
Abstract
Suspensions of conidia from the fungus Beauveria bassiana (Bals.) Vuill., Metarhizium anisopliae Sorok and Paecilomyces lilacinus (Deuteromycotina: Hyphomycetes) were evaluated in experiments with three media and two inoculation methods under laboratory conditions on adults of Cyrtomenus bergi. The insects of the control treatment were inoculated with sterilized distilled water plus tween 40 at 0.1%. Regarding the mortality, significant differences of Xβ=11.9; p≤0.01 and X2=6.39; p ≤ 0.05 were found between the species of fungus used and the inoculation method respectively. M. anisopliae was chosen as the most effective control agent and the most effective inoculation method corresponded to tarsal contact; the moist filter paper was chosen as the media for future experiments. Significant differences in mortality (p≤ 0.05) were found regarding the susceptibility between the nymphs and adults; nymphs were more susceptible than the adults to M. anisopliae. The fifth instar was chosen to evaluate thirteen isolates of M. anisopliae, of which the most promising were 9206, 9236 and 9501 with mortality of 62, 84 and 70% respectively. The isolate 9236 showed greater control, and the LD50 was determined as 1.19x108 viable conidia ml¹ with confidence limits between 5.91x103 and 3.36x109 and a LT50 between 4.7 and 8 days. Through another methodology in which sterilized soil was used as medium and the three best isolates in granulated formulation, it was detected that the isolate "9206" was superior to the isolates "9236" and "9501" with control levels of 63.9, 57.2, and 54.5% respectively. The isolate "9206" was the most effective within the habitat of the burrowing bug.
Introducción
Cyrtomenus bergi Froeschner (Hemiptera: Cydnidae) es un insecto que se encuentra ampliamente distribuido en América Latina, afectando diferentes cultivos. En Colombia se registró como plaga en el cultivo de yuca, a mediados de 1980, alimentándose de la raíz de la planta. El daño se caracteriza por la degradación de los tejidos alrededor de cada una de las lesiones causadas por el insecto (García 1982). C. bergi no ocasiona disminución del rendimiento; su daño reduce considerablemente el valor comercial del cultivo, por su efecto sobre la apariencia del producto y decrecimiento del contenido de almidón (Castaño et al.1985). En el CIAT se han realizado estudios básicos sobre este insecto tales como la biología y comportamiento (García 1982), fluctuación poblacional (Riis 1990), ensayos de control botánico con la leguminosa Crotalaria juncea (Castaño et al. 1985), de control químico (Vargas et al. 1986) y de control biológico del insecto con nemátodos parásitos de las familias Steinernematidae y Heterorhabditidae (Caicedo 1993; Barberena 1996). Hasta el momento estos controles no han sido implementados por no ajustarse a las exigencias sociales y ambientales. García (1982) encontró a la especie C. bergi parasitados por el hongo Metarhizium sp., en las colonias establecidas en el laboratorio. En Colombia, M. anisopliae se ha registrado parasitando 24 especies de insectos de 12 familias de los órdenes Coleoptera, Homoptera, Lepidoptera y Hemiptera (Hernández y Rodríguez 1992). Los hongos entomopatógenos se presentan como una alternativa de control natural de la chinche subterránea. Teniendo en cuenta que hasta el momento ningún tipo de estudio se había realizado para evaluar la patogenicidad de hongos entomopatógenos sobre este insecto, se propuso desarrollar una metodología para evaluar la patogenicidad de hongos entomopatógenos de la clase Hyphomycetes sobre C. bergi; determinar el estado biológico de la chinche más susceptible al patógeno; seleccionar entre aislamientos de M. anisopliae y determinar la DL50, en condiciones de laboratorio.
