
Abstract
Infected insects with fungus were collected in cassava crops in the Cauca Valley. In the laboratory, the fungus Beauveria bassiana (B.b.9601 and B.b.9602) and Aspergillus (Asp.9601) were isolated, purified and identified. These native isolates together with one of Metarhizium anisopliae (M.a.9206), received from CENICAFE, were evaluated on larvae of E. ello. The methodology to select the more pathogenic isolates and the best method of inoculation between aspersion on larvae, leaf and larvae-leaf was determined. The most susceptible larval Instar of the selected isolate was also determined to establish the LD50. The individual separation of the treated insects in cardboard boxes with plastic sheet of 14 x 11.5 x 5 cm with cap was the most adequate methodology to do pathogenic tests on E. Ello and was denominated "Experimental Unit Múnera De Los Rios". B.b.9601 was the most pathogenic isolate causing mortality between 77.5% and 87.5% presenting significant differences (P<0.05) compared to the other isolates; M.a 9206 caused mortalities no greater than 67.9%; B.b.9602 caused a lower mortality of 16.1% and Asp.9601 wasn't pathogenic. There was no significant difference between inoculation methods (P<0.05); choosing aspersion to larval-leaf to be most similar to field application. Instar III showed greatest susceptibility to B.b.9601. The LD50 of B.b.9601 was 1x104.8018 viable conidias ml−1 with confidence limits between 1x100.3523 and 1x107.4120. B.b.960 sprinkled on fifth instar larvae produced a mortality of 40%, the survivors showed a normal oviposition and eggs with high viability. B.b.9601 inoculated on eggs doesn't caused considerably mortality. The aspersion of a mixture of B.b.9601 and M.a 9206 caused 90% mortality in III ínstar larvae.
Introducción
El "Gusano Cachón de la Yuca" es considerado una de las plagas más importantes de la yuca debido a su alta capacidad de consumo foliar, tejido meristemático e incluso porciones del tallo (20 - 30 cm) especialmente en las últimas etapas de su fase larval (Bellotti et al. 1983; Bellotti y Arias 1977). Cuando la defoliación ocurre durante los primeros meses del cultivo puede ocasionar pérdidas en el rendimiento entre 10 y 50% y las plantas jóvenes pueden morir (Farias 1990). Según investigaciones realizadas en la localidad de Santander de Quilichao y Palmira estas disminuciones fueron hasta del 64% con dos ataques continuos en suelos pobres y hasta el 47% en suelo fértil (Arias y Bellotti 1984).
Los agricultores reaccionan frecuentemente a las infestaciones de Cachón con aplicaciones excesivas de Parathión etílico y Triclorfón, lo cual trae consecuencias negativas para el equilibrio biológico del ecosistema, más aún cuando se aplican insecticidas teniendo bajos niveles de infestación de la plaga, en mezclas inadecuadas, en dosificaciones incorrectas, y en épocas inapropiadas.
El desarrollo de poblaciones resistentes, la reaparición de ataques más severos, la conversión de plagas secundarias en plagas de importancia económica, la eliminación de los enemigos naturales benéficos y la contaminación general del medio ambiente en forma de residuos químicos nocivos para la fauna y para el hombre, son consecuencias directas del empleo del control químico sin las debidas precauciones (Bellotti et al. 1989).
Se han investigado formas de control como rotación, preparación de terreno y control de malezas para destruir larvas, pupas y hospedantes alternos (Montaldo 1981; Lozano et al. 1981); el empleo de Trichogramma spp., Telenomus sphingis, Apanteles sp., Euplectrus sp., Podisus sp., Polistes sp. y Zellus sp. (Bellotti et al. 1989); también Bacillus thurigiensis y virus (Belloti 1994). Bellotti y Arias citados por Bellotti et al. (1992) registran un gran porcentaje de pupas de E. ello destruidas por un hongo clasificado como Cordyceps sp en áreas de Armenia y Caicedonia, Colombia, durante la estación lluviosa, siendo esta la única información existente con respecto a hongos entomopatógenos en cultivos de yuca.
Teniendo en cuenta que hasta el momento no se había realizado ningún estudio para evaluar la patogenicidad de hongos entomopatógenos sobre esta plaga, se propusieron como objetivos de esta investigación: 1) obtener este tipo de hongos provenientes de cultivos comerciales de yuca en el departamento del Valle del Cauca, 2) establecer una metodología de evaluación de la patogenicidad en condiciones de laboratorio y 3) determinar, entre los aislamientos nativos y un aislamiento de M. anisopliae previamente evaluado, cuál presenta la mayor patogenicidad; 4) el ínstar más susceptible a él y 5) la DL50.
Materiales y Métodos
La investigación se desarrolló en el laboratorio de Entomología de Yuca del CIAT a una temperatura promedia de 25°C y 60% de humedad relativa. Los aislamientos se obtuvieron mediante recolección de insectos afectados por hongos en cultivos de yuca localizados en los municipios de Palmira, Calima, Jamundí, Caicedonia y La Cumbre en el Valle del Cauca; los insectos se identificaron en la colección entomológica de la Universidad Nacional de Colombia, sede Palmira.
