
Abstract
During 2004 and 2005, native strains of granulovirus obtained from Tecia solanivora larvae found in Cundinamarca, Nariño and Norte de Santander were isolated, identified and characterized. The effects of these infectious agents were studied on development of the potato moths T. solanivora and Phthorimaea operculella, evaluating variables such as width of the cephalic capsule, signs of viral infection and mortality. The effect of formulation was also determined on the efficiency of the different isolates formulated as a powder for potato seed protection. Additionally, the viral productivity of each isolate was studied using spectrophotometry. The highest viral production yields were obtained in P. operculella larvae, which were significantly higher than those obtained in T. solanivora larvae for all native isolates. The granulovirus infection in T. solanivora and P. operculella larvae caused a greater duration and overlap of the larval instars. The process of formulation did not significantly affect the efficacy of the granulovirus. Through a technique of weighted indicators, the isolate from Norte de Santander was selected as most promising due to its high efficiency and productivity.
Introducción
El manejo tanto en cosecha como en poscosecha de las dos principales polillas de la papa Tecia solanivora (Povolny, 1973) y Phthorimaea operculella (Zeller, 1873) (Lepidoptera: Gelechiidae), limitan la producción por los altos costos y contaminación ambiental que se genera con el uso de agroquímicos. Este problema ha llevado a la búsqueda de métodos de control menos riesgosos, más amigables con el ambiente y más sostenibles, como es el uso de productos biológicos a base de virus de la granulosis que afectan de forma natural las larvas de estas dos especies de insectos (López-Ávila 1996).
En diferentes partes del mundo se han encontrado larvas de P. operculella muertas a causa de una enfermedad natural producida por un virus del tipo granulosis (VG) que pertenece a la familia Baculoviridae (CIP 1996 citado en Zeddam et al. 1999). A partir de larvas de P. operculella infectadas con un granulovirus encontradas en Perú, el Centro Internacional de la Papa (CIP), desarrolló un bioplaguicida en polvo para la protección de semilla de papa en almacenamiento, el cual ha sido usado en diferentes países para el control de esta plaga (CIP 1996 citado en Zeddam et al. 1999). Para T. solanivora, hasta el año 2004 sólo se había reportado un aislamiento de virus de la granulosis encontrado infectando naturalmente larvas en condiciones de campo en Mérida, Venezuela (Niño de Gualdrón 1996 citado por Niño y Notz 2000). El Laboratorio de Control Biológico de la Corporación Colombiana de Investigación Agropecuaria-Corpoica, durante los años 2004 y 2005 realizó la búsqueda de aislamientos nativos de virus de la granulosis afectando de forma natural larvas de T. solanivora. A partir de este trabajo se aislaron, identificaron y caracterizaron cinco muestras de los departamentos de Cundinamarca, Nariño y Norte de Santander (Villamizar et al. 2006). Estos aislamientos nativos, se identificaron como virus de la granulosis mediante técnicas de microscopía, moleculares e inmunológicas, y se encontraron diferencias moleculares entre estos virus y el virus aislado de P. operculella proveniente del Perú correspondientes a deleciones en parte del gen 90-91 (Léry et al. 2008).
Con miras a desarrollar un nuevo bioplaguicida a base de uno de los aislamientos nativos para el control de las polillas de la papa bajo condiciones de almacenamiento y campo, surge la necesidad de seleccionar el aislamiento más virulento y el más promisorio desde el punto de vista tecnológico, donde las características más importantes a tener en cuenta son la obtención de altos rendimientos en la producción de partículas virales y en segundo lugar, que el aislamiento soporte el proceso de formulación sin perder patogenicidad. Por tal razón los objetivos del presente trabajo fueron estudiar la sintomatología de la infección causada por los aislamientos nativos de granulovirus en las polillas de la papa T. solanivora y P. operculella, determinar el efecto de la formulación sobre la actividad biocontroladora de los granulovirus nativos para las dos especies de polillas de la papa, comparar el rendimiento de producción de partículas virales de los aislamientos nativos de granulovirus en las polillas de la papa y seleccionar el aislamiento que presente las mejores características biológicas y tecnológicas.