Materiales y Métodos
El trabajo se realizó en el laboratorio de Manejo Integrado de plagas en Yuca del CIAT Palmira, a una temperatura promedia de 23° C y 65% de HR. Los especímenes C. bergi se tomaron de la colonia del laboratorio, establecida en bandejas plásticas cuyo sustrato consta de suelo estéril al 20% de humedad. Se usaron doce aislamientos de M. anisoplie y uno de B. bassiana provenientes de diferentes hospederos y zonas colombianas, cedidos por CENICAFE. El aislamiento nativo de M. anisopliae fue obtenido directamente de C. bergi. El aislamiento de P. lilacinus fue cedido por CORPOICA Palmira (Tabla 1). Los aislamientos se multiplicaron en medio nutritivo artificial compuesto de Sabouraud destrosa agar + extracto de levadura al 0.1% SDAY; los cuales crecieron a temperatura ambiente por 20 días; una vez esporulados se cosecharon las conidias de cada aislamiento en 20 ml de agua destilada estéril + Tween 40 al 0.1% (ADET). Mediante pruebas de germinación se determinó la viabilidad de las conidias con la metodología propuesta por Dillon y Charnley (1990), los cuales definieron la conidia germinada como aquella que produce un tubo germinal igual o mayor al tamaño de la conidia, la cual fue adoptada para este ensayo.
Origen de aislamientos de Metarhizium aisopliae, Beauveria bassiana y Paecilomyces lilacinus.
Aislamientos cedidos gentilmente por CENICAFE.
Aislamiento cedido gentilmente por CORPOICA.
Determinación de metodología
Se utilizó la metodología propuesta por González et al. (1993), en un bioensayo para determinar patogenicidad de B. bassiana sobre Hypothenemus hampei, mediante la cual se evaluaron tres aislamientos de las especies M. anisopliae, B. bassiana y P. lilacinus sobre C. bergi. Los insectos fueron desinfestados con hipoclorito de sodio al 0.5% durante dos minutos y sometidos a "cuarentena" en cajas de petri, papel filtro húmedo con ADE y semillas de maní durante diez días. Se probaron tres sustratos constituidos por papel filtro humedecido con ADE, suelo esterilizado y no esterilizado traído de Santander de Quilichao, donde el insecto es considerado plaga en cultivos de yuca. El inóculo se utilizó en formulación líquida de conidias en ADET a una concentración de 1x108 conidias ml¹. Se utilizaron dos métodos de inoculación, uno de los cuales fue el de contacto tarsal el cual consistió en dejar que los insectos caminaran sobre una superficie inoculada por un tiempo de 2,5 minutos; la superficie estuvo representada por una caja de petri y el inóculo por 500 microlitros de la suspensión del hongo. Otro método consistió en aplicar el mismo volumen de inóculo en los sustratos antes mencionados. La combinación de las especies de hongos, métodos de inoculación y sustratos representaron los tratamientos, y en cada uno se evaluaron 10 especímenes individualizados en viales para un total de 200 especímenes. Los insectos del tratamiento testigo se inocularon con ADET en los sustratos papel filtro y suelo estéril. Las variables medidas después de la inoculación fueron día de muerte y esporulación del hongo sobre el insecto muerto.
Estado biológico más susceptible
La susceptibilidad de los seis estados biológicos de la chinche fue evaluada con el aislamiento de M. anisopliae nativo de C. bergi; los insectos se inocularon con suspensión de 1x10¹⁰ conidias viables ml¹, esta concentración se consiguió mediante centrifugación por 10 minutos a 6000 rpm (Moorhouse et al. 1992). De cada uno de los estados biológicos de la chinche se inocularon 20 insectos los cuales representaron una repetición; el testigo fue inoculado con ADET. Se realizaron cuatro repeticiones en el tiempo, para un total de 960 especímenes evaluados.
Selección entre aislamientos de M. anisopliae
El primer bioensayo se realizó sobre chinches de quinto instar, en el cual se evaluaron trece aislamientos colombianos de M. anisopliae sin reactivar, incluido el aislamiento nativo obtenido de C. bergi. Los 14 tratamientos estuvieron representados por los trece aislamientos del hongo y el tratamiento testigo; con cuatro repeticiones. En cada repetición se evaluaron 20 insectos para un total de 1120 especímenes.
