
Abstract
The biocontrol activity of native isolates of the enthomogenous fungi Beauveria bassiana and Metarhizium anisopliae on Tecia solanivora was evaluated. A method for disinfecting the eggs and third instar larvae using sodium hypochloride [0.5%] and various rinses with sterile distilled water was standardized. First instar larvae and eggs were inoculated by immersion in a conidial suspension of 10⁸ propagules/ml, during 5 and 60 seconds, respectively. Eight treatments with a random design and five replica per treatment were used. They were conformed by strains, an absolute control and three treated controls: 1) Tween 80 [0.1%], 2) Sodium hyphocloride [0.5%] + sterile distilled water and 3) Baculovirus phthorimaea. There were differences among controls and treatments for eggs (p= 0.0003), without significant differences among treatments (p=0.0747). We also found mortality differences in larvae (p=0.0001), the Bv006 and Bv007 strains exhibiting the highest mortality (12%). The eggs were more susceptible to fungal isolates. M. anisopliae strains were more effective in reducing the level of consumption of the larvae. Desinfection methods for eggs and larvae and treatment with Tween, did not affect the eclosion of eggs or the larvae viability and could be used in future studies. The low percentage of mortality showed a low biocontrol activity by strains and for this reason these strains could not be selected for the control of this pest. We confirmed the low general potential of entomogenous fungi towards T. solanivora and therefore suggest that other control alternatives, using other organisms such as parasitoids or viruses be sought.
Introducción
Una de las plagas de insectos más dañina de los últimos 10 años en el cultivo de la papa en Colombia es la polilla guatemalteca, T. solanivora (Lepidoptera: Gelechiidae), que afecta la calidad del tubérculo ocasionando pérdidas en campo y almacenamiento hasta del 100% (Arias et al. 1996). Para el manejo de la polilla guatemalteca se dispone de una serie de tecnologías o prácticas de Manejo Integrado de Plagas que incluyen el monitoreo de las poblaciones mediante trampas con feromona sexual, la destrucción de fuentes de infestación y la interrupción del ciclo de vida por medio de prácticas culturales, biológicas y químicas. Las técnicas químicas no han mostrado efectividad, razón por la cual los productores han aumentado la frecuencia de las aplicaciones, elevando los costos de producción, la contaminación ambiental y alterando aún más el agroecosistema. Incluso en algunos casos, debido a la no-efectividad de las medidas de control, se han reducido las áreas sembradas.
Dentro de las alternativas MIP de control biológico se requiere conocer el potencial de los hongos entomopatógenos que se presentan en las zonas donde se encuentran las plagas y que en muchos casos han sido registrados como activos sobre plagas importantes del suelo y de los tubérculos, como es el caso del gusano blanco Premnotrypes vorax (Coleoptera: Curculionidae). Igualmente, dentro de la exploración del uso de cepas fúngicas es útil la evaluación de material con actividad sobre insectos cercanos filogenéticamente al insecto que se desea controlar.
La adaptación de biotipos patógenos a una área climática en particular y la capacidad controladora de éstos son cruciales en el éxito de los entomopatógenos para el control de insectos. Por lo anterior, se decidió evaluar el potencial de cepas de B. bassiana y M. anisopliae nativas del ecosistema papero y con actividad biológica sobre Prenmnotrypes vorax (Hustache) y aisladas de diferentes hospederos como Cargolia arana (Lepidoptera: Geometridae) y Antaeotricha spp. (Lepidoptera: Oecophoridae) para el control de huevos y larvas de 1º ínstar de T. solanivora (Lepidoptera: Gelechiidae), debido a que estos estadios están directamente relacionados con el daño económico. Además, estudios de patogenicidad de B. bassiana y M. anisopliae sobre huevos en condiciones de laboratorio han mostrado porcentajes de mortalidad del 94-100% a los 3.3 días con una concentración de 1x10⁸ conidios/ml sobre Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) (Zúñiga y Redolfi 1981; Fernández y Colmenares 1997).
Materiales y Métodos
En el C.I. Tibaitatá se evaluaron cuatro cepas de B. bassiana y cuatro de M. anisopliae, provenientes del Banco de Germoplasma de Microorganismos del Laboratorio de Control Biológico de Corpoica y del Laboratorio de Entomología de Cenicafé. La selección se hizo de acuerdo con la mayor virulencia que presentaron sobre el gusano blanco de la papa P. vorax (Bv025, Bv003, Bv006, Bv007, Mt001, Mt004, Mt015) y con base en la acción patogénica sobre insectos del orden Lepidoptera (Bv044 y Mt019).
