
Abstract
Thrips palmi is an exotic pest registered for Colombia since 1997. This research was made attempting to acquire a best knowledge about insect 's biology and ecology, specially his lifecycle. It was also wanted to account the consumption capability of Chrysoperla carnea (Neuroptera: Chrysopidae) on T. palmi. The environmental conditions were a medium temperature of 26°C (d.s. 2,19) and a medium relative humidity of 53, 06% (d.s.5, 54). The experimental unit was an uncolored photographic plastic can cutted by the middle. Adults of T. palmi were collected at the field and his progeny was used for the evaluation of inmature's lifetime and adult's longevity. The predatory capability of C.carnea was evaluated at 24 hours of predator-prey interaction at levels of 10, 20 and 30 preys and 0,1,2,3 predators. The medium times in days for inmatures feed on beans (Phaseolus vulgaris) was 2,14 for the first ninphal instar (n1); 1,56 for the second (n2); 1,05 for prepupae (pp) and 2,2 for pupae (p). The livehope (ex) for n1 was 1,99; for n2: 1,83; for pp: 1,71, for p: 0,91. The sex ratio was 1:3,5 (male/female). Chrysoperla carnea had a medium consumption of 14,4 trips nymphs in 24 hours. The variance analysis reveals that the statistically significative effect obtained (F: 23,71; p: 0,0001) was related to different predator densities and not to the different prey densities. The plots that included predator had significative differences with plots with no predator. At present prey level, there were no significative differences within two or three predators. Despite the crysopids may have a higher consumption of thrips at high-density level (3 predators), the individual effectiveness is reduced by interference and cannibalism (in this case related with laboratory conditions).
Keywords
Introducción
Thrips palmi Karny es una plaga polífaga exótica registrada en Colombia desde 1997 (Vergara 1999). Su rango de dispersión abarca hoy en día un gran número de agroecosistemas y se estima que su dispersión abarque en un futuro no muy lejano la mayoría de las áreas del planeta (Parrella y Mound 1998). Las características biológicas y ecológicas de este organismo, lo convierten en un elemento clave en la producción agrícola ya que sus poblaciones se desarrollan epidémicamente y posee una capacidad de adaptación genética elevada (Vergara 1998).
En todos los lugares del mundo donde este insecto ha sido registrado, se han producido cuantiosas pérdidas relacionadas con su presencia (Madrigal 1998; Pest Cabweb 1997; Cooper 1991; Agudelo y Vergara 1998; Rendón et al. 1999). Estas pérdidas se derivan tanto de los daños directos, al reducir la calidad o cantidad de la producción, como de los daños indirectos, relacionados con la transmisión de virus como el Tobaco Spoted Wiltlike Virus, TSWV y el Peanut Bud Necrosis Virus, PBNV (Yeh et al. 1992; Buiel et al. 1995).
Debido a la importancia de T. palmi en los ecosistemas agrícolas, esta investigación se adelantó para conocer la duración del ciclo de vida del insecto bajo condiciones de laboratorio y evaluar la capacidad de consumo de larvas de Chrysoperla carnea (Neuroptera: Chrysopidae) sobre las ninfas del trips.
Materiales y Métodos
Ubicación
El estudio se llevó a cabo en el Centro de Sanidad Agropecuaria adscrito a la Secretaría de Agricultura de Antioquia en la ciudad de Medellín, con una temperatura media de 26° C (d.s. 2,19) y una humedad relativa media de 53,06% (d.s. 5,54).
Unidad de experimentación
La unidad de experimentación empleada fue la adoptada por Durán (1999) y Vasco (1999), la cual consiste en un tarro para rollo fotográfico transparente con un diámetro aproximado de 2,5 cm, cortado a la mitad. Dentro de este se colocó papel absorbente para el manejo de la humedad y un disco de hoja de fríjol como sustrato alimenticio.
Material vegetal y Thrips
Se estableció una siembra escalonada de fríjol variedad ICA Viboral de la que se emplearon plantas que tuvieran como máximo tres semanas de edad. Para contar con un material vegetal uniforme libre del efecto de otros insectos, una vez germinadas las plántulas, se colocaron en cajones de madera de 33x33x100 cm forrados con malla fina.