Un segundo bioensayo de selección se realizó con los aislamientos de M. anisopliae 9206; 9236 y 9501 seleccionados en el bioensayo anterior, los cuales fueron masificados en arroz precocido en bolsas plásticas autoclavables (Londoño y Aterhortua 1995; Hernández y Rodríguez 1992). La producción de conidias en cada aislamiento se determinó por gramo de arroz esporulado. Para la selección entre aislamientos se utilizaron dos metodologías, una de las cuales correspondió a la del bioensayo anterior, bajo los mismos parámetros. La otra consistió en evaluar la formulación granulada del hongo en arroz, en cajas plásticas de 25x32x10 cm con tapa, en las cuales se depositó 1 Kg de suelo de Santander de Quilichao, tamizado, esterilizado con vapor de agua y secado al horno a 90°C por 72 horas. De cada aislamiento se pesaron 50 gramos de arroz esporulado, mezclado con el suelo. A cada bandeja se le adicionaron 300 ml de ADE quedando el suelo con una humedad aproximada del 30% la cual se corrigió semanalmente. En cada bandeja se colocaron 20 insectos de cada estado biológico de C. bergi previamente encuarentenados, para un total de 120 insectos por caja que representaron la población inicial. Los tratamientos estuvieron identificados por los aislamientos 9236; 9206; 9501 y el testigo con cinco repeticiones distribuidas en bloques completos al azar. A los 30 días se evaluó, contando los insectos vivos de III, IV, V instar y el estado adulto. Los resultados se analizaron mediante un análisis de varianza.
Dosis letal media
Se evaluó el efecto de doce concentraciones de conidias del aislamiento 9236 cuyo logaritmo en base 10 fue desde 1 hasta 11; y el tratamiento testigo se tomó como concentración cero. Cada concentración representó un tratamiento en el cual se evaluaron 20 chinches de V instar; con cuatro repeticiones para un total de 960 insectos de todos los tratamientos. Los datos obtenidos en las evaluaciones fueron sometidos a un análisis Probit para calcular DL 50
Resultados y Discusión
Todos los aislamientos presentaron porcentajes de germinación superiores al 92%, por lo cual se confirmó que el inóculo utilizado en el bioensayo fue viable y cumplió con los requisitos de calidad sugeridos por Bernal et al. (1994) donde el inóculo de M. anisopliae debe cumplir como mínimo 70% de germinación en medios nutritivos, a las 24 horas.
Determinación de la metodología
Se detectaron diferencias significativas en la variable "especies de hongo utilizadas" con respecto a la mortalidad (Xβ=11.9, gl=3, p < 0.01) entre los insectos tratados y el tratamiento testigo. La mortalidad total promedio de la población tratada con las diferentes cepas fue del 20%, distribuidas en cada especie de la siguiente manera: M. anisopliae 47.5%; B. bassiana 35%; P. lilacinus 12.5% y el tratamiento testigo 5%. No se detectaron diferencias estadísticas significativas con respecto a los diferentes sustratos usados; el análisis de la variable “método de inoculación" mostró diferencias significativas, con probabilidades de X2=6.39; gl=2; p < 0,05. La mortalidad total del 20% se distribuyó de la siguiente manera: Inoculación por contacto tarsal 12.5%; inoculación al sustrato 6.5%; contra el testigo que registró el 1%. En la figura 1 se observan manchas negras sobre el integumento de los insectos muertos coincidiendo con lo observado por Lacht (1976) en bioensayos con M. anisopliae sobre Oryctes rhinocerus; además de un insecto muerto con esporulación superficial verde que concuerda con observaciones de Alves (1986) y Hernández y Rodríguez (1992). Los insectos muertos por B. bassiana presentaron poca esporulación y los muertos por P. lilacinus no esporularon. Esto demuestra la especificidad de M. anisopliae con respecto a C. bergi bajo las condiciones anteriormente descritas, y la capacidad de formar epizotias como registraron Roberts y Humber (1983) en aislamientos que desarrollaron abundante esporulación sobre otros insectos.