Materiales y Métodos
Este trabajo se realizó en las instalaciones del Laboratorio de Control Biológico del Centro de Biotecnología y Bioindustria de la Corporación Colombiana de Investigación Agropecuaria-Corpoica, (Mosquera-Cundinamarca). Se utilizaron los aislamientos nativos de granulovirus VG001, VG003 y VG005 obtenidos a partir de larvas de T. solanivora recolectadas de papas almacenadas, que presentaban infestación natural, en municipios productores de los departamentos de Nariño, Cundinamarca y Norte de Santander, respectivamente (Villamizar et al. 2008); y una cepa de granulovirus de P. operculella proveniente del Perú y multiplicada por más de 20 generaciones en T. solanivora, utilizada como cepa de referencia. Todos los aislamientos se encuentran crioconservados a -70°C en el Banco de Germoplasma de Microorganismos con Interés en Control Biológico de Corpoica.
Propagación y purificación viral
Para cada uno de los aislamientos se tomaron posturas de T. solanivora provenientes de una cría ubicada en el Laboratorio de Entomología de Corpoica, luego se inocularon con una suspensión viral que se preparó a partir de una larva de T. solanivora infectada con el aislamiento y macerada en 1 mL de suero fisiológico, a esta suspensión no se le determinó la concentración. Después de la infección los huevos se incubaron a 23°C. Tras la emergencia de las larvas, éstas se ubicaron sobre tubérculos de papa variedad parda pastusa y se incubaron a 23°C durante 20 días; al final de este tiempo se realizó un análisis destructivo de cada uno de los tubérculos y se recuperaron todas las larvas. Con las larvas se llevó a cabo la purificación viral de forma independiente para cada aislamiento siguiendo la metodología estandarizada por Gómez (2005).
Determinación del rendimiento de producción de partículas virales
Se realizó una propagación viral de cada aislamiento para las dos especies de polilla de la papa (T. solanivora y P. operculella) como se describió previamente y se evaluó la producción de partículas virales expresada como cuerpos de inclusión por miligramo de tejido larval (CI/mg) siguiendo la metodología descrita por Gómez (2005). Las infecciones para cada cepa se realizaron al mismo tiempo, con la misma población larval. Se seleccionaron 15 larvas que presentaron síntomas típicos de infección por granulovirus para cada aislamiento en cada especie de polilla, se maceraron y filtraron individualmente. La suspensión se dispuso en microplacas de 96 pozos y se leyó la absorbancia a 450 nm en un lector de Elisa (Asys Hitech. Expert 96). Los resultados se extrapolaron en la curva de calibración estandarizada por Gómez (2005). El diseño experimental fue completamente al azar con 15 repeticiones por tratamiento y dos repeticiones en el tiempo. Los resultados de concentración se transformaron calculando el logaritmo decimal para cumplir con los principios de homogeneidad de varianza y normalidad de datos, y de esta forma utilizar un análisis paramétrico. El análisis de varianza y la prueba de comparación de medias de Tukey (a = 0.05) para determinar el aislamiento más productivo en cada especie de polilla se realizaron con el programa SAS 9.1; y mediante el programa Statistix 8.1.1.0 se realizó un análisis de varianza y una prueba de comparación de diferencias mínimas significativas (DMS) (α = 0.05), para determinar diferencias en la productividad de partículas virales de cada aislamiento en las dos especies.
Desarrollo de las larvas sanas de T. solanivora y P. operculella
Inicialmente se estableció el desarrollo de las larvas sanas de T. solanivora y P. operculella basados en la ley de Dyar. La variable evaluada fue el ancho de la cápsula cefálica (ACC), empezando por el tiempo cero y cada dos días para P. operculella y cada cuatro días para T. solanivora durante 32 días. Cada unidad experimental correspondió a una cubeta plástica de 16 onzas con tapa, en la cual se ubicó un tubérculo de papa criolla con 10 larvas neonatas de P. operculella, o un tubérculo de papa pastusa con 15 larvas neonatas de T. solanivora, según las preferencias alimenticias previamente establecidas para cada especie de insecto en condiciones de cría. Las unidades experimentales se incubaron a 23°C y a una humedad relativa (HR) del 65%.
En cada tiempo de medición y para cada polilla se realizaron muestreos destructivos de los tubérculos en busca de las larvas, se midió el ACC de cada una de las larvas con un micrómetro adaptado en el ocular de un microscopio óptico con un aumento de 4 X. Posteriormente se estableció el rango del ACC y se realizó una distribución de frecuencias con el fin de determinar los diferentes ínstares larvales y compararlos con el de las larvas sanas.