Los hongos se aislaron, purificaron y sembraron en medio nutritivo artificial Sabouraud Dextrosa Agar (SDA) y crecieron a temperatura ambiente para luego ser identificados mediante las claves taxonómicas de Barnet y Hunter (1972), Poinar y Thomas (1984) y Samson (1981), lo cual se confirmo en la Disciplina de Entomología de CENICAFE y en el laboratorio de Microbiología Vegetal de la Universidad Nacional de Colombia, sede Palmira. Los aislamientos se probaron sobre E. ello junto con el aislamiento M.a.9206 de M. anisopliae aislado de Aenolamia reducta y cedido por CENICAFE.
Los especímenes de E. ello se obtuvieron de la colonia establecida mediante la metodología de Arias y Reyes (1983) modificándose la unidad de cría por cajas de cartón de 50 x 50 x 40 cm con 30 larvas cada una y plántulas de yuca, ubicadas en cabinas climatizadas entre 18° y 20° C para conservar la humedad del alimento y evitar el aumento de la temperatura; al finalizar cada ciclo las cajas se desecharon. Esta unidad de cría redujo la mortalidad por hacinamiento y enfermedades entre un 80 - 90% y por heridas producidas por el roce entre las lavas en un 80% al compararse con la unidad de cría propuesta en la metodología original, lo cual ratifica lo expuesto por Watanabe (1987) quien señala que altas densidades de población incrementan el contacto entre hospederos sanos y los infectados aumentando la susceptibilidad a la infección debido a la competencia por alimento, también la temperatura y la humedad son factores físicos externos importantes para la susceptibilidad y multiplicación de patógenos en el hospedero.
El inóculo utilizado en cada uno de los experimentos se obtuvo cosechando las conidias de tres cajas petri (100 x 15 mm) y realizando una suspensión con 25 ml de agua destilada estéril más Tween 40 al 0.1% (ADET). La concentración se determino mediante diluciones sucesivas y la cámara de Newbauer, luego se desarrollaron pruebas de germinación para determinar la viabilidad de las conidias y expresar la concentración en conidias viables ml−1.
Determinación de Metodología
Se verificó la patogenicidad de los aislamientos empleando tres métodos de inoculación: 1) aspersión a larva, consistió en aplicar con aspersor de Vilbis, a dos libras de presión, la solución de conidias obteniendo una nube homogénea sobre el insecto; 2) contacto tarsal, consistió en hacer caminar grupos de 10 larvas durante dos minutos en cajas Petri esterilizadas que contenían 0.5 ml de solución de conidias y 3) contacto de superficie, se aplicó la solución de conidias sobre el dorso de la larva con una pipeta Pasteur esterilizada. Se utilizó un diseño completamente al azar con 20 repeticiones de una larva de tercer ínstar para cada método de inoculación, teniendo 60 larvas por aislamiento y un testigo común de 20 larvas, para un total de 320. En todos los experimentos el testigo se inoculó con ADET y se evaluó la mortalidad diariamente hasta cuando los insectos sobrevivientes alcanzaron el estado de adulto, tomándose como individuo muerto por hongo a la larva que presentó crecimiento típico del microorganismo aplicado al colocarse en cámara húmeda después de la muerte; de este crecimiento se montaron placas al microscopio verificando que fuera el hongo aplicado. Los resultados se cuantificaron como cantidad de individuos muertos.
Bell (1974) y Burge (1988) definen reactivación como la acción de pasar el hongo varias veces a través de un hospedero para así incrementar la virulencia atenuada por repetidas siembras en medio nutritivo artificial. Teniendo en cuenta esta definición se realizaron 2 reactivaciones sucesivas de los aislamientos que mostraron patogenicidad utilizando los métodos de inoculación anteriormente descritos. Se aprovechó la primera reactivación para corroborar la patogenicidad de los hongos implementando el mismo diseño y evaluación del experimento anterior. En la segunda se utilizó un diseño completamente al azar con tres repeticiones de 20 larvas de tercer ínstar por método de inoculación para un total de 180 larvas por aislamiento y 720 larvas para todo el experimento. Las concentraciones expresadas en conidias viables por mililitro fueron para B.b.9601, 6x109; B.b.9602, 8.7x109 y M.a.9206, 1x109. Los resultados en la segunda reactivación se expresaron en porcentajes de mortalidad para cuantificar el incremento de la patogenicidad. Luego los hongos se aislaron y almacenaron bajo la técnica de papel filtro propuesta por Aricapa y Correa (1990), dando igualdad de condiciones a los aislamientos y así tener disponibilidad de ellos para el desarrollo de la investigación.
Por otra parte, se determinó la unidad experimental para el desarrollo de los bioensayos definitivos. Para esto se realizaron experimentos preliminares evaluando, en primera instancia, la mortalidad por enfermedades y otras causas utilizando larvas de segundo ínstar individualizadas en 50 bandejas plásticas de 30 x 20 x 10 cm esterilizadas con luz ultravioleta durante 12 horas y 50 cajas de cartón de 14 x 11.5 x 5 cm con tapa. Se evaluó diariamente hasta la emergencia de adultos cuantificándose el porcentaje de mortalidad. Seguidamente se dispusieron 50 cajas plastificadas de cartón de 14 x 11.5 x 5 cm con tapa y 50 cajas del mismo tipo sin plastificar con un foliolo de yuca dentro de cada una de ellas, constituyéndose como una repetición para un total de cinco. Después de dos días se determinó en cuál de los dos tipos de caja las hojas estaban más frescas, garantizando a las larvas mayor tiempo de disponibilidad del alimento.