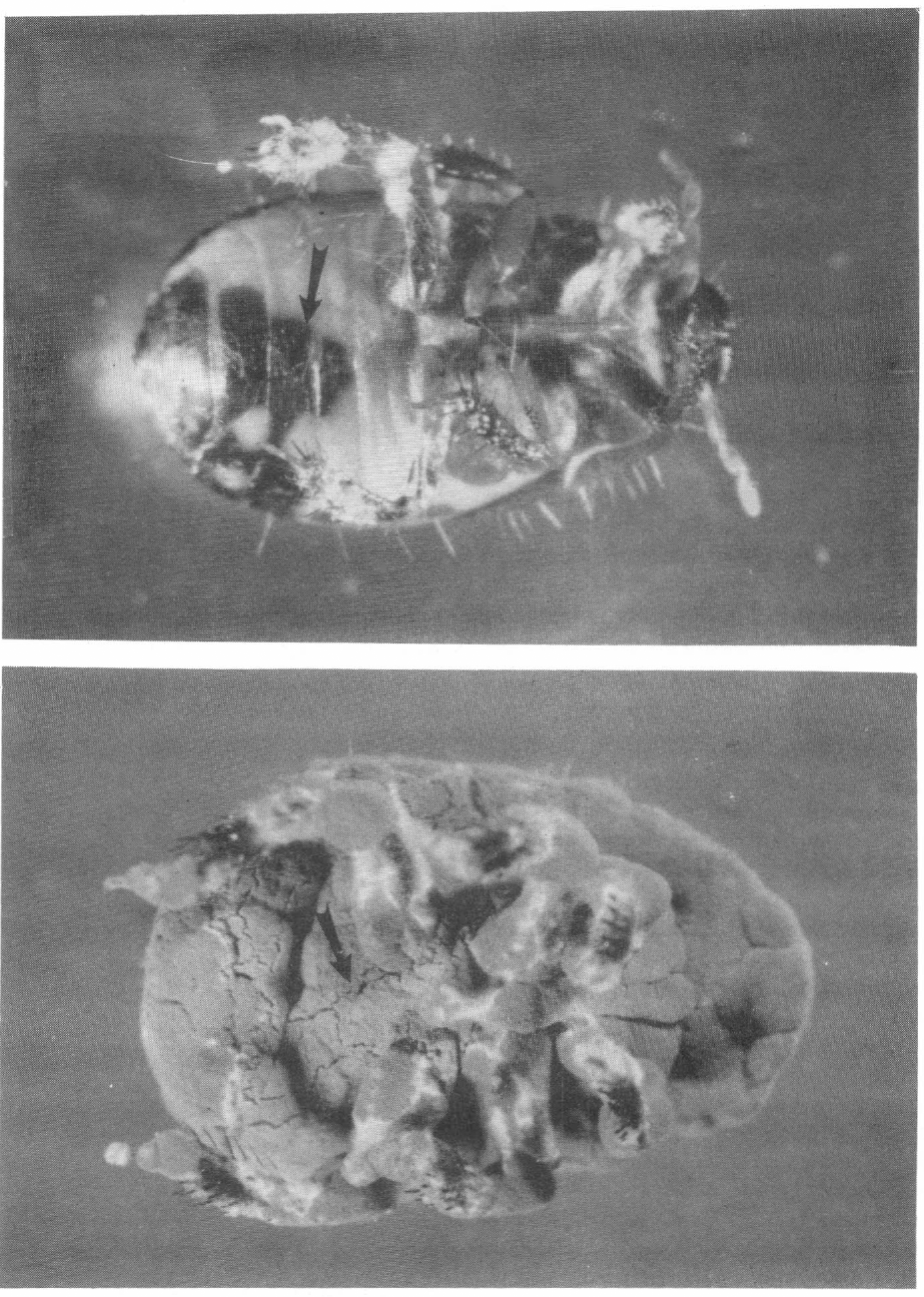
Cyrtomenus bergi F. afectado por M. anisopliae. Arriba sitio de penetración del hongo. Abajo insecto completamente esporulado. Fotografia Guillermo Guzmán, CIAT.