El montaje de los ensayos se realizó de la misma forma para todos los aislamientos de granulovirus en las dos especies de polilla. Huevos de las polillas depositados en papel absorbente (400-500 huevos/papel) se inocularon con 3mL de una suspensión viral a una concentración ajustada a 5x106 CI/mL para cada aislamiento. En el interior de un recipiente plástico de 16 onzas se ubicó un tubérculo de papa con 15 larvas neonatas obtenidas a partir de las posturas inoculadas. Se realizó un muestreo destructivo cada cuatro días para T. solanivora y cada dos días para P. operculella, tomando al azar tres unidades experimentales (tubérculos) durante 32 días. La variable evaluada fue el ACC como se describió anteriormente, se determinó la distribución de frecuencias para cada aislamiento viral y se comparó con la de las larvas sanas. El ensayo contó con una repetición en el tiempo.
Eficacia de los aislamientos de Granulovirus
Todos los aislamientos de granulovirus se formularon siguiendo la metodología utilizada por Corpoica para la producción del bioplaguicida registrado comercialmente utilizando la cepa del Perú. La concentración final del producto para todos los aislamientos virales fue de 1 x 106 CI/g. Los aislamientos sin formular consistieron en suspensiones virales ajustadas a la concentración de 1 x 106 CI/mL.
Para cada tratamiento, se pesaron tres tubérculos de papa y se ubicaron en una bolsa plástica que contenía el producto formulado según la dosis comercial (5g bioplaguicida/Kg papa. Producto a base de baculovirus registro ICA # 5416). Las bolsas se cerraron y se agitaron hasta que los tubérculos quedaron cubiertos con el producto. Para los tratamientos con los virus sin formular, los tubérculos se inocularon con una brocha de ½ pulgada aplicando 2 mL de la suspensión viral en la superficie. El tratamiento control consistió en tres tubérculos sin aplicación viral, sobre los que se colocaron 15 larvas recién emergidas y se incubaron a 23°C durante 20 días, momento en el cual se realizó un muestreo destructivo de los tubérculos en busca de las larvas. Se contó el número de larvas muertas, larvas vivas sin signos de infección, larvas vivas con signos de infección y el número de pupas obtenidas. Los resultados de mortalidad en cada tratamiento, se corrigieron determinando el porcentaje de eficacia mediante la fórmula de Schneider-Orelli (Zar 1999): Eficacia = (b – K/100 – K) x 100. En donde b es el porcentaje de mortalidad en el tratamiento y K es el porcentaje de mortalidad del testigo. El diseño experimental fue completamente al azar con tres repeticiones por tratamiento. El porcentaje de eficacia de los productos, se analizó con el programa Statistix 8.1.1.0 mediante un análisis de varianza y una prueba de comparación de diferencias mínimas significativas (DMS) (α = 0.05).
Selección del aislamiento de granulovirus
Con el fin de seleccionar el mejor aislamiento de granulovirus, se analizaron los valores de eficacia y productividad por medio de la técnica de ponderación de indicadores de Dean y Nishry (1965). Para esto se definió la importancia de dichas variables según el impacto de cada una en el proceso de producción de un controlador biológico. Se determinó el coeficiente de importancia (CIV) de cada variable, según la importancia relativa de cada una y con esto se calculó el coeficiente de selección de cada aislamiento viral (CSAV) para determinar la importancia de cada una de las variables en las cepas evaluadas. El CSAV se multiplicó por el CIV y se obtuvo el índice de selección final (ISF) con el que se construyó una matriz de decisión.
Resultados y Discusión
Propagación y purificación viral
En la propagación viral de todos los aislamientos se observaron síntomas típicos de infección por granulovirus en la mayoría de las larvas. Después de las centrifugaciones diferenciales y de correr todas las muestras en el colchón de glicerol, se evidenció la formación de una banda blanca entre las concentraciones de 60% y 80%, correspondientes a los cuerpos de inclusión purificados de cada aislamiento de granulovirus como se ha reportado por otros autores como Taha et al. (2000) y Sporleder et al. (2005), quienes confirmaron por microscopía electrónica que esta banda corresponde a partículas virales. Este método de purificación viral, permitió obtener de forma fácil y rápida las partículas virales.
Rendimiento de producción
Las 15 larvas mostraron claros signos de la infección por granulovirus, correspondiente a una coloración blanca y un retraso notorio en el crecimiento. Los datos de absorbancia a 450nm en todos los casos fueron mayores a 0,04nm. Los rendimientos de producción de partículas virales para cada uno de los aislamientos estuvieron en un rango de 109 a 1010 cuerpos de inclusión por miligramo de tejido larval, tanto para P. operculella como para T. solanivora. Para P. operculella se encontró que la producción de cuerpos de inclusión por miligramo de tejido larval fue significativamente mayor (P < 0,0004) con el aislamiento VG005, en comparación con los demás aislamientos; mientras que los aislamientos VG001, Perú y VG003 no presentaron diferencias significativas entre sí (Tabla 1).