Cada una de las cepas se activó sobre 15 larvas de 3º ínstar de T. solanivora, previamente desinfectadas por inmersión en una solución de hipoclorito de sodio [0.5%] durante 5 segundos con dos enjuagues con agua destilada estéril (ADE) por 30 segundos cada uno; así mismo, los tubérculos se desinfectaron por inmersión en una solución de hipoclorito de sodio [0.05%] durante 1 minuto con tres enjuagues con ADE de 5 minutos cada uno. Posteriormente, se realizó la inmersión de las larvas por 30 segundos y de los tubérculos por 5 minutos en una suspensión fungosa a una concentración de 10⁸ propágulos/ml, la cual fue obtenida mediante conteo en cámara de Neubauer. Se realizaron montajes conformados por un tubérculo y cinco larvas de 3° ínstar en un recipiente de poliuretano estéril. A los 15 días del montaje, las larvas que presentaron mortalidad se llevaron a cámara húmeda a 27°C durante 15 días. Una vez obtenido el hongo esporulado sobre el cuerpo de las larvas, se aisló en Papa Dextrosa Agar (PDA) y se incubó a 27°C durante 15 días. Luego, se hizo un segundo pase para tener el inóculo suficiente para las pruebas de patogenicidad.
Para la prueba de patogenicidad de huevos de T. solanivora se utilizó un diseño completamente al azar con cinco repeticiones por tratamiento, conformados por las diferentes cepas en estudio; además, se utilizaron los testigos tratados Tween 80 [0.1%], hipoclorito de sodio [0.5%] + ADE y testigo absoluto. La unidad experimental estuvo constituida por 20 posturas de tres días de incubación desinfectadas por inmersión en una solución de hipoclorito de sodio [0.5%] por un minuto, con dos enjuagues con ADE por 30 segundos cada uno. Posteriormente, se hizo la inmersión de las posturas en 15 ml de una suspensión fúngica a una concentración de 1x10⁸ propágulos/ml, por 1 minuto, y luego se colocaron en una caja de Petri estéril. Diariamente a las posturas se les adicionó 250 µl de ADE y cuando completaron 11 días de incubación, se hizo la lectura del número de huevos infértiles, fértiles eclosionados y no eclosionados; los huevos y las larvas se llevaron a cámara húmeda a 27°C durante 8 días para determinar la causa de la mortalidad. Para el análisis de los resultados se tomó como variable respuesta el número de huevos fértiles no eclosionados y los datos se transformaron con la raíz cuadrada. La comparación se realizó mediante un análisis de covarianza y una comparación por contrastes ortogonales con el 5% de significancia.
La prueba de patogenicidad sobre larvas de 1° ínstar de T. solanivora se realizó con un diseño completamente al azar con 10 repeticiones por tratamiento (cinco para la evaluación de la patogenicidad y cinco para la estimación del nivel de consumo), conformados por las diferentes cepas en estudio. Además, se utilizaron los testigos tratados con Baculovirus phthorimaea, con Tween 80 [0.1%] y un testigo absoluto. Posteriormente, se hizo la inmersión de las larvas en 15 ml de una suspensión fúngica con una concentración de 1x10⁸ propágulos/ml durante 5 segundos y la de los tubérculos durante 5 minutos. La unidad experimental estuvo constituida por 10 larvas de 1º ínstar, colocadas en la parte superior de un tubérculo depositado en un recipiente de poliuretano estéril. A los 30 días del montaje de la prueba, se contó el número de larvas que puparon y el número de larvas muertas encontradas fuera del tubérculo. Estas últimas se colocaron en cámara húmeda a 27°C durante 8 días, para determinar la causa de la mortalidad.
Para la estimación del nivel de consumo, cada tubérculo se dividió en ocho partes iguales, para cada una de las cuales se asignó un porcentaje de daño, de acuerdo con una apreciación visual. Se calculó el volumen del tubérculo dañado, con el promedio de las ocho partes y se promediaron los datos para cada una de las cinco repeticiones por tratamiento. Los porcentajes de mortalidad de larvas y los promedios de nivel de daño se compararon mediante un análisis de varianza, de contrastes ortogonales y una prueba de rango múltiple de Duncan, todas con el 5% de significancia.
