
Abstract
Thrips palmi Karny is an important pest of several crops, especially vegetables. The predatory mite Neoseiulus cucumeris has been successfully used in the control of various thrips species. The predatory mite T. aripo also has shown considerable capacity to consume cassava thrips. The potential impact of N. cucumeris and T. aripo on populations of T. palmi was evaluated. The objectives were, to determine the effects of consumption of T. palmi on the biology of N. cucumeris and T. aripo, and to compare to mite prey species Tetranychus urticae and Mononychellus caribbeanae; to quantify the consumption of the predators on T. palmi and evaluate the functional response of N. cucumeris and T. aripo to densities of 1, 3, 5, 10, 20 y 40 of first instar larvae of T. palmi. The studies were carried out under laboratory conditions (25 ± 5°C, 75 ± 5% of r. h. and 12:12 L:D). The development period of N. cucumeris when feeding on T. palmi and T. urticae was 8.9 and 8.5 days respectively. For T. aripo it was 15.1 days with T. palmi and 13.5 days with M. caribbeanae. Fecundity for N. cucumeris and T. aripo was 25.2 and 15.6 respectively, being higher on T. palmi than mite prey. The percentage of females in the population and longevity of predatory mites were similar with both prey species offered. N. cucumeris was the predator with the higher fecundity and longevity. An individual of N. cucumeris consumed 65.3 first instar larvae of T. palmi and 18.7 second instar larvae during its life cycle. T. aripo consumed 72.9 first instar larvae of T. palmi and 21.6 second instar larvae during its life cycle. N. cucumeris and T. aripo females displayed a type II functional response, consuming 16 and 18 first instar T. palmi in 24 hours, responding positively to higher densities of thrips.
Introducción
Thrips palmi Karny es nativo de la región que abarca Malasia e Indonesia y se encuentra en varios países de Asia, África, Oceanía, las Américas y el Caribe (Hall et al. 1993; Johnson 1986; Seal 1997; Vergara 1999a). Su presencia en Colombia se detectó en 1997 en varios cultivos de Antioquia y el Valle del Cauca (Vergara 1999a; Durán et al. 1999) y actualmente se distribuye por todo el país (Vergara 1999b).
T. palmi ataca más de 50 especies de plantas, dentro de las cuales están incluidas las solanáceas, cucurbitáceas y leguminosas (Wang y Chu 1986, citado por Hall et al. 1993). En Colombia, se encuentra en más de 30 hospederos, afectando en algunos cultivos hasta el 100% del área sembrada (Durán et al. 1999; Vergara 1999b).
Según Cermeli y Montagne (1993), el daño es causado por ninfas y adultos al raspar los tejidos para chupar la savia. Ataca preferiblemente las hojas, pero puede causar daños a flores y frutos. En las hojas, el daño comienza generalmente por las nervaduras y luego se propaga a toda la superficie dándole un aspecto de tostado que les causa la muerte. En los frutos, sobre todo en berenjena y en pimentón, los puntos de alimentación se convierten en cicatrices y deformaciones dependiendo del estado de crecimiento al momento del ataque. Las mayores poblaciones y daños se han observado en la estación seca.
En el manejo de Thrips palmi en Colombia se acude principalmente a un control químico irracional, pues en todas las zonas se usan productos que no tienen registro contra la plaga (Vergara 1999b). Sin embargo, se han adelantado experiencias en otras áreas de control (Alvarez et al. 2000; CIAT 2000; Vasco y Guarín 1999). Estudios presentados por Durán y Mesa (1999) indican que se puede lograr un buen control al integrar el uso de Biomel con liberaciones semanales de Chrysoperla externa y Guarín y Parra (1999) lograron prescindir de la aplicación de insecticidas de síntesis al hacer liberaciones de C. externa (Hagen) (Neuroptera: Chrysopidae). No obstante, T. palmi por ser una plaga introducida, no cuenta con una presión alta de controladores biológicos que en condiciones naturales regulen sus poblaciones adecuadamente.