Productividad y eficacia de los diferentes aislamientos de granulovirus en T. solanivora y P. operculella. Tratamientos con la misma letra no presentan diferencias significativas según prueba DMS (α = 0.05).
Teniendo en cuenta que el granulovirus VG005 se aisló a partir de larvas de T. solanivora, las diferencias de productividad viral encontradas podrían sugerir que este aislamiento está menos adaptado a P. operculella y que por lo tanto, necesita producir una mayor progenie viral para garantizar su mantenimiento y supervivencia en este hospedero (Moscardi 1999). En T. solanivora la producción de cuerpos de inclusión por miligramo de tejido larval fue mayor en el aislamiento de Perú, aunque no fue significativamente diferente a los obtenidos con los aislamientos VG005 y VG001, pero que sí fue estadísticamente superior (P < 0,0004) al rendimiento obtenido con el aislamiento VG003 (Tabla 1).
El granulovirus proveniente de Perú fue aislado a partir de larvas de P. operculella; sin embargo, produjo el mayor rendimiento en larvas de T. solanivora en comparación con los aislamientos virales cuyo hospedero original fue T. solanivora. Este resultado podría estar relacionado con diversos mecanismos de adaptación que los virus utilizan para mantenerse y reproducirse dentro de un nuevo hospedero (Parnell et al. 2002; Herniou et al. 2004). Sin embargo, para T. solanivora no se encontraron diferencias significativas con los aislamientos VG001 y VG005 aisladas de esta especie, lo que sugiere un avanzado proceso de adaptación del virus del Perú al hospedero secundario, debido posiblemente a la constante propagación que se ha realizado de éste en larvas de T. solanivora (más de 20 pases) (Moscardi 1999).
Se obtuvo una producción de CI por miligramo de tejido significativamente mayor (P = 0.0004) en larvas de P. operculella que en larvas de T. solanivora para los aislamientos VG003, VG001 y VG005 (Tabla 1). Para el caso del aislamiento del Perú, la producción de partículas virales no presentó diferencias significativas entre las dos especies de polillas. Para seleccionar el aislamiento más productivo de granulovirus tanto en larvas de P. operculella como de T. solanivora, es importante tener en cuenta la capacidad replicativa de los virus en estos hospederos, su historia evolutiva, su adaptación y la facilidad, costos y eficiencia de las crías de los insectos. Se podría sugerir la utilización de larvas de P. operculella para la producción masiva de los granulovirus, pues se obtuvieron los mayores rendimientos de partículas virales; sin embargo, estos rendimientos podrían disminuir tras pases consecutivos de los virus en este hospedero debido a los procesos de adaptación mencionados anteriormente; también se debe tener en cuenta el momento de infección, ya que existe un proceso de adaptación del virus que puede llevar a la disminución de la virulencia dependiendo si la transición del virus entre hospederos se hace temprana o tardíamente (Cooper et al. 2002). Teniendo en cuenta este fenómeno se recomienda mantener un abundante inóculo viral original crioconservado y adecuadamente alicuotado, de manera que después de algunos pases en el insecto se recurra a la cepa original conservada, no adaptada al hospedero y realizar el proceso de infección en una etapa temprana para obtener una rápida replicación del virus y aumentar su virulencia.
Desarrollo de los signos de infección
El crecimiento de las larvas de T. solanivora y P. operculella cumplió con la ley de Dyar, debido a que el crecimiento de las partes esclerotizadas como la cápsula cefálica de ínstares sucesivos siguió una progresión geométrica regular (Dyar 1935 citado por Gamboa y Notz 1990). Con el análisis de distribución de frecuencias del ancho de la cápsula cefálica (intervalo de 0,1 para T. solanivora y 0,04 para P. operculella), se establecieron cuatro grupos sin solapamiento los que sugieren la formación de cuatro ínstares larvales tanto para T. solanivora como para P. operculella (Fig. 1).
