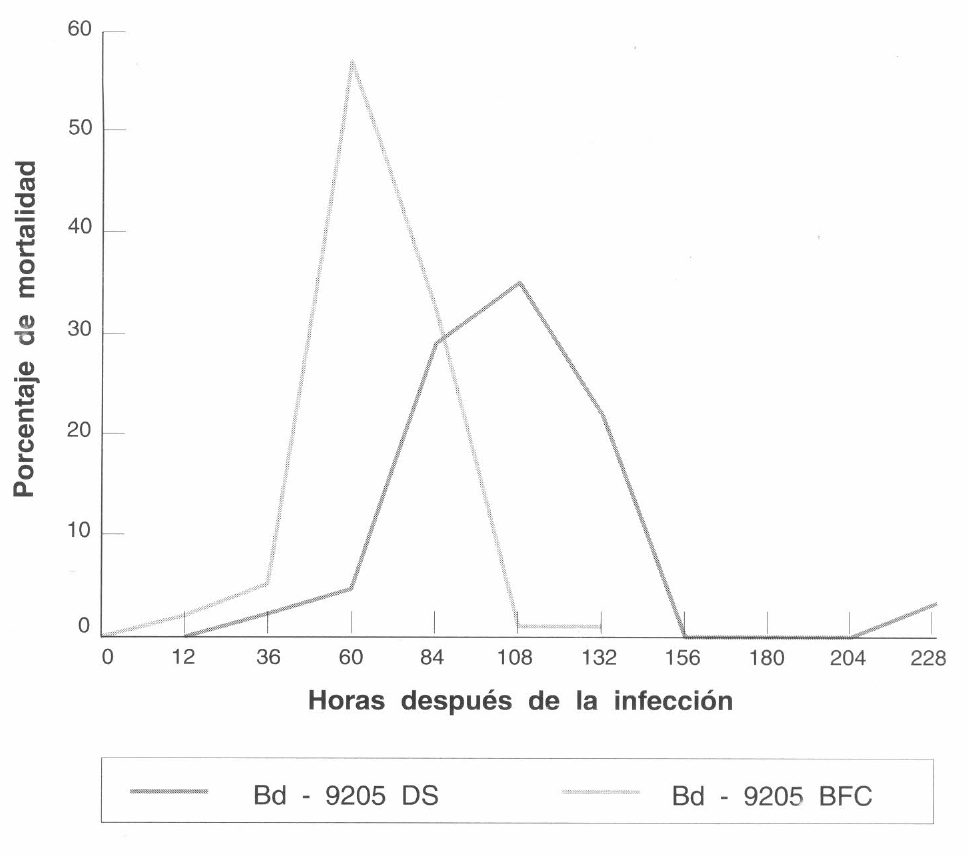
Distribución de la mortalidad diaria de Hypothenemus hampei causada por el aislamiento de Beauveria bassiana (Bb-9205) obtenido sobre larvas de D. saccharalis y activado en broca.
El tiempo letal medio (TL50) varió en los tratamientos (Fig. 2), siendo más corto con el aislamiento Bb-9205 BFC (54,7 horas), mientras para el aislamiento Bb-9205 DS fue de 92,4 horas. La disminución en el tiempo de mortalidad de las brocas tratadas con Bb-9205 BFC puede ser debida a la reactividad de la cepa al pasarla por el huésped susceptible. La disminución en el tiempo de mortalidad diaria y en el tiempo letal medio (TL50) puede estar indicando una mayor agresividad del hongo hacia el huésped susceptible y es un parámetro importante cuando se requiere establecer el patógeno bajo condiciones de campo.