Se colectaron del campo adultos de T. palmi (aproximadamente 200) para el establecimiento de una colonia de laboratorio. Se efectuaron periódicamente salidas al campo para la recolección de nuevos adultos en cultivos de fríjol. Para obtener cohortes de la misma edad, se tomaron plantas de fríjol de aproximadamente 10 días, se infestaron manualmente con trips adultos (variable entre 20 y 160 según la disponibilidad) y se dejaron sobre éstas por 48 horas. Después de este lapso de tiempo los insectos se removieron de las plantas. Dos días después de la desinfestación, se cortaron las hojas y se colocaron individualmente en cajas Petri con una mota de algodón humedecida adherida al pedúnculo. La parte superior de las cajas se cerró con plástico transparente de cocina (Vinipel®) hasta la emergencia del primer instar ninfal. Luego fueron trasladadas a la unidad de experimentación mediante un pincel fino (No. 000).
Diariamente los tarros se destaparon para airearlos, rehumedecer el papel cuando era necesario y registrar cambios. El disco de hoja se cambió cada uno o dos días, según su grado de deterioro, hasta que se observaron las prepupas. Si este estado se encontraba sobre el disco de hoja, se dejaba allí hasta la emergencia del adulto. Los adultos se mantuvieron en condiciones de aislamiento, alimentados con fríjol hasta su muerte. La relación de sexos se estimó utilizando las variaciones señaladas por Vasco (1999) y otros investigadores, y reconociendo las hembras por la presencia del ovipositor en observaciones con el microscopio (40X).
Con el total de los datos colectados, se elaboró una tabla de vida horizontal y posteriormente se construyó una curva de sobrevivencia según Rabinovich (1980) y Krebs (1985) adoptando las modificaciones de Ortega (1992).
Evaluación de la capacidad depredadora de C. carnea
Se confinaron diferentes densidades de ninfas de trips (10, 20 y 30) con varias densidades de larvas de primer y segundo ínstar de C. carnea (0, 1, 2 y 3) en la unidad de experimentación.
Análisis estadístico
Se efectuaron 3 repeticiones del ciclo de vida. En cuanto a los experimentos de depredación, cada tratamiento contó con un número variable de replicaciones, entre 9 y 11 de acuerdo con los trabajos de Laing y Osborn (1974), Kajita (1986), Nicholls (1994), Coll y Ridgway (1995) y Croft et al. (1998).
El análisis estadístico del proceso de la depredación se realizó tomando los datos a las 24 horas, como lo proponen los mismos autores. Diversas transformaciones de los datos fueron empleadas (%, arcsen %, arcsen√%) según lo recomendado por varios investigadores (Ehler y Miller 1973; Obrycki et al. 1989; Heinz et al. 1995; Kontsedalov et al. 1998; Vasco 1999). El procedimiento lineal del programa SAS® (GLM) se empleó para el análisis de varianza, así como la prueba de rangos múltiples de Duncan y la prueba de la mínima diferencia significativa (LSD) para el análisis de las medias.
Resultados y Discusión
Aspectos biológicos
La duración promedio en días de los diferentes estados del ciclo de vida de T. palmi fue: 2,14 (0,59 d.s.) para la ninfa de primer ínstar (n1). Esta es de tamaño muy reducido comparada a los adultos, los artejos antenales son globosos y poco diferenciados y sus ojos tienen una coloración roja muy fuerte. La ninfa de segundo ínstar (n2) duró 1,56 (0,63 d.s.), exhibiendo un tamaño aproximadamente el doble que el ínstar anterior y con los artejos antenales bien definidos. La duración de la prepupa (pp) fue de 1,05, y se caracterizó por la presencia de los primordios antenales y por tener las antenas perpendiculares al plano del cuerpo. La duración de la pupa (p) fue de 2,2. Esta tiene los cojines antenales mucho más desarrollados y las antenas plegadas sobre el dorso del cuerpo. Los adultos tuvieron una duración promedio de 8,21. La duración total del ciclo fue de 15,6 días, estimándose además un tiempo aproximado para el huevo entre 3 y 6. La relación macho-hembra fue de 1:3,5.
Los datos encontrados en este trabajo son similares a los de Bueno y Cardona (1999) cuyos experimentos fueron desarrollados a una temperatura media similar, a excepción del estado de prepupa en el cual, los mencionados investigadores señalan una duración de 2,6 (±1) días.
Con esta información se construyó la tabla de vida de T. palmi (Tabla 1). La tabla se inicia a partir de las ninfas de primer ínstar, debido a que la duración del estado de huevo sólo pudo ser aproximada a partir de las fechas de infestación al inicio de cada cohorte y las fechas de aparición de las ninfas de las mismas. Sin embargo, el rango calculado es similar a los arrojados por otros trabajos en el país (Bueno y Cardona 1999; Durán et al. 1999; Vasco 1999).