Selección de aislamientos entre B.b.9601, B.b.9602 y M.a.9206
Se empleó un diseño completamente al azar con cuatro repeticiones de 20 larvas de tercer ínstar (80 larvas por método de inoculación) para un total de 240 larvas por aislamiento y 960 larvas para todo el ensayo, utilizando la unidad experimental estandarizada. Los resultados se expresaron en porcentajes de mortalidad y se realizó un análisis de varianza con test de Rangos Múltiples de Duncan. Las concentraciones en conidias viables ml−1 fueron: B.b.9601, 5.9787x109; B.b.9602, 7.5168x109 y M.a.9206, 6.342x109.
Teniendo en cuenta el hábitat de E. ello donde las larvas utilizan como alimento hojas de yuca, se emplearon los siguientes métodos de inoculación: 1) aspersión a larva, consistió en aplicar la solución de conidias con aspersor de Vilbis a 2 libras de presión obteniendo una nube homogénea sobre el insecto; 2) aspersión a hoja, consistió en asperjar la solución de conidias, bajo las mismas condiciones del método anterior, sobre hojas de yuca utilizadas como alimento para las larvas sanas del tratamiento y 3) aspersión a larva-hoja, consistió en asperjar la suspensión de conidias, bajo las mismas condiciones anteriores, sobre el conjunto larva hoja de yuca.
Selección entre B.b 9601 y M.a.9206
Se implementó un diseño completamente al azar con cuatro repeticiones de 10 larvas de tercer ínstar cada uno (40 larvas por método de inoculación) para un total de 120 larvas por aislamiento y 360 larvas para todo el ensayo. Los resultados se expresaron en porcentajes de mortalidad y se realizó un análisis de varianza con test de Rangos Múltiples de Duncan; también se escogió el método de inoculación más adecuado. Las concentraciones utilizadas en conidias viables ml−1 para M.a.9206 y B.b.9601 fueron 4.30x109 y 1.76x109 respectivamente. Los métodos de inoculación fueron los mismos que se utilizaron en la selección anterior, al igual que la unidad experimental.
Determinación del ínstar larval más susceptible a B.b.9601
Se emplearon larvas de segundo, tercero, cuarto y quinto ínstares, cada ínstar con un testigo. No se evaluó el primer ínstar, debido a que su tamaño pequeño ofrece alto riesgo de mortalidad por lesiones causadas durante la manipulación, evitando así falsos resultados de mortalidad, debido a que el hongo puede colonizar el cadáver del insecto enmascarando la causa real de la muerte. Se empleó una concentración de 3.75 x 1010 conidias viables ml−1 y el método de inoculación utilizado fue aspersión a larva - hoja. Se adoptó un diseño completamente al azar, cada ínstar con cuatro repeticiones de 10 larvas cada una (80 larvas por ínstar incluyendo su testigo) para un total de 320 larvas en todo el ensayo; la unidad experimental fue igual a la del experimento anterior. Se realizó un análisis de varianza y test de Rangos Múltiples de Duncan.
Determinación de la dosis letal media de B.b.9601
Se realizó un diseño completamente al azar en el que cada concentración representó un tratamiento y en cada uno se evaluaron 10 larvas de tercer ínstar con cuatro repeticiones para un total de 400 insectos. Se utilizaron nueve concentraciones del inóculo cuyo logaritmo en base 10 fue de 2 a 10. El método de inoculación y la unidad experimental fueron iguales a las del experimento anterior. Los datos obtenidos se sometieron a un análisis Probit para calcular la DL50.
Efecto de la aplicación en quinto ínstar sobre adultos y viabilidad de huevo
Se inocularon larvas de quinto ínstar con B.b.9601 a una concentración de 8.52 x 1010 conidias viables ml−1 evaluándose el desarrollo hasta la oviposición, al igual que la viabilidad de los huevos. Se utilizó un diseño completamente al azar con seis repeticiones de 10 larvas teniendo un total de 120 larvas en todo el ensayo. La evaluación se realizó diariamente y cuando los sobrevivientes alcanzaron el estado de pupa se ubicaron en jaulas de emergencia, los adultos se llevaron a jaulas de oviposición. Los huevos se recolectaron diariamente y se colocaron en cajas de Petri estériles para su eclosión. Los resultados se expresaron porcentual-mente. El método de inoculación utilizado fue aspersión a larva hoja y la unidad experimental fue la misma del experimento anterior.
Efecto de la aplicación de B.b.9601 sobre huevo
Se empleó un diseño completamente al azar inoculando posturas con B.b.9601 en una concentración de 9.9 x 109 conidias viables ml−1, ubicadas en hojas de yuca provenientes de las jaulas de cópula de la colonia. Se evaluó el porcentaje de mortalidad causado por el hongo hasta cuando las larvas emergidas alcanzaron el segundo ínstar. Cada hoja de yuca, con 10 posturas de E. ello, se depositó dentro de una caja Petri estéril después de la inoculación, constituyendo una repetición. Cada tratamiento (B.b.9601 y testigo) tuvo 20 repeticiones para un total de 400 huevos en todo el ensayo. La evaluación se realizó diariamente.