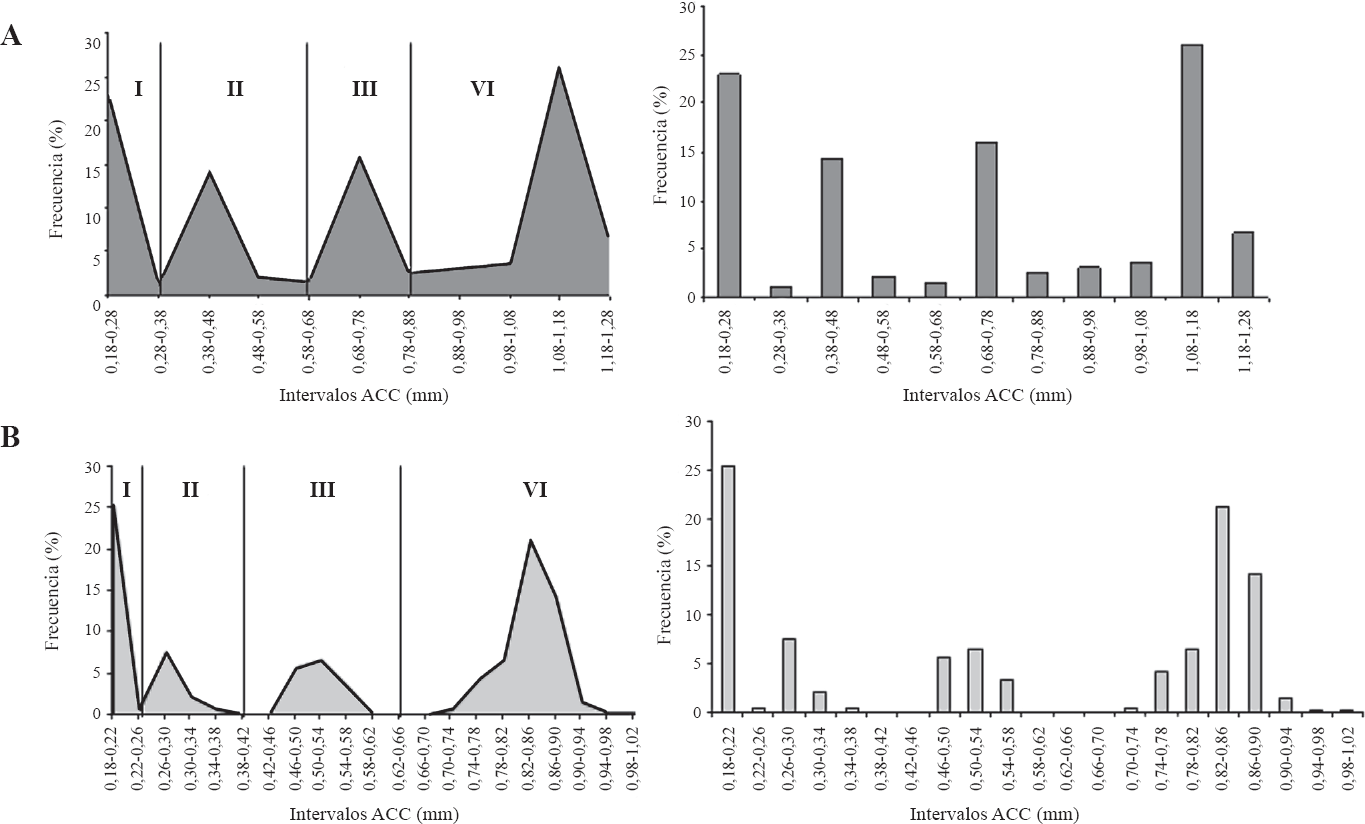
Distribución de frecuencias del ancho de la cápsula cefálica.
Los cuatro ínstares larvales de T. solanivora y su duración fueron registrados también por y Niño y Notz (2000) y Villamizar et al. (2006). Para el caso de P. operculella este mismo comportamiento fue reportado por Llanderal (1993) y Gamboa y Notz (1990). Para T. solanivora el AAC va de 0,18 a 0,38 mm para el primer ínstar, de 0,39 a 0,68 mm para el segundo ínstar, de 0,69 a 0,88 mm para el tercer ínstar y de 0,89 a 1,28 mm para el cuarto instar (Fig. 1A). Para P. operculella el ACC va de 0,18 a 0,26 mm para el primer ínstar, de 0,27 a 0,42 mm para el segundo ínstar, de 0,43 a 0,66 mm para el tercer ínstar y de 0,67 a 1,02 mm para el cuarto ínstar (Fig. 1B). Estas escalas han sido reportadas por otros autores con algunas diferencias que se pueden presentar por los diferentes intervalos de medición utilizados influenciados por cambios ambientales, condiciones nutricionales variables debido a diferencias en el sustrato alimenticio, cambio en la proporción de sexos y variaciones en el estado fisiológico de las larvas (Llanderal 1993).