Tabla de vida de Thrips palmi
Se puede apreciar que la esperanza de vida (ex) de cada ínstar se va reduciendo a medida que se avanza en estado de desarrollo. Por otra parte, el estado en donde se encontró una mayor mortalidad (qx) fue la pupa, mientras que el menor porcentaje de mortalidad se obtuvo en la prepupa. Una de las causas naturales de mortalidad que con más frecuencia se detectó, tiene que ver con problemas relacionados con la formación de individuos atípicos, durante la metamorfosis. Se encontraron especímenes con las patas o las antenas pegadas, justo después del cambio de ínstar.
A partir de la tabla de vida se construyó la curva de sobrevivencia, es decir, la proporción de sobrevivientes por edad (Ix) en cada estado de desarrollo. Esta se aprecia en la Figura 1.
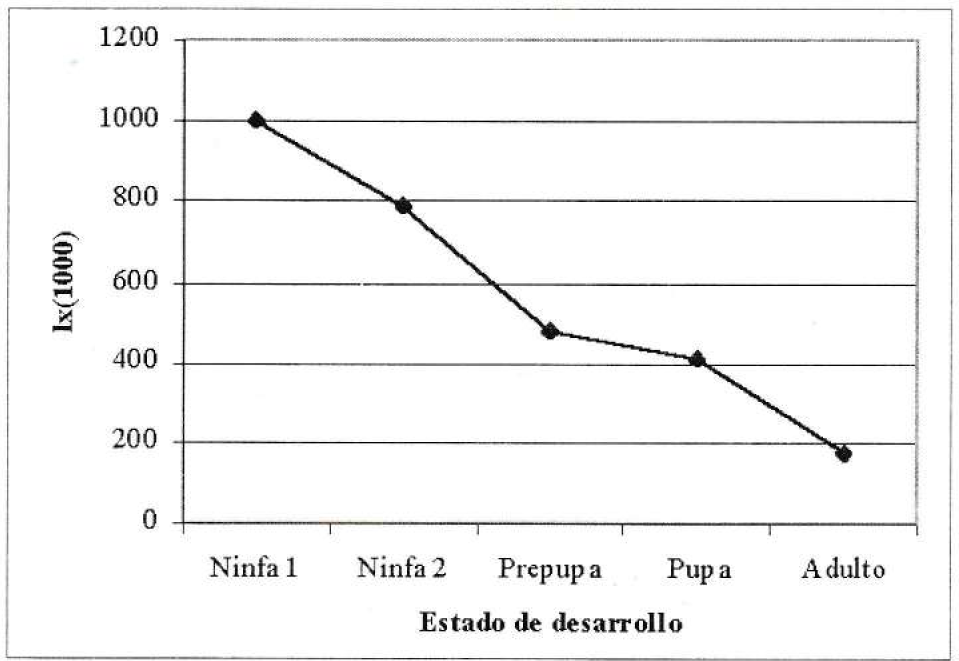
Curva de sobrevivencia de Thrips palmi.
La curva de sobrevivencia encontrada es del tipo II; es decir, la mortalidad de los diferentes estados fue aproximadamente constante en ellos a medida que se avanza en su desarrollo. Sin embargo, debe tenerse presente que dicha curva es elaborada en condiciones "ideales" para los individuos al ser un trabajo de laboratorio (Nicholls 1994).
El porcentaje de adultos resultantes con relación al número de ninfas de primer ínstar eclosionadas fue de 7,4% aproximadamente. Esto evidencia el alto potencial destructivo del T. palmi, si se tiene en cuenta que las explosiones de insectos- plaga frecuentemente se desarrollan a partir de la sobrevivencia de menos del 2% de los insectos inmaduros (Whitcomb y Hydorn 1979).
Consumo de Chrysoperla carnea
C carnea presentó un consumo promedio de 14,4 ninfas de trips en 24 horas. El análisis de varianza reveló que el efecto estadísticamente significativo encontrado (F: 23,71; p: 0,0001) se debió a las diferentes densidades del depredador (0, 1, 2, 3) (F: 81,71; p: 0,0001; variable dependiente: arcsen√%mortalidad) y no a las diferentes densidades de presa (10, 20, 30) (F: 0,03; p: 0,9735). Esto en cierta medida concuerda con lo afirmado por Holdsworth (1968), quién encontró que las poblaciones de C. rufilabris y de C. carnea están relacionadas tanto con densidades altas como con densidades bajas de su presa, lo cual podría considerarse como una característica adecuada para un insecto que es incluido en programas de manejo integrado de plagas.