Estado biológico más susceptible
El análisis estadístico mostró significancia para la variable mortalidad de la chinche tratados con M. anisopliae y en todos los estados tratados y los no tratados. Los insectos muertos esporularon aproximadamente entre los 8 y los 12 días después de ser inoculados. Los resultados muestran dos grados definidos de susceptibilidad; el análisis no detectó diferencias significativas entre los estados inmaduros de la chinche, mientras que éstas fueron significativas entre la susceptibilidad del estado adulto (mortalidad del 62,5%) y los estados inmaduros (mortalidad del 86 al 92%) (Tabla 2). La mayor susceptibilidad de los estados inmaduros se debió probablemente a que éstos insectos no han alcanzado su máxima quitinización y los requerimientos energéticos del hongo para ejercer presión mecánica y acción enzimática sobre la cutícula, deben ser menores que los que necesita para penetrar la cutícula del insecto adulto; a diferencia de lo concluido por Vestergaard et al. (1995), quienes mencionan que el estado adulto es el más susceptible por la afinidad entre el hongo y la constitución química bien definida de la quitina. Por otra parte, los insectos tienen el menor número de hemocitos en su hemolinfa, al iniciar o terminar el instar. El sistema inmunológico del insecto se encuentra debilitado; las observaciones registradas por Patton y Flint (1959) citados por Mullet (1979) mencionan el fenómeno en Periplaneta americana que presenta reducción del número de hemocitos en los períodos de pre y post muda. Por lo anterior, la acción tóxica de M. anisopliae puede aumentarse considerablemente durante los períodos de pre y post muda en ninfas de C. bergi. Los resultados de esta investigación muestran que es posible cortar el ciclo del insecto, reduciendo las futuras poblaciones de la chinche. Los valores de mortalidad natural de C. bergi se encontraron dentro de los límites registrados por Riis (1990) y se ajustaron a los publicados por González et al (1993) y Vestergaard et al. (1995) sobre otros insectos (Tabla 2). Se eligieron ninfas de V instar para continuar la 2ª parte del estudio.
Porcentaje de mortalidad de los seis estados biológicos de C. bergi inoculados con M. anisopliae a 1 x 1010 conidias ml y no inoculados, evaluados en viales plásticos. Análisis hecho sobre la variable porcentaje de mortalidad transformado como Arc Sen Proporción de muertos.
Promedios seguidos por la misma letra no son estadísticamente diferentes según prueba de Ryan, Elliot, Gabriel, Welsch.
Selección entre aislamientos de M. anisopliae
En el primer bioensayo, se seleccionaron todos los aislamientos de M. anisopliae evaluados sobre C. bergi, por su patogenicidad contra el insecto determinado por mortalidades del 20 a 56.66%; a pesar de no encontrarse diferencias significativas al nivel de p≤ 0.05. Los criterios para seleccionar los mejores aislamientos se fundamentaron en los menores valores del coeficiente de variación y la media aritmética (≥ 40%) de las mortalidades obtenidas con cada aislamiento (Tabla 3). Es necesario realizar previamente la selección de los aislamientos patogénicos contra un insecto plaga dentro de un programa de control biológico como fue sugerido por Bernal et al. (1994) y otros autores. Los resultados de este estudio corroboran dicha sugerencia debido a que se confirmaron diferentes grados de susceptibilidad de la chinche a los aislamientos evaluados; el criterio de estabilidad de la patogenicidad de los aislamientos de M. anisopliae sobre un insecto hospedero nativo, se midió con el coeficiente de variación entre las repeticiones y fue importante para la selección de los aislamientos con patogénica semejante. A pesar de que existen registros sobre la alta efectividad patogénica de aislamientos de M. anisopliae sobre su insecto hospedero nativo (Farges y Robert 1976, 1978; Farges y Remaudière 1977 citados por Bernal et al. 1994), los resultados de este estudio al igual que los de Bernal et al. (1994) indican que no son tan específicos. La patogenicidad fue independiente del hospedero de origen y aparentemente el aislamiento de M. anisopliae nativo de C. bergi fue el mejor; sin embargo, con este hongo se obtuvo un mayor coeficiente de variabilidad entre repeticiones; posiblemente una de las causas de esta variabilidad está relacionada con la heterocariosis de la especie citada por Alves (1986) quien propuso que dicha recombinación genética influye sobre la patogenicidad de los diferentes aislamientos de M. anisopliae. Por los criterios de patogenicidad confirmada se seleccionaron los aislamientos 9206 y 9236; y el 9501.
Porcentaje promedio de mortalidad causada por trece aislamientos de M. anisopliae, a una concentración de 1 x 1010 conidias viales ml sobre C. bergi.