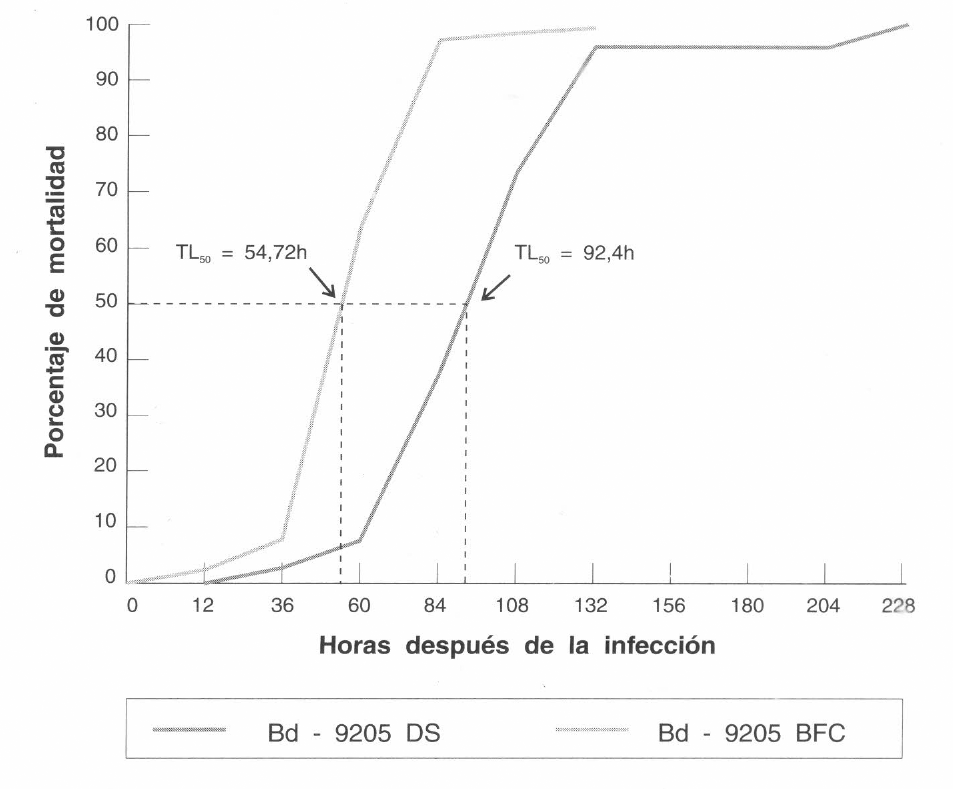
Mortalidad acumulada y tiempo letal medio (TL50) de la broca del café H. hampei infectada con el aislamiento Cenicafé Bb-9205 de Beauveria bassiana.
Los resultados de este estudio permiten concluir que aislamientos del hongo que sean activados sobre el huésped al cual se van a dirigir, son más patogénicos que cuando no se pasan a través de este. También es importante destacar cómo aislamientos provenientes de otros insectos pueden ser potencialmente más activos contra insectos de otros órdenes. Lo anterior ha sido documentado por otros investigadores (Jiménez 1992; Paccola y Azevedo 1990; Moore y Prior 1988).
En el ciclo de desarrollo del hongo sobre la broca se identificaron cinco etapas: Inoculación a muerte, muerte a inicio de micelio, muerte a cubrimiento de micelio, muerte a conidiogénesis y muerte a liberación de conidias (Tabla 2). Estas etapas expresan los signos de la enfermedad, los cuales se caracterizaron por el aspecto y el tiempo de duración.
Duración de las diferentes etapas de desarrollo de B. bassiana sobre la broca del café bajo condiciones de laboratorio (25 ±1°C)
En el tratamiento con el aislamiento Bb-9205 DS, las brocas murieron en 106,1 horas, en promedio, mientras que para la cepa Bb-9205 BFC la duración fue de 66,72 horas, en promedio, después de la inoculación. La etapa de muerte a inicio de micelio en las brocas tratadas con el aislamiento Bb-9205 DS duró 20,16 horas y para el Bb-9205 BFC fue de 20,88 ± 6,72 horas, en promedio.
En relación con la etapa de muerte a cubrimiento de micelio, la duración fue de 31,92 ±14,40 horas para el aislamiento Bb-9205 DS y de 44,4 ±6,96 horas, en promedio, después de la inoculación para el aislamiento Bb-9205 BFC. La duración de la etapa muerte a conidiogénesis con el aislamiento Bb-9205 DS fue de 56,40 horas y para el aislamiento Bb-9205 BFC fue de 66,88 horas, en promedio, después de la inoculación.
Con respecto a la etapa de muerte a liberación de conidias, el tiempo de duración fue de 83,28 horas con el aislamiento Bb-9205 DS y de 116,0 horas, en promedio, después de la inoculación para el aislamiento Bb-9205 BFC.
El tiempo trancurrido desde la inoculación a la liberación de conidias para el tratamiento con el aislamiento Bb-9205 DS fue de 198 horas, en promedio, después de la inoculación y de 182 horas para el aislamiento Bb-9205 BFC. La tendencia en la duración del ciclo de desarrollo del hongo sobre la broca del café fue menor con el aislamiento Bb-9205 BFC. Estos resultados indican la conveniencia de reactivar el hongo mediante pase por brocas, lo cual se traduce en la obtención más rápida y abundante de inóculo en el ambiente.