En cuanto al comportamiento de los estados de desarrollo larval de T. solanivora y P. operculella infectadas con los diferentes aislamientos de granulovirus se observó en todos los casos una mayor duración de los diferentes ínstares, lo que generó un traslape de los mismos (Figs. 2 y 3). Este efecto fue reportado por Reed (1971 citado por Alcázar et al. 1992), Alcázar et al. (1992), Niño y Notz (2000) y Villamizar et al. (2006).
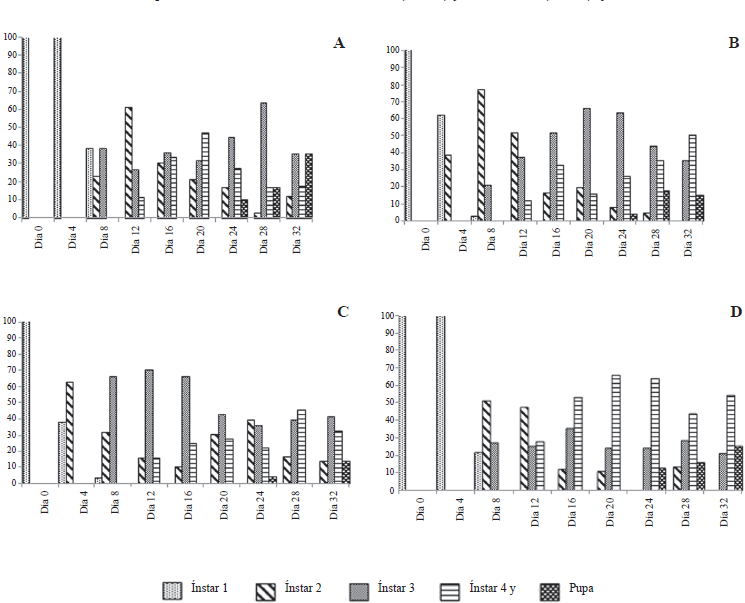
Comportamiento de los ínstares larvales (% frecuencia) de T. solanivora infectada con granulovirus.
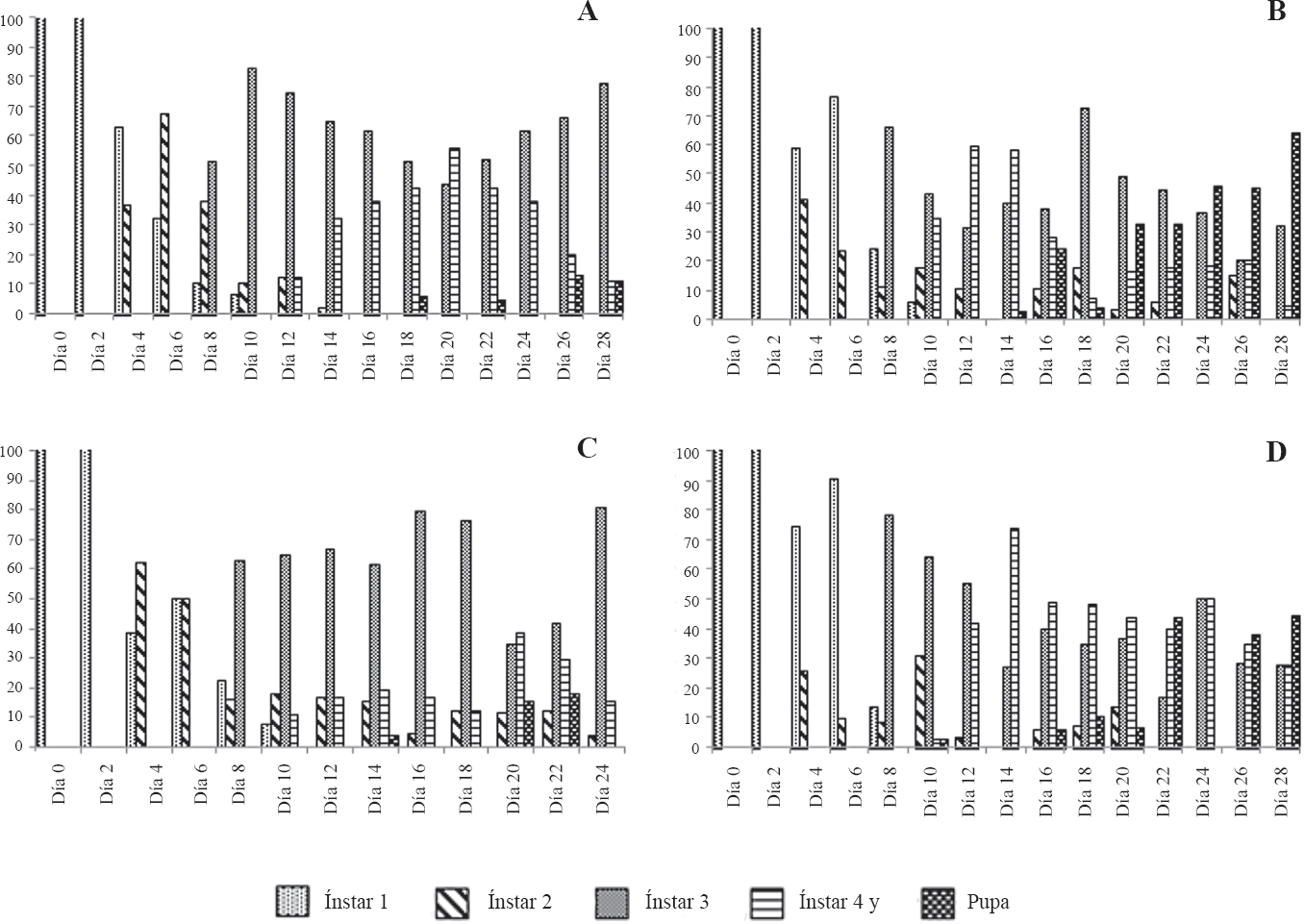
Comportamiento de los ínstares larvales (% frecuencia) de P. operculella infectada con granulovirus.
El retraso en el desarrollo de las larvas infectadas con los diferentes aislamientos pudo deberse al efecto negativo que tiene el virus en los hábitos alimenticios y en la expresión hormonal de las larvas durante el cambio sucesivo de ínstares (Burand y Park 1992; Nakai et al. 2004; Subramanian et al. 2005). Según Moscardi (1999), una característica de la infección ocasionada por baculovirus es el retraso del desarrollo y la reducción de la capacidad alimenticia de las larvas.
Por otro lado, en los primeros días de desarrollo no se evidenciaron signos de infección en las larvas de T. solanivora y P. operculella, observándose larvas de coloración parda translucida. Luego de algunos días aparecieron aglomeraciones de estructuras blanquecinas observadas a través del abdomen de las larvas, principalmente en la parte posterior. Estas aparecieron por todo el cuerpo a medida que transcurrió la infección, hasta que las larvas adquirieron un color blanco lechoso en todo su cuerpo. Estos signos también han sido reportados por Alcázar et al. (1992), Niño y Notz (2000) y Villamizar et al. (2006).