La concentración de conidias por gramo de arroz esporulado probada en el 2º bioensayo, fue de 5.94x109; 2.21x109 y 1.3×10¹º para los aislamientos 9206, 9236 y 9501 respectivamente, con porcentajes de germinación superiores al 92%. Estos resultados concuerdan con los obtenidos por Bernal (1992), citado por Posada (1993), donde se comprobó que la mayor producción de conidias de M. anisopliae se obtuvo en arroz (2.2x109 conidias gramo¯¹). Todos los aislamientos de M. anisopliae preseleccionados sobre C. bergi fueron patogénicos a la chinche, destacándose el aislamiento 9236 (origen desconocido) con el 84% de mortalidad, del chinche, comparado con los aislamientos 9501 (C. bergi) y 9206 (Aenolamia reducta) con los que se obtuvieron 70 y 62% de mortalidad, respectivamente. Lacht en 1976 encontró resultados similares con una cepa de M. anisopliae introducida, con la cual obtuvo mayor porcentaje de patogenicidad comparados con un aislamiento nativo de O. rhinocerus. La mortalidad de chinche obtenida en el tratamiento testigo no pasa del 10% (Fig. 2, Tabla 4). Con respecto a la mortalidad de C. bergi, obtenida con cada aislamiento, se observó un incremento en la virulencia de las mismas contra la chinche, debido probablemente a que los aislamientos del segundo bioensayo provenían de C. bergi, lo que se denomina "aislamiento reactivado" y por consiguiente al ser evaluados de nuevo, expresaron su máximo potencial patogénico (Londoño y Atehortúa 1995).
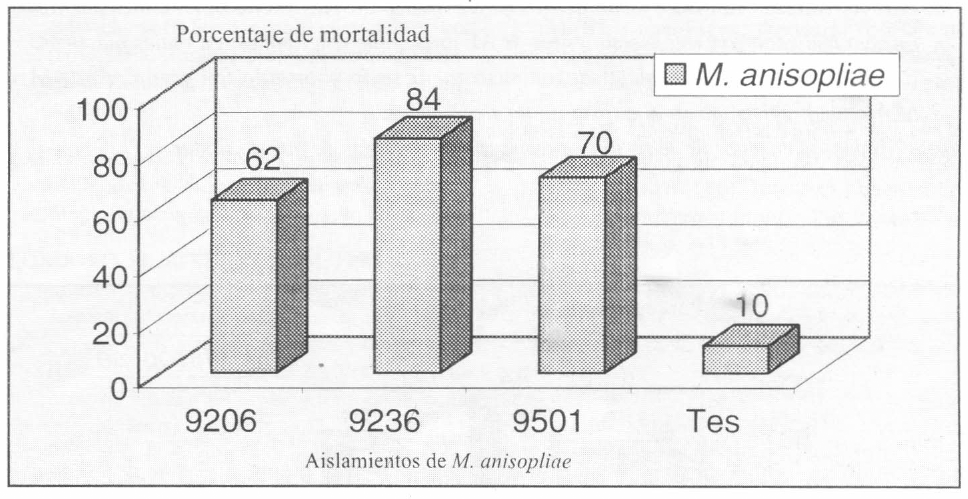
Porcentajes de mortalidad causadas por tres aislamientos promisorios de M. anisopliae sobre de C. bergi evaluado en viales plásticos.
Porcentajes de mortalidad causadas por tres aislamientos de M. anisopliae seleccionados como promisorios sobre C. bergi evaluados en viales plásticos. Análisis hecho sobre la variable porcentaje de mortalidad transformado como Arc Sen Proporción de muertos.
Promedios seguidos por la misma letra no son estadísticamente diferentes según prueba de Ryan, Elliot, Gabriel, Welsch.