Los ensayos que incluyeron depredador demostraron ser significativamente diferentes al testigo con cero depredadores. Con el nivel de presas evaluadas, no hubo diferencias significativas entre los tratamientos con dos y con tres depredadores. Aunque se observó que las crisopas pueden llegar a consumir más presas a la densidad superior (tres depredadores), la efectividad individual se vio reducida por los efectos de interferencia y canibalismo. Los datos globales de depredación pueden apreciarse en la tabla 2 y gráficamente se puede ver la relación depredador-presa en las figuras 2 y 3.
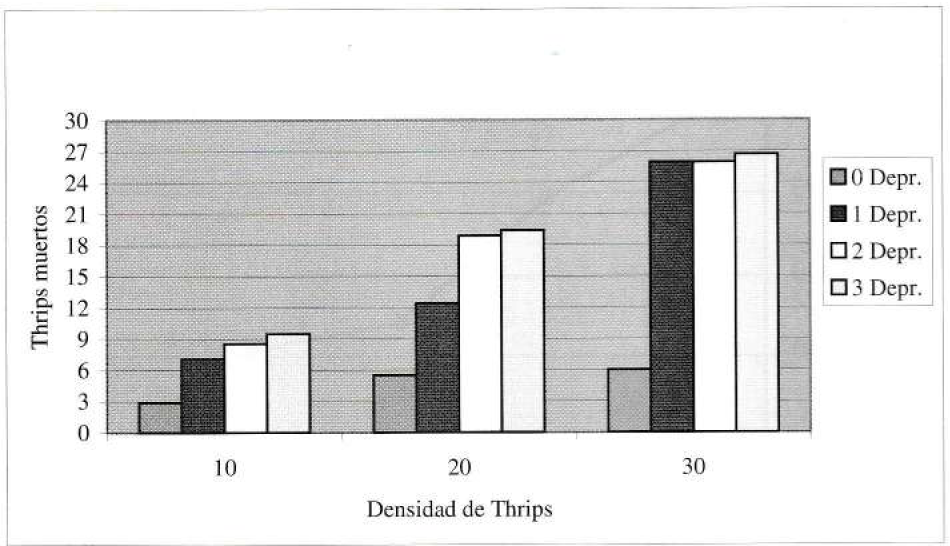
Número promedio de Thrips palmi muertos a diferentes densidades.
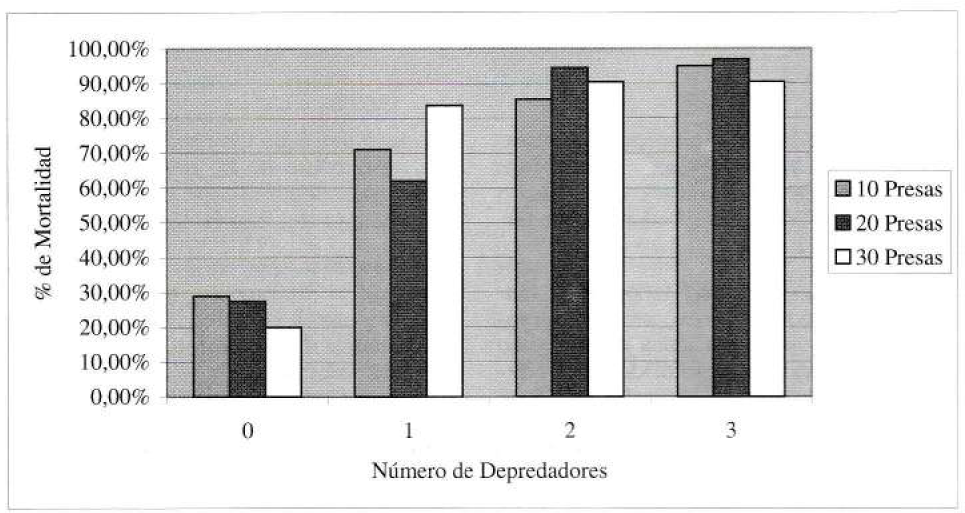
Porcentaje promedio de mortalidad de Thrips palmi a diferentes densidades del depredador.