Efecto de la mezcla B.b.9601 y M.a.9206 sobre larvas de tercer ínstar
Se inocularon larvas de tercer ínstar con una mezcla de B.b.9601 y M.a.9206 cuyas concentraciones fueron 3.4 x 1010 y 1.6 x 1010 conidias viables ml−1 respectivamente, mediante aspersión larva - hoja. Se implementó un diseño completamente al azar con seis repeticiones de 10 larvas cada una, para un total de 60 larvas por tratamiento (mezcla y testigo) y 120 en todo el ensayo. La mortalidad se expresó porcentualmente.
Resultados y Discusión
De las cuatro zonas evaluadas solamente en el municipio de La Cumbre se hallaron los siguientes insectos afectados con hongos:
En la vereda El Carmen se encontró un lepidóptero de la especie E. ello que presentó crecimiento de un hongo verde oliva perteneciente al género Aspergillus sp. la identificación fue corroborada en el laboratorio de Microbiología Vegetal de la Universidad Nacional de Colombia, sede Palmira, codificándose como Asp. 9601.
En la vereda Jiguales se encontraron adultos de un coleóptero del género Curinus (Col: Coccinelidae) presentando un crecimiento algodonoso de color blanco, el cual se ubicó sobre la parte abdominal y dorsal (entre y sobre los élitros) del insecto, el cual fué identificado como B. bassiana, identificación confirmada por la Disciplina de Entomología de CENICAFE y codificado como B.b.9602. En la misma vereda se encontró una larva de un lepidóptero de la familia Arctiidae con crecimiento felposo de color blanco con estructuras que salían del interior del individuo, el cual fué identificado como B. bassiana, confirmado por la Disciplina de Entomología de CENICAFE y codificado como B.b. 9601. En los lugares donde se recolectaron los especímenes afectados no se realizaron aplicaciones de productos biológicos, lo que fue corroborado por el Comité de Cafeteros del Municipio de La Cumbre que se encarga del suministro y aplicación de B. bassiana para el control de la broca, por lo tanto se puede decir que estos microorganismos son nativos de la zona.
Determinación de Metodología.
Los aislamientos que presentaron patogenicidad a E. ello fueron B.b.9601 con cinco individuos muertos, B.b.9602 con cuatro individuos muertos y M.a.9206 con 18 individuos muertos, los cuales manifestaron signos típicos de las muscardinas blanca y verde. El aislamiento Asp.9601 no presentó patogenicidad sobre los insectos tratados, por lo que no se incluyó en los siguientes experimentos. En el testigo se obtuvo una mortalidad de 10 individuos sin presencia de hongo, considerada como mortalidad natural.
Los aislamientos probados en la primera reactivación manifestaron patogenicidad sobre E. Ello. En cuanto a la segunda reactivación el porcentaje de mortalidad en todo el experimento fue del 25.45% distribuido de la siguiente forma: M.a.9206 43.8%, B.b.9601 23.3% y B.b.9602 16.1%. El testigo presentó 18.3% de mortalidad natural (Tabla 1). Los porcentajes de mortalidad causados por M. anisopliae sobre E. ello se deben a características propias ya que en ensayos realizados por Sánchez (1996), se inoculó este aislamiento sobre Cyrtomenus bergi, obteniendo mortalidades mayores al 63%.
Porcentaje de mortalidad en la reactivación y selección de los aislamientos de Beauveria bassiana y Metarhizium anisopliae en condiciones de laboratorio*
Temperatura promedio 25°C; Humedad relativa promedia 60%
Promedios con la misma letra no son estadísticamente diferentes según Test de Rangos Múltiples de Duncan
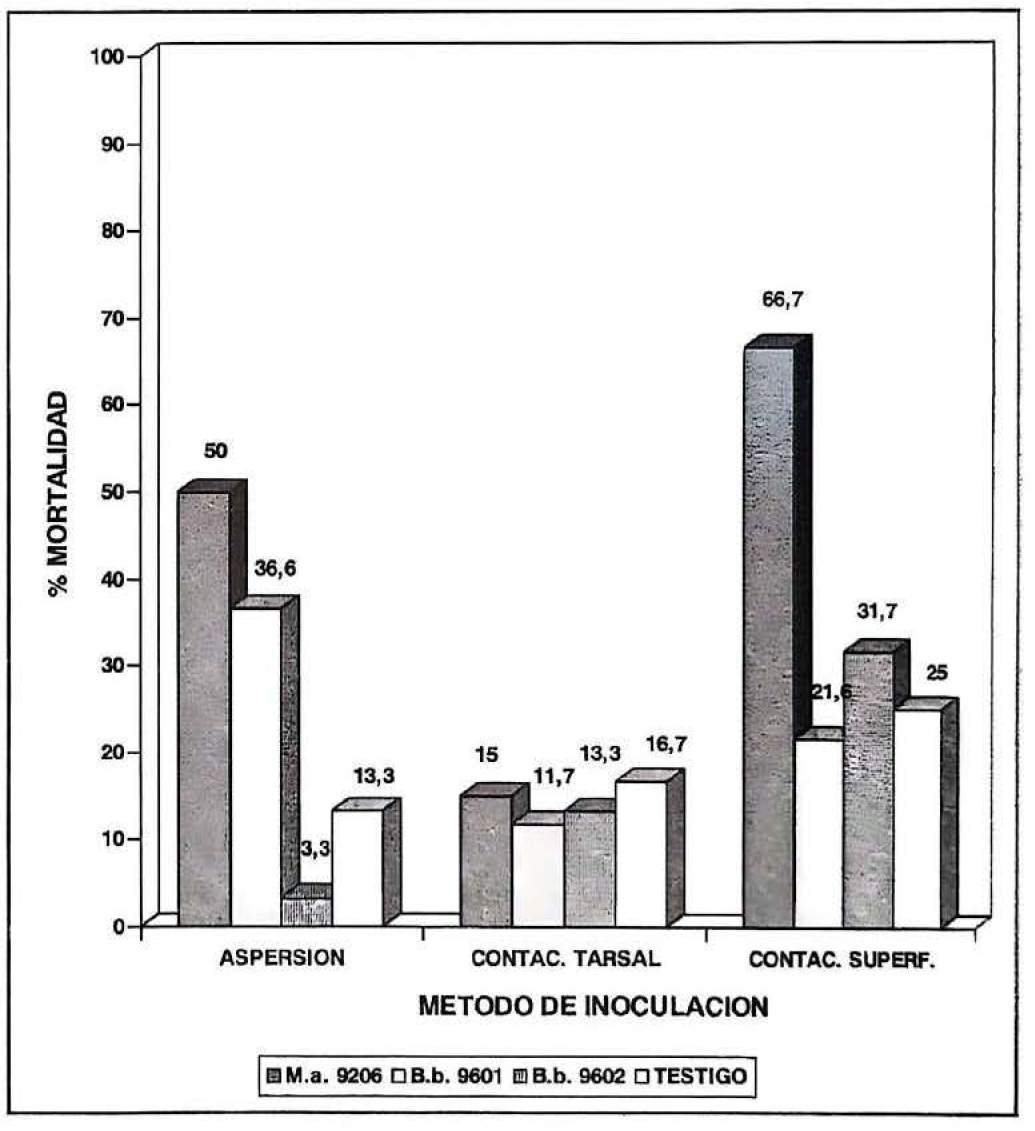
Porcentajes de mortalidad por método de inoculación en la reactivación de Beauveria bassiana (B.b. 9201 y B.b. 9602) y Metarhizium anisopliae (M.a. 9206).