En el análisis de varianza del porcentaje de mortalidad de hongos entomopatógenos, por tratamiento del suelo con inóculo desarrollado en arroz, se identificaron tres grupos, de acuerdo con el valor de la media; se destaca el aislamiento 9206 con 63.9% de mortalidad. El tratamiento testigo mostró diferencias significativas con respecto a los aislamientos 9236 y 9206, pero no con respecto al aislamiento 9501 nativo de C. bergi. Así mismo, se encontraron diferencias entre los aislamientos 9501 y 9206; por el contrario, entre los aislamientos 9236 y 9206 no se detectaron diferencias estadísticas significativas (Fig. 3, Tabla 5). La formulación granulada de los tres aislamientos causaron mortalidad en C. bergi, bajo condiciones favorables al insecto, en lo que respecta a los requerimientos de humedad registrados por Riis (1990); la temperatura, bajo la cual se trabajó, favoreció a ambos organismos (chinche y hongo) y corresponde a las temperaturas registradas en el Municipio de La Tebaida (Quindío) y en el corregimiento de la Bella (Risaralda) donde el insecto es problema en diferentes cultivos. Los porcentajes de mortalidad obtenidos con los mejores aislamientos en todos los bioensayos, muestran que la utilización de suelo contaminado con los patógenos desarrollados en granos de arroz, en cajas plásticas, es promisoria, porque simula las condiciones donde C. bergi es plaga en cultivos de yuca y porque permitió seleccionar el aislamiento 9206 como superior al 9236 seleccionado en papel filtro húmedo (Tabla 6).
Porcentajes de mortalidad causadas por tres aislamientos promisorios de M. anisopliae, evaluados con suelo en cajas plásticas y formulación granulada sobre C. bergi. Análisis hecho sobre la variable porcentaje de mortalidad transformado como Arc Sen √Proporción de muertos.
Promedios seguidos por la misma letra no son estadísticamente diferentes según prueba de Ryan, Elliot, Gabriel, Welsch.
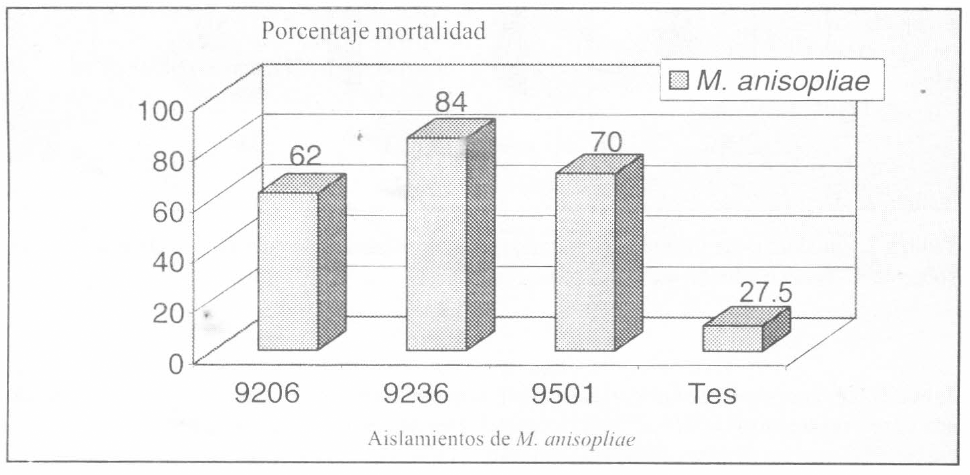
Porcentajes de mortalidad de C. bergi causada por tres aislamientos de M. anisopliae en formulación granulada, evaluada en cajas plásticas con sustrato de suelo.
Dosis letal media
La dosis letal media es obtenida con M. anisopliae a 1x108.075 conidias viables/ml con un límite de confianza de 1x103.7716 y 1x109.5268. entendiéndose estos valores como la concentración de conidias viables necesarias para causar el 50% de mortalidad a C. bergi. El tiempo que necesitó el hongo para causar la DL50 estuvo en un rango estimado de 4.7 a 8 días. Se comprobó que la mortalidad de la chinche está directamente relacionada con el incremento en la dosis del inóculo: con 1x10¹ conidias/ml se obtuvo 12.5% de mortalidad mientras que con 1x105 esporas/ml la mortalidad aumenta a 85% (Fig. 4). Estos mismos conceptos los han registrado Butt et al. (1994) y otros autores.
Comparación del comportamiento de tres aislamientos de M. anisopliae seleccionados como promisorios para controlar C. bergi en condiciones de laboratorio.
Ensayo realizado con trece aislamientos de M. anisopliae sin reactivar, en viales plásticos.