Es importante destacar que las manifestaciones del hongo generalmente se inician en las partes intersegmentales de la broca. Posteriormente, todo el cuerpo de la broca es cubierto por el hongo, incluyendo las áreas quintinizadas. La conidiogénesis es altamente dependiente de las condiciones de humedad y se caracteriza por la presencia de conidióforos, de los cuales emergen conidias de aspecto blanco cristalino (Fig. 3), finalmente se presenta la liberación de conidias de aspecto pulverulento, caracterizada por el desprendimiento de estas.
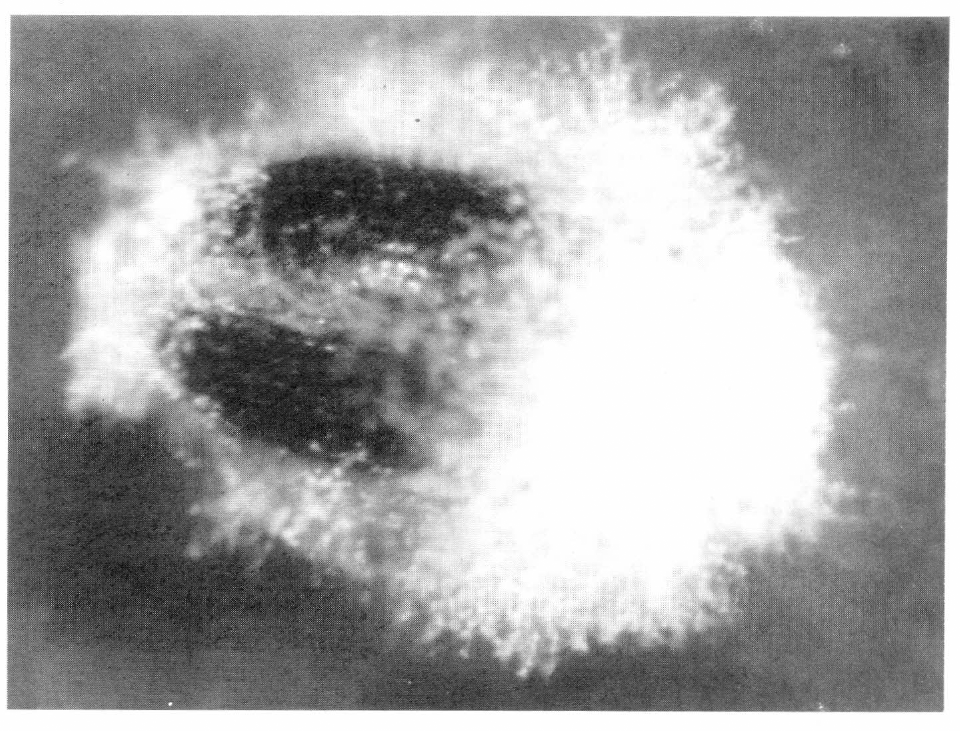
Formación de conidióforos en la etapa de conidiogénesis del hongo Beauveria bassiana sobre la broca del café
La individualización de las brocas permitió realizar evaluaciones de la patogenicidad ocasionada por la acción directa del hongo sobre cada una de ellas y no como consecuencia de la oportunidad de enfermarse por la presencia permanente de inóculo, lo cual ocurre cuando se evalúa la patogenicidad sobre poblaciones en la que una broca enferma puede estar afectando la población y servir de vehículo para la infección del grupo como lo registrado por Lazo (1990), Méndez (1990) y García (1991). Otro aspecto de este bioensayo fue que no se utilizó como sustrato para la broca un medio nutritivo con agar como el utilizado por Jiménez (1992) y así no hay incidencia de contagio causado por otros individuos que al enfermarse producen infección en el resto de la poblacion cuando se realiza el bioensayo con grupos.
Conclusiones
La metodología de bioensayo utilizada en esta investigación es confiable, de fácil manejo y reproducible para realizar los estudios de patogenicidad del hongo B. bassiana sobre la broca del café, Hypothenemus hampei.
Parámetros como el porcentaje de mortalidad, la mortalidad diaria, el tiempo letal medio (TL50) y las etapas de desarrollo del hongo deben tenerse en cuenta para determinar el potencial patogénico de los diferentes aislamientos del hongo B. bassiana sobre la broca del café.
El método de bioensayo permite evaluar la calidad de cualquier hongo entomopatógeno producido en medios diferentes, con diferentes tiempos de almacenamiento, diferentes formulaciones y concentraciones, y seleccionar las mejores cepas para el control biológico de la broca del café.
La reactivación del hongo, mediante el pase por la broca, incrementó la patogenicidad de la cepa Bb-9205 en un 11,12% y disminuyó el tiempo medio de mortalidad en 37,68 horas. Estos resultados sugieren la conveniencia de reactivar las cepas de B. bassiana mediante el pase por el huésped susceptible.