La lista de parasitoides, predadores y patógenos que controlan a T. palmi en varios países del mundo es amplia (Etienne et al. 1990; Gillespie 1989; Hirose et al. 1993; Hirose et al. 1999; Seal 1997). Dentro de los predadores asociados en condiciones naturales con T. palmi están los ácaros de la familia Phytoseiidae de los géneros Amblyseius, Euseius, Fundiseius, Iphiseiodes, Neoseiulus, Phytoseius y Proprioseiopsis (Duran et al. 1999; Etienne et al. 1990; Hirose et al. 1993; Kreiter y De Moraes 1997). Sin embargo, el impacto de éstos en la regulación de las poblaciones de trips no está bien definido.
Neoseiulus (= Amblyseius) cucumeris (Fig. 1 A) es el ácaro predador más utilizado en el control de especies de trips como Frankliniella occidentalis y Thrips tabaci (Gillespie 1989; Steiner 1990; Hoy y Glenister 1991; Shipp y Whitfield 1991; Wittman y Leather 1997). Se ha empezado a usar con éxito para el control de T. palmi (Castineiras et al. 1997; Wada 1999). Typhlodromalus aripo (Fig. 1 B), otro ácaro predador encontrado en yuca y ocasionalmente en fríjol, se encuentra predando en las colonias sobre T. palmi. De otra parte, tiene una excelente capacidad de consumir Scirtothips manihoti, una especie de trips asociada a yuca (CIAT 1998). Aunque la yuca es un hospedero accidental de T. palmi, en el Laboratorio de Acarología del CIAT, se ha observado a los trips consumir y desarrollarse, desde larva hasta adulto, sobre hojas de yuca, lo que constituye un peligro potencial para este cultivo (CIAT 1999).
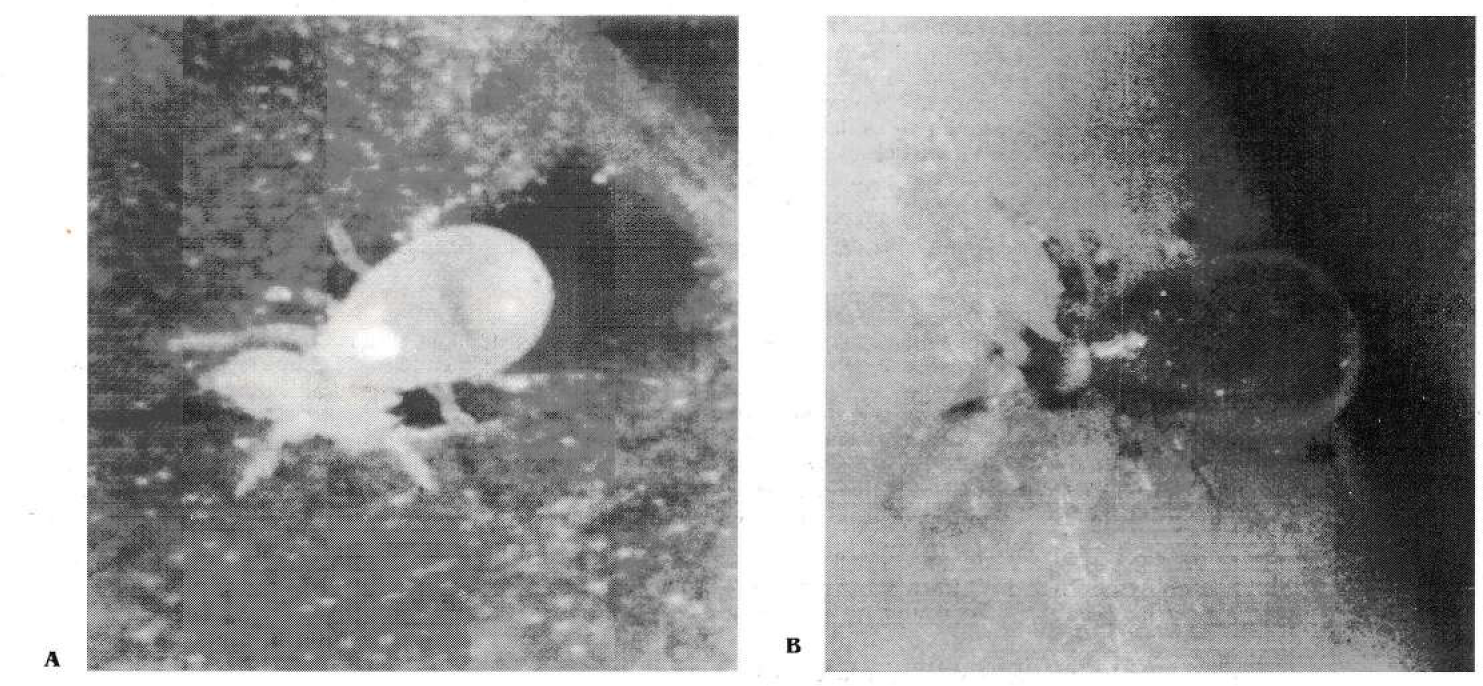
Hembras de Neoseiulus cucumeris (A) y Typhlodromalus aripo (B) consumiendo una larva de primer ínstar de Thrips palmi (fotografía de Rodrigo Zúñiga).