Para los últimos días de medición (días 28 y 32) se encontró que las larvas infectadas con el granulovirus VG005 y Perú adquirieron una tonalidad rosada en la parte dorsal del cuerpo, pero sin perder el color blanco-lechoso adquirido por la infección de granulovirus. Según Niño (2004) y Villamizar et al. (2006), esta pigmentación rosada que aparece en la parte dorsal de las larvas en los últimos días de desarrollo, denotaría el inicio del cambio del estado de larva al estado de pupa, el cual se presentaría normalmente para larvas sanas. La ausencia de la coloración rosada en algunas larvas, puede deberse al efecto de la infección viral (Alcázar et al. 1992; Caballero et al. 2001).
Eficacia de los aislamientos de Granulovirus
Para T. solanivora se encontró que los tratamientos que produjeron un mayor porcentaje de eficacia fueron los aislamientos VG003 formulado (81,8%) y sin formular (69,6%), VG005 formulado (75,7%) y sin formular (72,7%), y el aislamiento del Perú sin formular (75,7%), los cuales no fueron significativamente diferentes (DMS a = 0.05); pero sí presentaron un porcentaje de eficacia estadísticamente superior (P = 0.1658) con respecto al aislamiento VG001 formulado (36,37%) (Fig. 4A). En P. operculella se encontró que los tratamientos que produjeron un mayor porcentaje de eficacia fueron los aislamientos VG001 formulado (94,7%) y sin formular (100%), VG003 formulado y sin formular (97,3%), Perú formulado (94,7%) y sin formular (96,7%) y VG005 sin formular (97,4%); los cuales no presentaron diferencias estadísticas entre sí, pero si presentaron diferencias (P = 0.1120) con respecto al aislamiento VG005 formulado (84,2%) (Fig. 4B).