Promedio de Thrips palmi depredados por Chrysoperla carnea
Depr: Número de depredadores; n: Repeticiones; P.P.C.: Promedio Presas Consumidas; E.E.: Error estándar.
La tasa promedio de consumo de C. carnea fue 14,4 ninfas/día, similar a lo encontrado por Callan en 1943, citado por Sabelis y Van Rijn (1997), quien utilizó como presa al trips Selenothrips rubrocintus (Thysanoptera: Thripidae) obteniendo una tasa máxima de depredación por larva/día de 14,2 trips.
C. carnea consumió alrededor de 21 ninfas a la densidad de 30 presas. Comparando esto con diferentes tasas de consumo diarias de algunos antocóridos depredadores de reconocida importancia (Seal 1997; Kajita 1986; Mituda y Callilung 1988) se reafirma el potencial que tiene C. carnea para ser incluido en los programas de manejo del T. palmi. Por otra parte, se encontró que bajo las condiciones evaluadas, el máximo de 30 presas fue insuficiente para llegar al tope de la capacidad de consumo del depredador.
Debe anotarse que en las primeras 8-10 horas de la interacción, de 63 unidades experimentales posibles, 26 presentaron canibalismo de algún tipo. Esto concuerda con lo afirmado por Tejada (1976), Bar y Gerling (1985), Dreistadt et al. (1986), Vargas (1988) y Villanueva (1985) sobre la alta incidencia de depredación intraespecífica de estos insectos. El área de la unidad de experimentación sin duda influye en la cantidad de encuentros entre depredadores y por tanto, en la probabilidad de ocurrencia del canibalismo. Tejada (1976) afirma que el número total de presas destruidas se incrementa con la adición de más depredadores, lo cual a rasgos generales concuerda con los resultados obtenidos en esta investigación.
La reducida diferencia encontrada entre las densidades de depredadores tiene relevancia en un futuro en los programas de control biológico del T. palmi. Al optar por usar una u otras densidades del depredador, deberá valorarse claramente tanto el estado fenológico del cultivo y la severidad del ataque, junto con los costos que implique utilizar a los depredadores y los beneficios económicos que se persigan al nivel de producción. En investigaciones de campo, Ridgway y Jones (1969) encontraron que el efecto (en la reducción de los huevos de las presas) de liberaciones inundativas de C. carnea para controlar el complejo Heliothis en algodón, fue similar con liberaciones de 10.000, 50.000 y 200.000 huevos/acre. De todas formas, Kawai (1995) encontró que aunque diferentes cantidades del depredador, pueden ejercer un control similar sobre su presa, el tiempo requerido para alcanzar el pico máximo de las poblaciones de depredadores está influenciado por este factor. Este aspecto es un punto clave para mantener bajo control al insecto plaga.
Conclusiones
La duración promedia de los diferentes estados inmaduros de Thrips palmi fue de 3 días para el huevo; de 2,14 días para la ninfa de primer instar; de 1,56 días para la ninfa de segundo ínstar; de 1,05 para la prepupa y de 2,2 para la pupa, con un total de 9,95 días. La longevidad promedio del adulto fue de 8,21 días.
Bajo las condiciones evaluadas, Chrysoperla carnea es un depredador efectivo de T. palmi, con un consumo promedio de 14,4 ninfas de trips por día. Al observar el aumento en la mortalidad de T. palmi con un aumento en el número de depredadores, se puede afirmar que C. carnea tiene potencial como agente regulador de este insecto-plaga.
En las condiciones evaluadas, C. carnea no presentó un marcado aumento en el consumo a diferentes densidades de la presa, siendo su efectividad similar a las densidades de trips.
Debido a la importancia económica del insecto, se deben seguir desarrollando estudios básicos relativos a su biología, así como a la de sus potenciales controladores biológicos y sobre las interacciones entre estos, para poder hacer un manejo eficaz y ambientalmente aceptable de este organismo.
Footnotes
Agradecimientos
Los autores agradecen a la Secretaría de Agricultura de Antioquia y a la Universidad Nacional de Colombia sede Medellín por permitir y apoyar esta investigación. Igualmente a Nora Cristina Mesa, Germán Cabarcas I., Alvaro Lema T., Oscar Efraín Ortega, Natalia Andrea Gallego, Gonzalo Abril y Jhon Albeiro Q. por su valioso apoyo, colaboración y comentarios.