El uso potencial de un predador se puede medir en apectos de su biología y capacidad de consumo sobre una presa determinada. Igualmente, con estudios de respuesta funcional, la cual describe la relación entre la densidad de presa y el número de presas atacadas por un predador por unidad de tiempo (Eveleigh y Chant 1981; Sabelis 1985; Solomon (1949), citado por Sabelis 1985). Estos aspectos aún no han sido estudiados con predadores fitoseidos teniendo a T. palmi como presa.
Con el fin de evaluar a N. cucumeris y T. aripo como controladores potenciales de T. palmi, los objetivos de este estudio fueron: determinar el efecto del consumo de T. palmi sobre el desarrollo, fecundidad y longevidad de N. cucumeris y T. aripo, en comparación con las presas acarinas Tetranychus urticae y Mononychellus caribbeanae; cuantificar el consumo de los predadores sobre T. palmi, y evaluar a N. cucumeris y T. aripo en su respuesta funcional a diferentes densidades de T. palmi.
Materiales y Métodos
Presas ofrecidas
A los predadores se ofrecieron individuos de T. palmi, los cuales se colectaron en plantas de fríjol en el municipio de Pradera (Valle) y se establecieron en una colonia en el laboratorio a 25 ± 2°C, 75 ± 5% de HR y 12:12 (L:O), sobre plantas de fríjol de la variedad ICA-Pijao. Como presas comparativas se proporcionaron los ácaros fitófagos Tetranychus urticae Koch para N. cucumeris y Mononychellus caribbeanae (McGregor) para T. aripo. Las colonias de los tetraníquidos se establecieron en plantas de yuca de dos meses de edad de la variedad CMC-40 en casas de malla del CIAT.
Ácaros predadores
N. cucumeris fue obtenido de IPM Laboratories, Inc. Locke, Nueva York y la colonia se estableció con el método McMurtry y Scriven (1965), donde se ofrece a los predadores huevos de T. urticae, polen de higuerilla (Ricinus communis) y miel de abejas. La otra especie de fitoseido, Typhlodromalus aripo, procedente de Cruz das Almas (Brasil), se estableció usando el método de cría de Mesa y Bellotti (1987), en el cual se ofrecen hojas de yuca infestadas con la presa M. caribbeanae. Estas colonias y todos los estudios se realizaron en el laboratorio a 25 ± 5°C, 75 ± 5 de HR y 12:12 (L:O).
Estudios de biología y consumo
Las unidades experimentales consistieron en frascos plásticos de 2 cm de diámetro por 1 cm de alto, dentro de los cuales se colocó un disco de papel filtro humedecido, y sobre éste un disco de hoja limpia de yuca. En cada una de estas unidades se individualizaron huevos de una misma cohorte de cada uno de los predadores y se adicionaron cinco larvas de primer ínstar de T. palmi. Los frascos se taparon con plástico vinipel. Se registró el cambio de estado de los fitoseidos y la cantidad de presa consumida fue contada y restablecida cada día.
Del mismo modo, se colocaron huevos de una misma cohorte de N. cucumeris y de T. aripo en frascos que contenían hojas de yuca infestadas con abundante cantidad de estados mezclados de los ácaros T. urticae y M. caribbeanae, respectivamente. El cambio de estado de los fitoseidos se registró diariamente. La cantidad de presa acarina ofrecida siempre excedió los requerimientos de los predadores en este estudio al igual que en los de fecundidad y longevidad.