En cuanto a los métodos de inoculación, la mayor mortalidad se presentó al utilizar contacto de superficie, con 66.7% y 31.7% de mortalidad para M.a.9206 y B.b.9602 respectivamente. El aislamiento B.b.9601 causó mayor mortalidad con el método aspersión a larva, con un 36.6%. Con lo anterior se pudo concluir que para efectos de reactivación de M.a.9206 y B.b.9602 el método de inoculación más adecuado es contacto de superficie y para B.b.9601 aspersión a larva. El método de inoculación contacto tarsal fue el menos efectivo (Fig. 1).
En los experimentos preliminares para la determinación de la unidad experimental, un 6% de mortalidad se presentó en las larvas ubicadas en las cajas de cartón mientras que todas las larvas ubicadas en bandejas plásticas resultaron muertas, por lo que se presume la contaminación de las bandejas por patógenos debido a la utilización previa en diferentes experimentos. Por otro lado, las cajas de cartón plastificadas mostraron mejores resultados que las cajas sin recubrimiento plástico manteniendo la humedad de las hojas en su interior por un mayor tiempo. Por lo anterior se estandarizó el uso de larvas individualizadas en cajas de cartón plastificadas de 14 x 11.5 x 5 cm con tapa como unidad experimental para todos los experimentos, denominando ésta metodología "Unidad Experimental Múnera – De Los Ríos".
Selección de aislamientos entre B.b.9601, B.b.9602 y M.a.9206.
Se presentaron diferencias significativas (P < 0.05) entre los aislamientos con respecto a la mortalidad entre los insectos tratados y el testigo. El aislamiento B.b.9601 causó 31.6% de mortalidad, seguido de M.a.9206 con 25 % y B.b.9602 con 11.25%, en el testigo se observó un 5.45% de mortalidad. Según la prueba de Rangos Múltiples de Duncan B.b.9602 fue igual estadísticamente al testigo y B.b.9601 fue igual a M.a.9206, y teniendo en cuenta que B.b.9602 causó el porcentaje de mortalidad más bajo, se decidió realizar una nueva selección bajo la misma metodología pero solo entre B.b.9601 y M.a.9206 (Tabla 1). La baja patogenicidad de B.b.9602 puede ser explicada por la variación tanto entre aislamientos de una misma especie de hongo, como también entre el potencial de las toxinas que juegan un papel importante en el ataque de los insectos, presentando baja patogenicidad llegando a no controlar el hospedero concordando con lo determinado por Burge (1988).
Porcentaje de mortalidad y comportamiento de los aislamientos con respecto a los métodos de inoculación empleados en la selección entre aislamientos de Beauveria bassiana y Metarhizium anisopliae en condiciones de laboratorio*
Temperatura promedio 25°C; Humedad relativa promedia 60% Promedios con la misma letra no son estadísticamente diferentes según Test de Rangos Múltiples de Duncan
Se presentaron diferencias significativas (P < 0.05) entre métodos de inoculación con respecto al porcentaje de mortalidad. El método aspersión larva - hoja presentó el mayor porcentaje de mortalidad con respecto a los demás métodos (33.7%), con lo que se presume un aumento en la patogenicidad de los aislamientos por intervenir al mismo tiempo aspersión a larva y aspersión a hoja. El método de inoculación que presentó el menor porcentaje de mortalidad fue aspersión a larva (7.8%) (Tabla 2). Según la prueba de Rangos Múltiples de Duncan los métodos de aspersión a larva y aspersión a hoja no mostraron diferencias significativas y difieren al método de aspersión a larva hoja, por lo que se concluye una posible interacción entre método de inoculación y la patogenicidad del aislamiento (Tabla 2).