Resultados y Discusión
El efecto de las cepas de B. bassiana y M. anisopliae sobre huevos de la polilla guatemalteca registrado como el nivel de eclosión de huevos fue bajo (Tabla 1). El porcentaje de huevos infértiles se mantuvo dentro de los rangos de fertilidad según Herrera (1997); sin embargo, el número de huevos infértiles se concentró hacia el testigo tratado Tween 80 [0.1%]. A pesar de que las posturas utilizadas en cada tratamiento provenían de una mezcla al azar, es posible que en el testigo con Tween hayan quedado las posturas provenientes de hembras infértiles. García (1980) claramente identifica una acción ovicida como un huevo que no eclosiona ya que ocurre el desarrollo de la larva pero no su salida, por su muerte poco antes de la eclosión. De acuerdo con lo anterior, se descarta la acción ovicida del Tween debido a que en los huevos infértiles no se observó el embrión y además, no se apreciaron chupados o semichupados como ocurre por acción de ovicidas.
Los huevos fértiles no eclosionados se tomaron como la variable respuesta de la acción de los tratamientos biológicos. El análisis para el número de huevos fértiles no eclosionados no mostró diferencias significativas entre los tratamientos con controladores biológicos (p=0.0747) y tampoco entre testigos (p= 0.1893). Las diferencias son significativas únicamente cuando se compara el grupo de testigos con el de controladores biológicos (p= 0.0003). En la figura 1 se muestra la mortalidad corregida del número de huevos fértiles no eclosionados. Las diferencias registradas no son importantes desde el punto de vista de potencialidad para control biológico, pues en procesos de selección de cepas en laboratorio se espera que los porcentajes de mortalidad sean mayores al 75%. En este trabajo, con los tratamientos biológicos se encontró un porcentaje máximo de control de 15.72% y mínimo de 7.84% para las cepas Mt001 y Bv006, respectivamente.
La mortalidad de larvas de 1º ínstar que no entraron al tubérculo y que se trataron con diferentes cepas de B. bassiana y M. anisopliae se muestra en la figura 2, en la cual se aprecia una baja acción de las cepas sobre las larvas de T. solanivora, encontrándose porcentajes de mortalidad inferiores a los cuantificados en la activación de las cepas. Bosa (comunicación personal, 1999), al activar los hongos Paecilomyces fumorosus-roseus, Trichoderma koningii y Penicillium spp. sobre larvas de 3° ínstar de T. solanivora, obtuvo porcentajes de mortalidad de 36%, 7.9% y 5.2%, respectivamente. Estos niveles bajaron a 5.8%, 5.8% y 0%, al evaluar las cepas en larvas de 1° ínstar, a pesar de que las cepas ya estaban activadas. Las diferencias en la susceptibilidad a la infección entre ambos estadíos puede relacionarse con diferencias nutricionales reflejadas en la composición de la epicutícula o con un mayor número de setas que aparecen en las larvas de 3° ínstar y que están directamente relacionadas con la adhesión del conidio a la cutícula (Boucias et al. 1988). También pueden estar asociadas a la existencia de un mayor contacto entre los conidios del hongo en suspensión y la superficie de una larva de mayor tamaño, como la de 3º ínstar comparada con la de 1º ínstar.
Se ha observado que la reactivación mejora la respuesta enzimática. Valdés y Vélez (1998), al comparar la actividad enzimática de aislamientos de B. bassiana no activados y reactivados a través de la broca del café, encontraron diferencias significativas en el tiempo medio de reacción enzimática, y además, comprobaron que enzimas que no se presentaban antes de la reactivación, se comenzaban a expresar después de ésta. Sin embargo, no siempre la activación puede promover una mejor respuesta enzimática, ya que ésta depende principalmente de la especificidad entre el patógeno y su hospedero y de la virulencia de la cepa.
El análisis de varianza para el número de larvas muertas fuera del tubérculo mostró que existen diferencias en la mortalidad para las cepas de B. bassiana y M. anisopliae (p= 0.0001); además, se encontraron diferencias significativas entre las cepas de B. bassiana (p= 0.0026). De acuerdo con la prueba de rango múltiple de Duncan, se determinó que existían diferencias entre la mortalidad causada por la cepa Bv025, y las cepas Bv006 y Bv007, no encontrándose diferencias significativas entre las dos últimas.