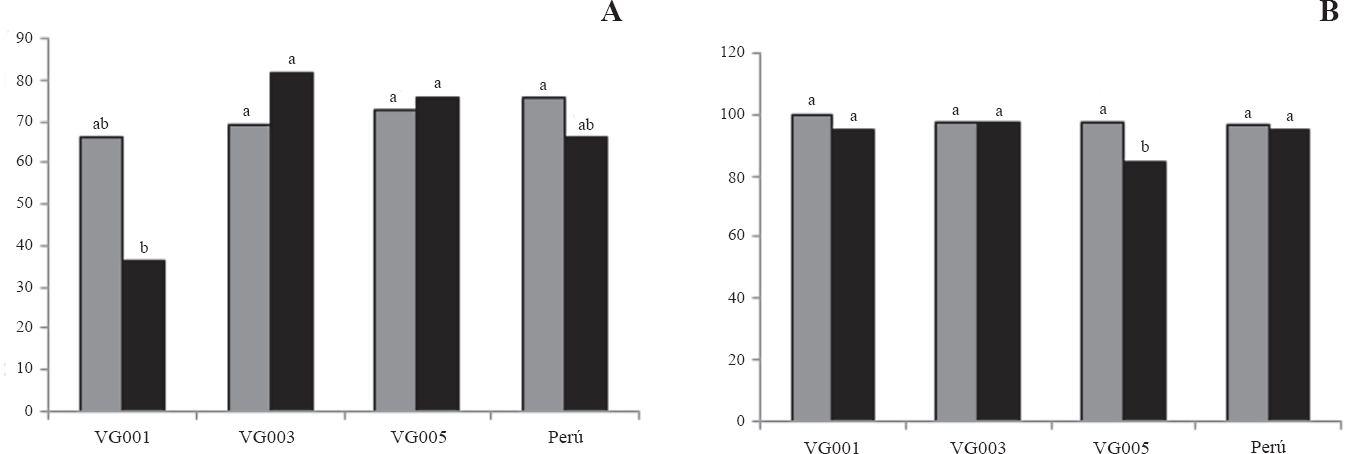
Eficacia de aislamientos de granulovirus formulados y sin formular en larvas de las polillas de la papa.
En términos generales no hay diferencias estadísticas entre la actividad biocontroladora de cada aislamiento formulado y sin formular, resultados que sugieren que el proceso de formulación no afectó la eficacia de los granulovirus. Resultados similares fueron reportados por Niño y Notz (2000), quienes no encontraron diferencias en la mortalidad de las larvas de T. solanivora ocasionada por un granulovirus aplicado como suspensión acuosa de partículas virales y el mismo granulovirus formulado. Cossentine y Jensen (2004) tampoco encontraron un incremento significativo de la eficacia de un granulovirus de Cydia pomonella (Linnaeus, 1758) (Lepidoptera: Tortricidae) al incorporar algunos aceites en su formulación.
Selección del aislamiento
Con miras a seleccionar el aislamiento más promisorio para el desarrollo de un bioplaguicida para el control de T. solanivora y P. operculella, se seleccionaron las variables de eficacia y productividad, las cuales fueron analizadas mediante la técnica de ponderación de indicadores de Dean y Nishry (1965). El coeficiente de importancia para cada factor fue mayor para la variable de eficacia con un valor de 0.5875 en comparación al obtenido para productividad con un valor de 0.4125. Posteriormente se llevó a cabo la ponderación de los factores de selección para cada uno de los aislamientos virales, con base en los datos de los ensayos de productividad viral y eficacia (Tabla 1). Los valores correspondientes a los coeficientes de selección de cada aislamiento viral se muestran en la Tabla 2.
Coeficientes de selección de los aislamientos de granulovirus (CSAV).
A partir de la matriz de decisión se obtuvieron los mayores índices de selección final tanto para T. solanivora como para P. operculella con el aislamiento nativo VG005 proveniente de Norte de Santander con valores de 0.111 y 0.189 respectivamente (Tabla 3). Teniendo en cuenta este resultado se recomienda el uso de este aislamiento para la producción de bioplaguicidas para el control de las dos polillas de la papa tanto en campo, como en almacenamiento.
Matriz de decisión para la selección del mejor aislamiento viral para el desarrollo de un bioplaguicida para el control de T. solanivora y P. operculella.