Para los estudios de fecundidad se depositaron en las unidades ya descritas cinco larvas de primer ínstar y cinco de segundo instar de T. palmi, y en cada una de éstas se individualizaron hembras de los predadores de un día de emergencia y cópula. Diariamente se registró la oviposición, el consumo y se restableció la cantidad de los trips hasta la muerte de las hembras. La fecundidad sobre las presas acarinas se realizó en frascos con hojas infestadas con T. urticae para N. cucumeris y M. caribbeanae para T. aripo.
La utilización de presas acarinas de alta aceptación por parte de los fitoseidos (Castagnoli y Simoni 1991; CIAT 1998), permite comparar los resultados de desarrollo y fecundidad con los obtenidos con T. palmi y determinar si el trips es una presa importante para estos predadores.
Los parámetros biológicos evaluados se analizaron estadísticamente con el software Statview (Abacus concept, USA). Antes de hacer los ANOVAS respectivos, se examinó la homogeneidad de la varianza y la normalidad de los datos por la prueba de F y el método Kolmogorov-Smirnov. Solamente el parámetro tiempo de desarrollo no mostró una distribución normal de los datos, por lo tanto, se utilizó la prueba de U. de Mann Whitney para comparar los promedios. La prueba Chi2 se utilizó para comparar los porcentajes de sobrevivencia y de hembras. Para comparar los promedios de fecundidad, longevidad y consumo se utilizó la prueba de Fisher PLSD (Protected Least Significant Difference), siguiendo la ANOVA.
Respuesta funcional de Neoseiulus cucumeris y T. aripo
Se utilizaron hembras de los predadores de tres días de emergencia y después de haber iniciado su periodo reproductivo. Las densidades ofrecidas fueron 1, 3, 5, 10, 20 y 40 larvas de primer ínstar del trips, y se sometieron al consumo durante 24 horas. Se realizaron 15 repeticiones por cada densidad.
Se mencionan en la literatura tres tipos principales de respuesta funcional que describen el consumo de un predador por su presa: el tipo I, una línea recta que se eleva hasta alcanzar una meseta; el tipo II, una curva que se eleva hasta alcanzar una meseta; y el tipo III, una curva sigmoide que se eleva hasta alcanzar una meseta (Sabelis 1985). Los modelos de respuesta funcional tipo I y tipo II se tomaron de Holling (1959) y el modelo de tipo III fue el presentado por Hull et al. (1977).
Los modelos fueron:
Tipo I y = an + b
Tipo II y = aTn/(1 + aThn)
Tipo III y = A/(1 + Exp (B - Cn))
Donde: y es el número de presas consumidas; a, tasa de ataque; n, densidad inicial de presa; b, intercepto con el eje y; T, tiempo de exposición a la presa; Th, tiempo de manipulación y A, B, C parámetros estimados.
La estimación de los parámetros para el tipo I se realizó por cuadrados mínimos mediante una regresión lineal. La estimación de los parámetros para los tipos II y III se realizó usando el algoritmo Marquardt por cuadrados mínimos no lineales (SAS Institute 1979). El criterio para la evaluación de los modelos fue el valor de r2.
Resultados y Discusión
Efecto del consumo de T. palmi sobre el desarrollo, fecundidad y longevidad de N. cucumeris y T. aripo en comparación con presas acarinas
El tiempo de desarrollo de N. cucumeris cuando se alimentó de T. palmi fue de 8.9 dias, y de 8.5 cuando se alimentó de T. urticae (Tabla 1). Para T. aripo el tiempo de desarrollo con T. palmi fue de 7.7 días, significativamente mayor que con M. caribbeanae que fue de 6.5 días. Para N. cucumeris los dos tipos de presa tienen el mismo efecto en la duración del ciclo de vida (no se presentaron diferencias significativas), y aunque para T. aripo es mayor con el trips, no puede afirmarse que su ciclo se prolongue considerablemente con esta presa. Adicionalmente, una de las ventajas de los predadores Phytoseiidae es la de tener un ciclo de vida menor que el de sus presas (Mesa y Bellotti 1987), que en el caso de T. palmi está entre los 11 y los 15 días de acuerdo con el hospedero (Bueno y Cardona, 2001; Duran et al. 1999).