La patogenicidad baja hacia huevos y larvas de 1º ínstar de T. solanivora puede explicarse en términos de especificidad de cepas; la literatura indica que los hongos entomopatógenos pertenecientes a la clase Hyphomycetes atacan un amplio rango de hospederos. Sin embargo, los aislamientos de B. bassiana aisladas de especies particulares de insectos, probados sobre un rango de especies en laboratorio, tienen un estrecho rango de hospederos, pudiendo ser relativamente específicos (McCoy et al. 1988). Los hongos entomopatógenos tienen la capacidad de especializarse en su acción patogénica en los insectos a los cuales se les aplica el control (Valdés y Vélez 1998), planteándose la existencia de cierta especificidad entre el hospedero y el hongo. Para Evans (1992), el mayor grado de infección de los hongos entomopatógenos se debe a que éstos han sufrido un proceso de selección natural y coevolución con su hospedero.
En este trabajo las cepas provenientes de especies del orden Lepidoptera, no ocasionaron mortalidad sobre larvas de T. solanivora. Hasta el momento no ha sido posible establecer una relación clara entre la virulencia de la cepa y el tipo de hospedero, aunque en algunos casos, la cercanía filogenética origina una mayor virulencia. Maurer et al. (1997), encontraron que cepas de B. bassiana aisladas de Ostrinia nubilalis (Lepidoptera: Pyralidae) presentaron alta mortalidad (65-100%) sobre individuos del mismo hospedero y niveles de sólo el 30% para aislamientos de otros Pyralidae, con excepción de un aislamiento proveniente de Sitona spp. (Coleoptera), el cual presentó una alta mortalidad atribuida a la estrecha relación genética de esta сера con los aislamientos altamente virulentos. Castro (1997) encontró que cepas de B. brogniartii, aisladas de Premnotrypes latitorax, a pesar de pertenecer al mismo género del gusano blanco, fueron menos patogénicas en larvas de 1º ínstar de P. vorax que las cepas provenientes de homópteros. Varela y Morales (1996) señalan que aislamientos de B. bassiana de Coleoptera, mostraron menor virulencia contra Hypothenemus hampei que los aislados de materia orgánica en descomposición, lo que sugiere que el origen del hospedero probablemente no es un buen indicador de la virulencia de un aislamiento.
La especificidad de cepas de hongos entomopatógenos también se ha asociado con la composición química de la cutícula de los insectos. En Lepidoptera se han identificado compuestos como el aldehído (E)-2-decenal, que a pesar de servir como puente entre la cutícula y el conidio del hongo, no es un elicitor de la germinación. En muchos casos la actividad fungistática se ha asociado con la presencia de lípidos tóxicos y compuestos fenólicos (Sosa et al. 1997).
Respecto al nivel de consumo de las larvas, se encontraron diferencias en las tratadas con cepas de B. bassiana en comparación con las tratadas con M. anisopliae (p= 0.0008). No existen diferencias significativas entre los testigos tratados (p= 0.76); las diferencias fueron significativas cuando se comparó el grupo de testigos con los tratamientos de hongos controladores (p= 0.008). Finalmente, las cepas de M. anisopliae son más efectivas para atacar al insecto que las de B. bassiana (p<0.05), por lo cual se reduce el nivel de consumo del tubérculo, lo que contrasta con los resultados de la prueba de patogenicidad de larvas de 1° ínstar. Esto sugiere que no existe una relación directa entre el número de larvas que atacan el tubérculo y el nivel de consumo que ocasionan en el mismo, cuando existe la acción del entomopatógeno. Aunque las cepas de B. bassiana ejercieron una acción inmediata sobre un mayor número de larvas de 1º ínstar, las larvas que sobrevivieron causaron un mayor daño que las larvas tratadas con M. anisopliae; posiblemente en éstas se manifestó en una menor actividad larval debido a que la larva posiblemente utiliza gran parte de su energía en respuestas de defensa contra el hongo.