Selección entre B.b.9201 y M.a.9206.
La distribución de los datos obtenidos en este experimento no permitieron realizar ninguna determinación, por lo que se decidió realizar una réplica bajo las mismas condiciones y tener en cuenta los resultados de ambas experiencias para la selección. Para cada una de estas B.b.9601 causó el mayor porcentaje de mortalidad con 86.6% y 70%, seguido de M.a.9206 con 71.7% y 64.1%, el testigo presentó 8.16% y 2.5% (Tabla 2). Promediando los resultados, B.b.9601 causó el mayor porcentaje de mortalidad con 78.3%, mientras que M.a.9206 causó un 67.91% y el testigo presentó un 5.33% de mortalidad natural, presentando una diferencia significativa entre ellos (P < 0.05) (Tabla 1). Según lo anterior, y teniendo en cuenta la prueba de Rangos Múltiples de Duncan donde los aislamientos se ubicaron en grupos diferentes dependiendo de su media, se seleccionó a B.b.9601como el aislamiento más patogénico sobre E. ello, sin indicar con esto que el aislamiento de M.a.9206 no sea patogénico y promisorio para el control del insecto.
Comparación de métodos de inoculación con respecto a las réplicas en la selección de los aislamientos de Beauberia bassiana (B.b. 9601) y Metarhizium anisopliae (M.a. 9206) en condiciones de laboratorio*
Temperatura promedio 25°C; Humedad relativa promedia 60%
El método aspersión a larva presentó el mayor porcentaje de mortalidad con 54.1%, seguido de aspersión a larva - hoja con 51.2% y por último aspersión a hoja con 46.2% de mortalidad (Tabla 3), lo cual no coincide con lo observado en la selección anterior, ya que la mayor mortalidad se presentó en aspersión a larva - hoja. Se seleccionó este último como el método mas adecuado para experimentos con hongos entomopatógenos sobre E. ello ya que el contacto del inóculo tanto con la larva como con la hoja se asemeja a lo que sucedería con la aspersión de una suspensión de conidias en un cultivo, además, la prueba de Rangos Múltiples de Duncan para métodos de inoculación determinó que aspersión a larva - hoja se comportó estadísticamente igual a los otros dos métodos de inoculación.
Determinación del ínstar larval más susceptible a B.b.9601.
Se presentaron diferencias significativas (P < 0.05) para la variable mortalidad en los ínstares evaluados de E. ello, como también entre las larvas inoculadas con B.b.9601 y el testigo. El ínstar que presentó mayor porcentaje de mortalidad fue el tercero con 87.5%, mientras que el quinto mostró el menor porcentaje de mortalidad con 27.5% (Fig. 2). Ferron (1985) observó que M. anisopliae causó el mayor porcentaje de mortalidad en el ínstar tercero de Oryctes monoceros, lo que coincide con los resultados obtenidos en este experimento. Esto puede atribuirse a diferentes mecanismos de defensa por parte de las larvas, según Watanabe (1987), éstos pueden ser de índole cuticular durante la penetración del hongo, factores en la hemolinfa contra el crecimiento del patógeno y resistencia por parte del insecto a los metabolitos tóxicos producidos por el hongo. Por otra parte, Tanada y Kaya (1993) afirman que la composición de aminoácidos de la superficie del integumento de las larvas varía según el ínstar y el tiempo antes de la muda. Los mismos autores citan que estos aminoácidos junto con glucosaminas inician la germinación y crecimiento de B. bassiana sobre Heliotis zea. Lo anterior podría explicar la susceptibilidad del tercer ínstar a B.b.9601 al suponer que la composición de su integumento en un momento dado permitió la efectiva penetración y por lo tanto causar mayor mortalidad.