El porcentaje de sobrevivencia de huevo a adulto en general fue alto con los dos tipos de presa. Sin embargo, fue significativamente mayor con las presas acarinas con respecto de T. palmi. Así, N. cucumeris presentó un 94.5% de sobrevivencia con T. urticae y 81.2% con T. palmi. T. aripo tuvo una sobrevivencia de 98.8% con M. caribbeanae y de 89.8% con T. palmi. Entre especies de predadores no se presentaron diferencias significativas al ofrecerse presas acarinas o el trips (Tabla 1).
Para ambos predadores las presas ofrecidas favorecieron la producción de hembras. De este modo, el porcentaje de hembras para N. cucumeris fue de 65.2% y de 53.7% con T. palmi y T. urticae, respectivamente. Para T. aripo fue de 81% y 71% con T. palmi y M. caribbeanae, respectivamente. Para este parámetro evaluado, ni la especie de predador ni el tipo de presa influyeron significativamente en la cantidad de hembras de la población (Tabla 1).
Efecto de dos tipos de presa sobre el tiempo de desarrollo (promedio ±DS), sobrevivencia de inmaduros y porcentaje de hembras de N. cucumeris y T. aripo
Promedios seguidos por la misma letra, para cada especie de predador, no difieren significativamente al nivel del 5% (Prueba de U. de Mann Whitney).
Promedios seguidos por la misma letra, para ambas especies de predadores, no difieren significativamente al nivel del 5% (Prueba de Chi2). a,b: comparación por tipo de presa, u: comparación por especie de predador.
La fecundidad de N. cucumeris cuando se alimentó de T. palmi fue de 25.2 huevos, significativamente mayor que cuando se alimentó de T. urticae que fue de 18.6 huevos (Tabla 2). La fecundidad para T. aripo con T. palmi y M. caribbeanae fue de 15.6 y 11.8 huevos respectivamente, siendo significativamente mayor con el trips como presa.
En cuanto a la longevidad de los predadores no se presentaron diferencias significativas con ninguna de las dos presas. Para N. cucumeris fue de 48.8 y 36.4 días con T. palmi y T. urticae, respectivamente (Tabla 2). Para T. aripo la longevidad fue de 24.6 días con T. palmi y de 20 días con M. caribbeanae.
Fecundidad (promedio ± DS) y longevidad (promedio ± DS) de N. cucumeris y T. aripo con dos tipos de presa
Promedios seguidos por la misma letra para cada especie de predador no difieren significativamente al nivel del 5% (PLSD de Fisher).
El tipo de presa (A) influye de manera significativa en la fecundidad de los predadores (p< 0.05), (Tabla 3). En efecto, cuando se usa el trips como presa se observa un aumento significativo de la fecundidad de N. cucumeris y T. aripo. Considerando el factor Especie de predador (B), la fecundidad es superior cuando el predador es N. cucumeris independiente del tipo de presa ofrecida (p < 0.0001). En la interacción tipo de presa por especie de predador (A × B) se obtuvo un valor de p superior de 0.05, indicando que esta interacción no influye sobre la fecundidad (Tabla 3).
Resultados del ANOVA-2 cuantificando el efecto del tipo de presa y la especie de predador sobre la fecundidad y la longevidad de hembras de fitoseidos
Aunque no hay influencia del tipo de presa sobre la longevidad de los predadores (Tabla 3), se observa que los individuos viven más tiempo cuando se alimentan de trips (Tabla 2). La especie de predador si influye fuertemente en este parámetro (p < 0.0001), siendo mayor la longevidad de N. cucumeris que la de T. aripo. No hay efecto en la interacción A × B sobre la longevidad (Tabla 3).
Como ya se mencionó, las presas acarinas utilizadas son de alta preferencia por los predadores (Castagnoli y Simoni 1991; CIAT 1998). Los resultados aqui obtenidos indican que con T. palmi la reproducción y la longevidad de los predadores es mayor o igual que con las presas acarinas. Por esto, se sugiere que N. cucumeris y T. aripo son predadores potenciales que podrían controlar a T. palmi.