Eclosión de huevos de Tecia solanivora bajo el efecto de cepas de Beauveria bassiana y Metahrizium anisopliae
Porcentajes de eclosión de huevos a los 11 días de incubación
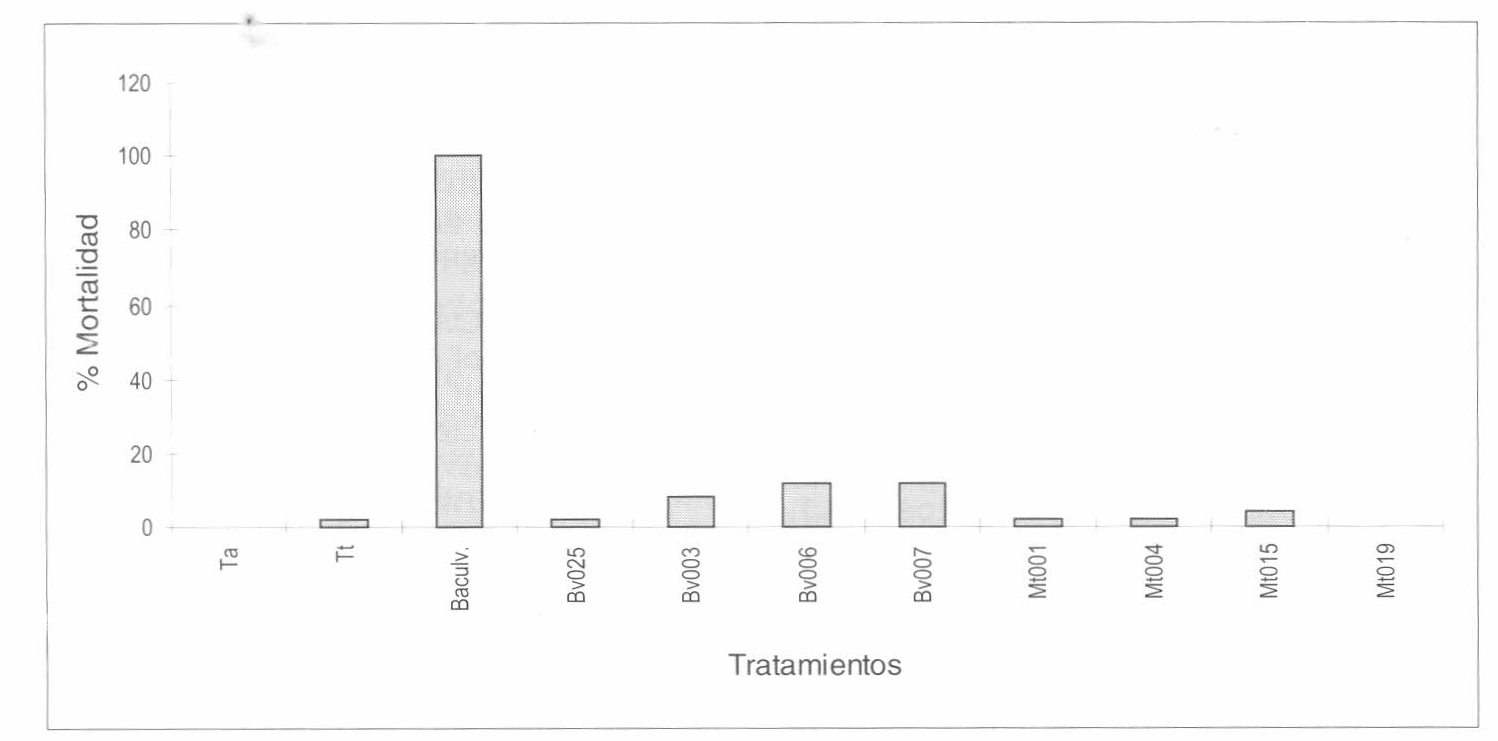
Mortalidad de huevos fértiles de Tecia solanivora bajo el efecto de cepas de Beauveria bassiana y Metarhizium anisopliae, en comparación con los controles tratados y negativo.