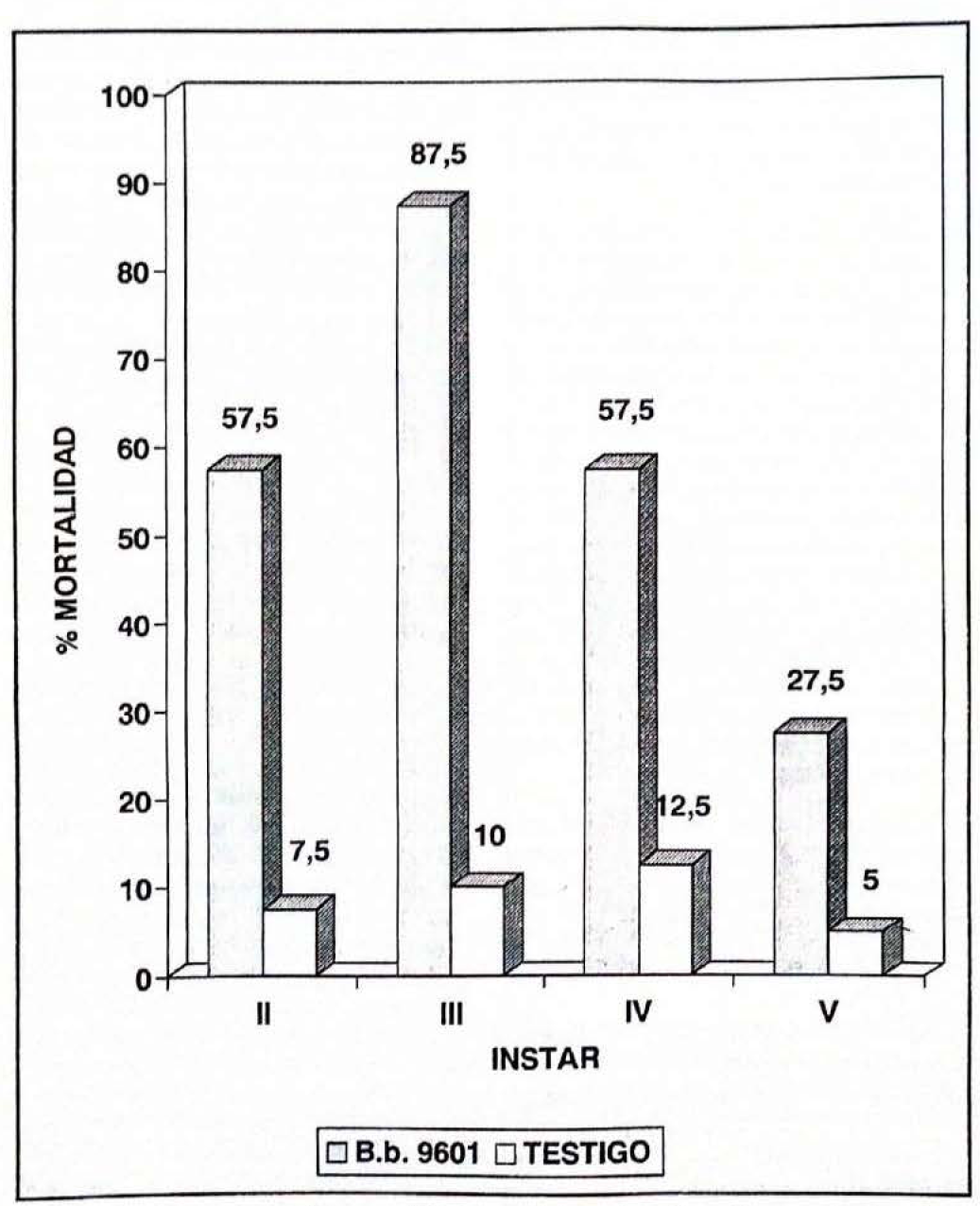
Porcentajes de mortalidad por ínstar al aplicar Beauveria bassiana (B.b.9601) sobre Erinnyis ello.
Determinación de la dosificación letal media de B.b.9601.
El 50% de mortalidad en las larvas de tercer ínstar de E. ello se obtuvo con una concentración de 1x104.8018 conidias viables ml−1 con un límite de confianza de 1x100.3523 y 1x107.4120 (Fig. 3).
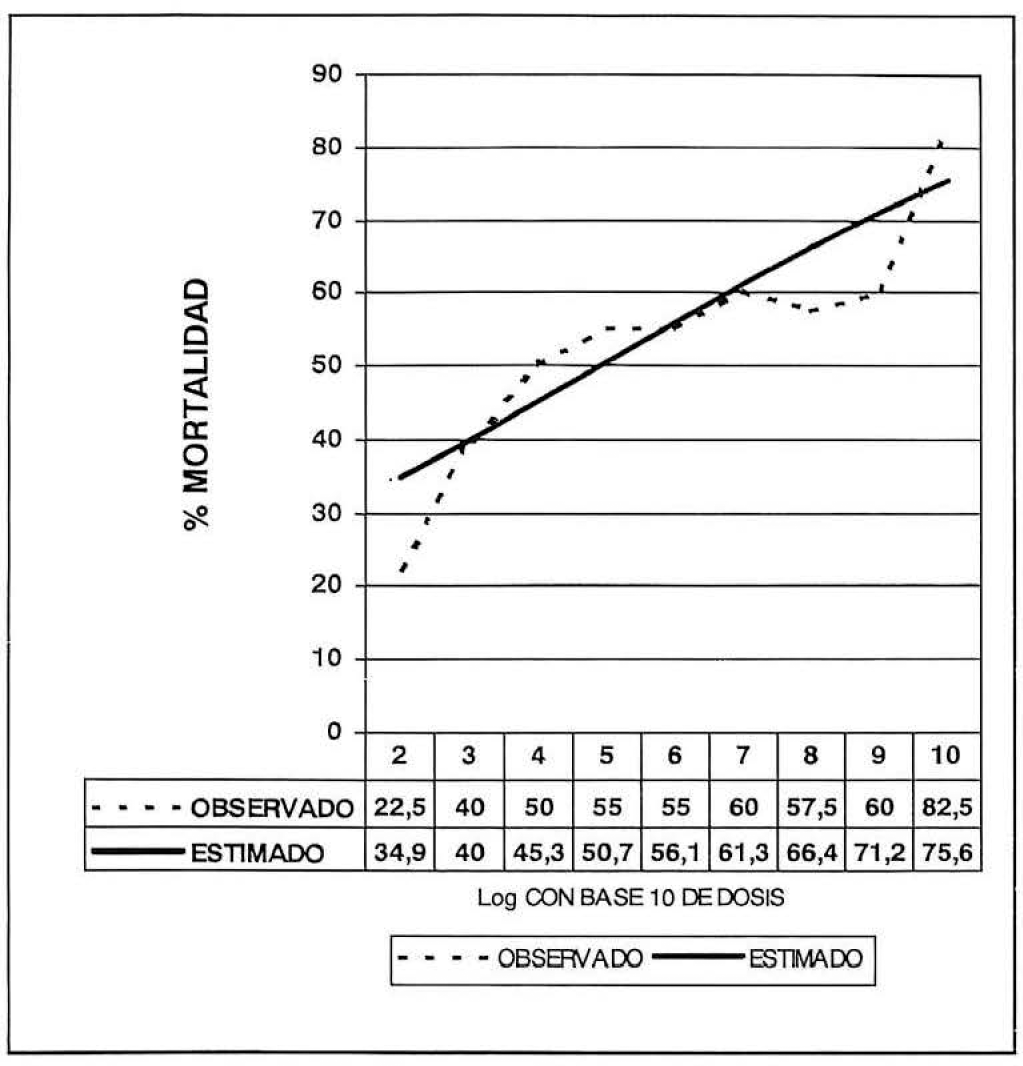
Porcentajes de mortalidad en diferentes concentraciones para determinar la dosis letal media de Beauveria bassiana (B.b. 9601) sobre Erinnyis ello.