Capacidad de consumo de N. cucumeris y T. aripo sobre Thrips palmi
En la Figura 2 se presenta el consumo diario de hembras de los predadores sobre larvas de T. palmi. El mayor consumo de trips se registró desde los dos días de iniciado el periodo reproductivo hasta el día 13, tiempo que coincide de acuerdo con el periodo de oviposición de las hembras. Los picos de consumo máximo se registraron entre los días tres a cinco, predando un promedio de seis trips por día por T. aripo y cinco por N. cucumeris. A partir del día 16 para ambas especies el consumo fue inferior a dos presas por día, lo cual sólo contribuye al mantenimiento de las hembras hasta su muerte. Así, las hembras son potencialmente importantes para controlar T. palmi durante los 13 primeros días de edad.
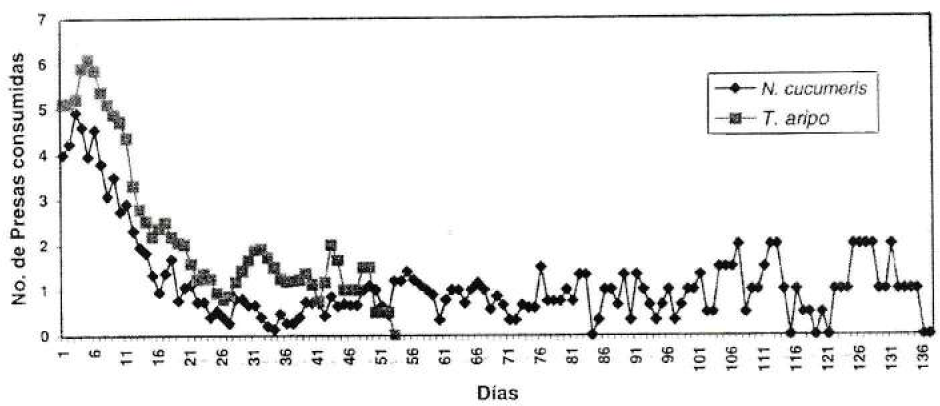
Consumo diario de hembras de N. cucumeris y T. aripo sobre larvas de T. palmi.
Se observó que cuando los predadores incrementan la talla, capturan más fácilmente las larvas del trips. De este modo, los estados inmaduros predan principalmente larvas de primer ínstar. A diferencia, las hembras de los fitoseidos, evadiendo el comportamiento defensivo de los trips, capturan con menos dificultad las larvas de segundo ínstar. No obstante, prefieren agotar las del primer ínstar. Se observó que cuando las larvas de trips son "tocadas" por un predador, éstas levantan agresivamente el abdomen. Y como ocurre usualmente, si son alcanzadas por la parte trasera, baten su abdomen arrojando lejos a su atacante. Este fenómeno ya ha sido descrito para otros tisanópteros, especialmente en los machos, hecho que está involucrado en la competencia por las hembras, pero también se presenta en la defensa al ataque de los predadores (Bakker y Sabelis 1989).
Las larvas de N. cucumeris no se alimentaron, confirmando lo observado por Shipp y Whitfield (1991), como ocurre con otras especies de fitoseidos (Sabelis 1985). Así, el consumo sobre larvas de primer ínstar de T. palmi por inmaduros de N. cucumeris, desde protoninfa hasta adulto, fue de 10 larvas y por hembras fue de 55.3 larvas (Tabla 4). El consumo desde larva hasta adulto de T. aripo fue de 8.25 y por hembras fue de 64.7 larvas de primer instar de T. palmi. Para ambas especies de predadores se presentaron diferencias significativas en el consumo entre inmaduros y hembras (p < 0.05). De otra parte, el consumo por hembras de los predadores sobre larvas de segundo ínstar de T. palmi fue de 18.7 por N. cucumeris y de 21.6 por T. aripo.
Los resultados de ANOVA-2 indican que el estadío del predador para las dos especies influye considerablemente en la capacidad de consumo, siendo las hembras más voraces que los estados inmaduros (p < 0.0001) (Tabla 4). La especie de predador no tiene una influencia marcada en el consumo (p < 0.0406). En contraste, el factor Estadío de la presa si tiene un efecto fuerte, presentándose un consumo mayor sobre larvas de primer ínstar que sobre larvas de segundo instar de T. palmi (p < 0.0001) independiente de la especie del predador.