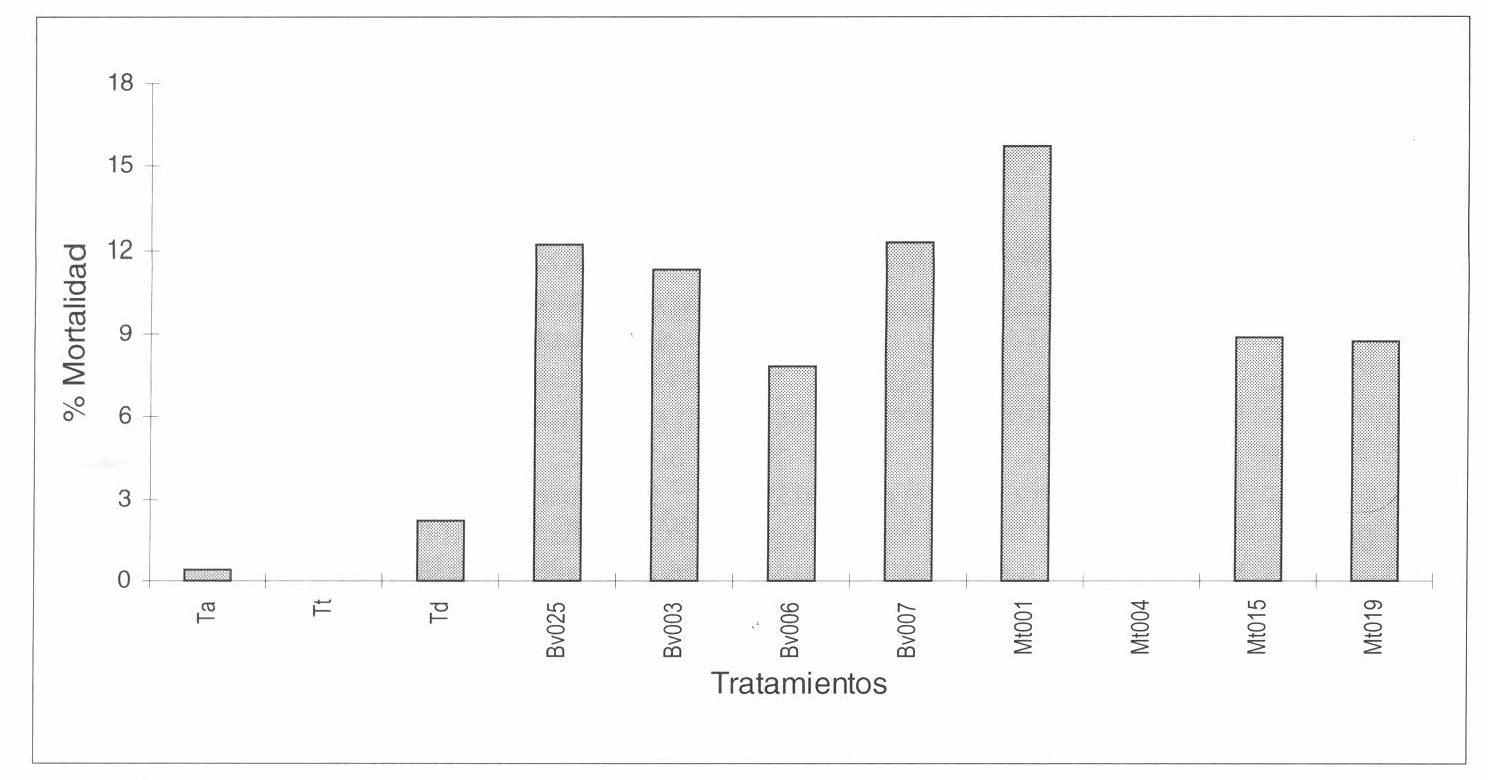
Mortalidad de larvas de 1º ínstar de Tecia solanivora fuera de tubérculo bajo el efecto de cepas de Beauveria bassiana y Metarhizium anisopliae, en comparación con los controles tratados y negativo.
Conclusiones
Existe una baja actividad biocontroladora de las cepas de B. bassiana y M. anisopliae evaluadas sobre la polilla guatemalteca.
Las técnicas de desinfección y el tratamiento con Tween no afectan la eclosión de huevos fértiles y la viabilidad de larvas de 1º y 3° ínstar de T. solanivora. Estas técnicas pueden ser empleadas para ensayos y estudios futuros con esta especie plaga.
Los huevos presentan mayor susceptibilidad a las cepas de B. bassiana y M. anisopliae, que los otros estadios de T. solanivora evaluados.
Las larvas de 1º ínstar de T. solanivora son más susceptibles a cepas de B. bassiana que a cepas de M. anisopliae, siendo las cepas Bv006 y Bv007 las que ocasionan mayor mortalidad.
Las cepas de M. anisopliae son más efectivas para reducir la actividad larval de T. solanivora que las de B. bassiana; sin embargo, estas dos especies presentan menor actividad patogénica que Baculovirus phthorimaea, altamente efectivo sobre larvas de 1º ínstar de T. solanivora.