Efecto de la aplicación en quinto ínstar sobre adultos y viabilidad de huevos.
Se presentó una mortalidad por B.b.9601 del 40% antes de que el insecto entrara al estado de pupa, mientras que el testigo mostró un 10% de mortalidad natural. El resto de la población (60%) alcanzó el estado de pupa y de adulto, los cuales no presentaron ningún tipo de deformación o manifestación del hongo evaluado. Las hembras emergidas ovipositaron normalmente; en cuanto a los huevos provenientes de estas presentaron una viabilidad del 97.81% y sólo el 2.19% no eclosionó, estas cifras sugieren un desarrollo normal del ciclo de E. ello. Cabe anotar que los huevos no eclosionados no presentaron manifestaciones de hongo. El porcentaje de eclosión en el testigo fue de 91.42%. Lo anterior indica que los efectos del hongo no se transmiten de una generación a otra.
Efecto de la aplicación de B.b.9601 sobre huevo.
De los huevos tratados con B.b.9601 el 79.5% eclosionaron, de estas larvas el 8.17% presentó mortalidad por causa del hongo. El 20.5% de huevos no eclosionados no presentaron signos del aislamiento. En el testigo eclosionaron un 84% de los huevos, de estas larvas el 1.7% presentaron mortalidad natural. La mortalidad por B.b.9601 en las larvas emergidas fue causada por el contacto con las conidias que se encontraban en la parte externa del corión y sobre la superficie de la hoja. Se puede concluir que aplicaciones de B.b.9601 sobre posturas de E. ello no presentan un efecto de mortalidad alto al compararse con los datos obtenidos en las aspersiones sobre larvas, lo que confirma lo registrado por Zacharuk et al citados por Roberts y Yendol (1971), quienes señalan que larvas de lepidópteros tienden a ser más susceptibles a hongos entomopatógenos que los huevos, pupas o adultos del mismo orden.
Efecto de la mezcla B.b.9601 y M.a.9206 sobre larvas de tercer ínstar.
Asperjar larvas de tercer ínstar con la mezcla de B.b.9601 y M.a.9206 causó una mortalidad del 90%, de la cual el 53.70% mostraron signos de B.b.9601 y el 46.3% de M.a.9206. En el testigo se presentó un 6.7% de mortalidad por causas naturales. Con estos resultados se deduce un efecto de sinergismo debido a que el porcentaje de mortalidad obtenido al inocularse esta mezcla superó a los datos de los experimentos anteriores. Aparentemente con la mezcla de los dos aislamientos se obtiene una mortalidad alta, por lo que se puede pensar en una implementación de este tipo de mezclas en un programa de manejo integrado de plagas.
Conclusiones
La recolección de insectos afectados por hongos entomopatógenos en cultivos de yuca fue baja ya que de cinco zonas evaluadas sólo en una se encontró esta clase de hongos. Se estandarizó una unidad experimental para evaluar hongos entomopatógenos sobre E. ello, implementando una larva inoculada con la suspensión conidial mediante aspersión larva - hoja de yuca e introduciéndola en una caja de cartón tipo cena #1 plastificada, disminuyendo la acción de otros patógenos y heridas causadas entre los insectos, metodología que se denominó "Unidad Experimental Múnera - De Los Ríos".
B.b.9601 causó la mayor mortalidad a E. ello. (31.6% a 87.5%) en condiciones de laboratorio, siendo el ínstar más susceptible el tercero y la dosificación letal media del aislamiento se obtuvo con 1x104.8018 conidias viables ml−1 y límites de confianza entre 1x100.3573 y 1x107.4120 conidias viables ml−1.
La acción del hongo no es transmitida de una generación a otra. Larvas emergidas de posturas de E. ello inoculadas con B.b.9601 no sufren un efecto considerable. La mezcla de B.b.9601 con M.a.9206 asperjada sobre larvas de tercer ínstar causó un porcentaje de mortalidad alto (90%) y no presentó antagonismo entre los aislamientos debido a que las larvas muertas manifestaron los signos típicos de ellos en forma individual.