Efecto del estadío de desarrollo de T. palmi sobre el consumo (promedio DS) de inmaduros y hembras de N. cucumeris y T. aripo y resultados de ANOVA a dos factores
Promedios seguidos por la misma letra para cada especie de predador no difieren significativamente al nivel del 5% (PLSD de Fisher). a, b: comparación entre columnas; u, v: comparación entre filas.
Respuesta funcional de N. cucumeris y T. aripo sobre larvas de primer ínstar de T. palmi: comparación de modelos
Modelos 1 y 2, Holling (1959); Modelo 3, Hull et al. (1977). Para los modelos II y III, los parámetros fueron estimados usando el algoritmo Marquardt por cuadrados mínimos no lineales (SAS Institute, 1979).
Totalizando el consumo se tiene que un individuo de N. cucumeris consume 65.3 larvas de primer instar de T. palmi y 18.7 larvas de segundo ínstar durante su vida, y T. aripo consume 72.9 larvas de primer ínstar y 21.6 de segundo ínstar durante su vida.
Respuesta funcional de N. cucumeris y T. aripo sobre larvas de primer ínstar de T. palmi
Las ecuaciones de regresión representando los tres modelos se encuentran en la tabla 5. Según los valores del coeficiente de correlación r2, el valor más alto para las dos especies de predadores corresponde al tipo II del modelo de Holling (1959). Por lo tanto, es el que permite ajustar y representar mejor los datos. Así, la respuesta funcional de tipo II presentada por N. cucumeris y T. aripo (Fig. 3) se caracteriza por un aumento en el número de presas atacadas a un nivel decreciente, a medida que aumenta la densidad de presa. No obstante, aunque el máximo consumo observado para N. cucumeris fue de 16.1 y de 18 larvas para T. aripo a la densidad 40, las curvas no alcanzaron claramente la meseta al rango de densidades probadas. Las curvas descritas por ambos predadores son similares en consumo, dada la similitud de los valores de a (tasa de ataque) y Th (tiempo de manipulación) calculados.
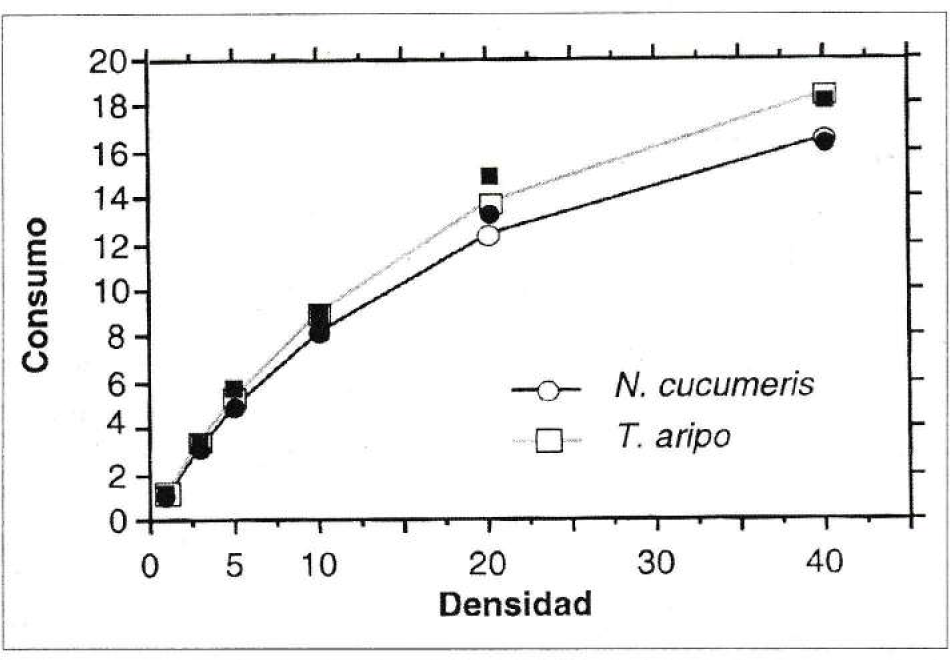
Respuesta funcional de hembras de N. cucumeris y T. aripo sobre larvas de primer ínstar de T. palmi. a = tasa de ataque; Th = tiempo de manipulación. Para N. cucumeris, a = 0.052; Th = 0.96. Para T. aripo, a = 0.056, Th 0.85. Las curvas muestran los valores esperados según la ecuación del disco de Holling (1959). Los símbolos en negrita representan los valores promedios observados de consumo. N. cucumeris(●); T. aripo (■).