Ensayo realizado con tres aislamientos de M. anisopliae reactivados, en viales plásticos.
Ensayo realizado en cajas plásticas con sustrato de suelo y formulación granulada de M. anisopliae.
Mortalidad registrada en el quinto instar de C. bergi.
Nota: Todos los ensayos de selección se realizaron con ninfas de quinto instar.
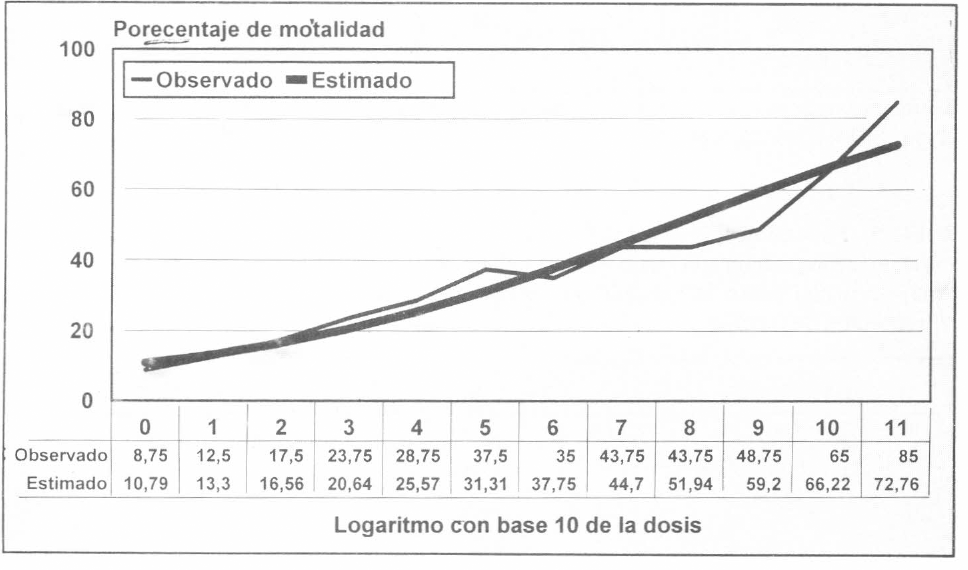
Porcentaje de mortalidad causada por el aislamiento 9236 de M. anisopliae sobre C. bergi, evaluado con diferentes concentraciones de conidias en viales plásticos.
Conclusiones
Se determinó que M. anisopliae causó mayor mortalidad a C. bergi que B. bassiana y P. lilacinus. La unidad experimental más adecuada para la evaluación de hongos Hyphomycetes sobre C. bergi. fue el insecto inoculado con una suspensión de conidias contaminado por contacto tarsal y depositado en viales de plástico con papel de filtro húmedo, a temperaturas de 23°C.
Los signos de M. anisopliae desarrollados sobre C. bergi fueron los típicos de la muscardina verde; empezando por pérdida de movilidad, manchas negras en integumento de los estados inmaduros, inicio de crecimiento de micelio blanco (antenas, aparato bucal y tarsos), cubrimiento de micelio en toda la superficie del insecto, finalizando con la esporulación de color verde y dispersión de conidias.
La cuarentena es importante tenerla en cuenta en el desarrollo de bioensayos para evaluar aislamientos de M. anisopliae sobre C. bergi; porque éste es un hongo de ocurrencia natural. Si no es tenida en cuenta puede incrementar el registro real de la patogenicidad del aislamiento evaluado.
Los estados inmaduros de C. bergi fueron los más susceptible a M. anisopliae en comparación con el estado adulto en O. rhinocerus se registró menor mortalidad. Los aislamientos de M. anisopliae más eficientes para el control de C. bergi en condiciones de laboratorio fueron: el 9236 (origen desconocido), el 9206 (Aenolamia reducta) y el 9501 (C. bergi). Se determinó una relación directa entre la dosis y el porcentaje de mortalidad, y la dosis letal media se obtuvo con 1.19x108.075 conidias viables/ml, con límites de confianza entre 5.91x10³ y 3.36x109; y el tiempo que necesitó el hongo para causar dicha mortalidad en un rango estimado de 4.7 a 8 días.