El tipo II de respuesta funcional es el que comúnmente describen los ácaros Phytoseiidae (Eveleigh y Chant 1981; Sabelis 1985), y fue el presentado para N. cucumeris cuando la presa fue Frankliniella occidentalis (Shipp y Whitfield 1991). Sin embargo, el máximo valor de consumo por el predador fue de 10 larvas de primer ínstar de trips en 24 horas, menor al encontrado en este trabajo con T. palmi como presa.
Con el presente estudio se mejora el entendimiento sobre la dinámica de las relaciones entre estos predadores y el trips como presa. Adicionalmente, se puede afirmar que N. cucumeris y T. aripo responden funcionalmente a densidades altas de T. palmi. En sentido práctico, estos modelos permiten estimar el número de hembras del predador a liberar con una frecuencia determinada y a cierta densidad de población de la plaga.
N. cucumeris es un predador ampliamente distribuido (Castagnoli y Simoni 1991), presente en una gran variedad de hospederos como pimentón, pepino, y berenjena entre otros (Shipp y Whitfield 1991; Wada 1999), mostrando una gran capacidad de adaptación en los hábitats de T. palmi. De otra parte, T. aripo es una especie que puede ser eficiente en fríjol. Los hábitats donde se encuentra T. palmi son muy agresivos para los enemigos naturales debido a la cantidad de aplicaciones de insecticidas que se realizan contra ésta y otras plagas (Vergara 1999a). Sin embargo, los trabajos de Durán y Mesa (1999) indican que es posible integrar el control químico con el control biológico de manera eficiente. Con la presente investigación, se inicia el desarrollo de una nueva estrategia de control biológico para T. palmi en Colombia, que puede unirse al esfuerzo de establecer un programa de manejo de esta plaga ecológicamente más acorde con los cambiantes agroecosistemas actuales.
Conclusiones
El tiempo de desarrollo para N. cucumeris es igual con los dos tipos de presa. En contraste, para T. aripo fue mayor con la presa T. palmi. La sobrevivencia de los predadores durante el desarrollo fue alta con los dos tipos de presa, pero superior con las presas acarinas. El porcentaje de hembras estuvo sesgado a favor de las hembras y el tipo de presa no afectó este parámetro.
La fecundidad fue mayor con T. palmi para ambos predadores, y la longevidad fue igual con los dos tipos de presa. N. cucumeris es la especie con mayor capacidad reproductiva y mayor longevidad.
Se presenta una mayor capacidad de consumo por hembras que por los estados inmaduros de los predadores, y existe una preferencia marcada por larvas de primer ínstar que por las de segundo ínstar. Un individuo de N. cucumeris consume 65.3 larvas de primer ínstar y 18.7 larvas de segundo ínstar de T. palmi durante su vida, y T. aripo consume 72.9 larvas de primer ínstar y 21.6 de segundo ínstar durante su vida.
Las hembras de N. cucumeris y T. aripo presentaron una respuesta funcional de tipo II con T. palmi, respondiendo positivamente a densidades altas del trips. Así, las hembras de N. cucumeris y T. aripo consumen 16 y 18 larvas de primer ínstar de T. palmi en 24 horas, respectivamente.
Footnotes
Agradecimientos
A las siguientes personas del Programa de Manejo Integrado de Plagas y Enfermedades del CIAT: de modo especial a Paul Calatayud por su permanente asesoría, análisis estadístico y revisión del manuscrito. Rodrígo Zúñiga por su colaboración en la realización de esta investigación. Josefina Martínez por su ayuda en la presentación. Guillermo Sotelo del Programa de Entomología de Forrajes y Myriam Cristina Duque del Programa de Arroz y Biodiversidad del CIAT, por la asesoría y el análisis de los parámetros de respuesta funcional en SAS. Jorge Gallego de la Unidad de Artes Gráficas por su colaboración en la realización de las gráficas.