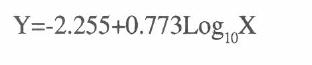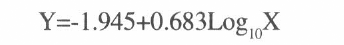The subterranean burrowing bug, Cyrtomenus bergi is a pest that damages cassava roots. Its feeding mode facilitates the introduction of root pathogens, deteriorating root quality. The present research, undertaken at the International Center for Tropical Agriculture (ICTA), initiated the development of methodology to mass rear the nematode Heterorhabditis bacteriophora, measured the parasitic capacity of two strains LFR92 and SQC92 on C. bergi, determined the most susceptible development stage; distinguished parasitic comparison between strains, determined if C. bergi responds differentially to several doses of n/ml, and the DL50 for each strain. There were no significant differences in the parasitic capacity between the strains LFR92 and SQC92 of H. bacteriophora. The most susceptible stage of C. bergi to H. bacteriophora parasitism was the fifth nymphal stage for both strains. C. bergi showed a differential response to several doses of n/ml. The DL50 for SQC92 was determined as 826 n/ml with limits [545:1137], and for LFR92 as 720 n/ml, with limits [81:1551]. The strain LFR92 proved to be the more aggressive of the two strains. H. bacteriophora is a potential control agent for C. bergi since it has presented high levels of parasitism and because it's a native strain.
El cultivo de la yuca es de gran importancia para los países tropicales ya que es una fuente de carbohidratos con bajo costo de producción y crece en suelos poco fértiles, permitiendo el uso de tierras marginales (Riis 1990).
El cultivo es atacado por un complejo de plagas, dentro de las cuales se encuentra Cyrtomenus bergi Froeschner registrada hacia 1981, la cual, por su modo de alimentación, causa un daño constante en la raíz deteriorando la calidad del producto, y por su hábito subterráneo, puede pasar desapercibido durante todo el período vegetativo de la planta, provocando que al momento de la cosecha se obtengan pérdidas considerables aún con bajos niveles de infestación (García y Bellotti 1980). Igualmente ha sido señalada en países como Ecuador, Brasil, Cuba y Panamá. En Colombia también ataca otros cultivos como cebolla, espárrago, maíz, maní y café (Riis 1990).
Se han intentado varios métodos de control, como el uso de agroquímicos, con los cuales no se ha logrado evitar el daño, aumentándose los costos y deteriorándose el medio ambiente. También se ha ensayado el control cultural con cultivo intercalado con Crotalaria juncea donde el daño se ha disminuido pero el rendimiento ha caído hasta un 22% (García y Bellotti 1980). Se hizo necesario, entonces, buscar un método alternativo de control que no deteriorara el ambiente, como es el caso de los enemigos naturales, dentro de los cuales se encuentra el nemátodo Heterorhabditis bacteriophora Poinar (Rhabditida: Heterorhabditidae).
Los nemátodos entomopatógenos tienen ventajas frente a otros tipos de control biológico pues tienen un amplio rango de hospederos y gran crecimiento en cultivos artificiales (Klein 1990). Estos han sido utilizados como agentes de control de varios insectos plaga como Popillia japonica (Selvan et al. 1993), Cyclocephala borealis (Klein 1990), Cylas formicarus (Mannion y Janson 1992), Ceratitis capitata (Lindegren et al. 1990), Rhizotrogus majalis, Diaprepes abbreviatus, Hylobius sp. (Klein 1990).
H. bacteriophora tiene una asociación mutualística con la bacteria Protorhabdus luminiscens (Poinar 1990) quien causa septicemia en el insecto 48 horas después de haber penetrado (Hom 1994). Los nemátodos poseen un rango de aplicación en el campo similar al de la bacteria Bacillus popilliae, y se propone aplicarlos por irrigación, avioneta, inyecciones en el suelo (Georgis 1990), también se propone liberar cadáveres infectados de Galleria mellonella (Jansson et al 1993). Una característica importante es que las poblaciones de artrópodos depredadores del suelo no se ven afectados negativamente por estos nemátodos entomopatógenos (Georgis et al.1991).
Por lo anterior se planteó la presente investigación con tres hipótesis de trabajo: 1. Conocer si existen diferencias en cuanto a la capacidad parasítica entre las razas LFR92 y SQC92 del nemátodo H. bacteriophora sobre la chinche C. bergi (H1). 2. Conocer si existe un estado de desarrollo del insecto más susceptible al ataque del nemátodo (H2). 3. Establecer si C. bergi presenta una respuesta diferencial a varias dosis de nemátodos por mililitro (n/ml) (H3) y determinar la DL50 para cada una de las razas del nemátodo.
Materiales y Métodos
Esta investigación se realizó en el laboratorio de Entomología de Yuca, en el Centro Internacional de Agricultura Tropical CIAT, Palmira (Valle), con una temperatura de 23°C y una humedad relativa de 65%, en promedio.
Se trabajó con dos razas del nemátodo nativo H. bacteriophora, previamente aisladas y renovadas durante la realización del presente estudio, y denominadas LFR92 (La Florida, Risaralda) y SQC92 (Santander de Quilichao, Cauca). Los individuos de C. bergi se obtuvieron de la colonia previamente establecida en laboratorio.
Con las razas del nemátodo se desarrolló un método de cría en laboratorio que consistió en el uso de dos metodologías modificadas. Para la cría in vitro se siguió el método propuesto por Bedding (1981, 1984), y para la cría in vivo se siguió aquel propuesto por Woodring y Kaya (1988).
Con los nemátodos así obtenidos se montaron los ensayos para la medición de la capacidad parasítica. En éstos, las razas fueron evaluadas separadamente y los especímenes de C. bergi individualmente. Los insectos se ubicaron en recipientes cónicos de 5x3 cm, con un grano de maíz pregerminado, 5 g de suelo franco-arcilloso y 1 ml de la solución de nemátodos de concentración deseada para un contenido de humedad del 45% P/P. Al testigo se le adicionó agua destilada estéril. La evaluación se realizó 10 días después por disección al estereoscopio.
De esta forma, se montó un ensayo preliminar, en el cual se enfrentaron las dos razas del nemátodo a todos los estados de desarrollo del insecto, es decir, se tuvo un tamaño de muestra de 40 individuos de C. bergi/estado de desarrollo del insecto/raza del nemátodo, con una concentración de 8000 n/ml; la población testigo consistió de 40 individuos testigo de C. bergi/estado de desarrollo del insecto y se realizó una repetición. A partir de este ensayo, se determinó si las razas presentaban un parasitismo diferencial, cuál estado de desarrollo del insecto era más susceptible y cuáles eran las concentraciones de n\ml para determinar la DL50.
Los ensayos para establecer la DL50 para cada una de las razas del nemátodo se montaron de la forma anteriormente descrita. Para la raza LFR92 se utilizaron las concentraciones 200, 500, 1000, 2000, 3000, 4000 y 6000 n/ml. Para la raza SQC92 se utilizaron estas mismas concentraciones y además, 10000 n/ml. El tamaño de la muestra fue de 10 individuos de 5o. estado de desarrollo de C. bergi/concentración de n/ml/raza del nemátodo/repetición; la población testigo consistió de 10 individuos de 5o. estado de desarrollo del insecto/repetición; se realizaron 4 repeticiones. Con los resultados obtenidos se determinó si el insecto presentaba una respuesta diferencial a las concentraciones de n/ml y la DL50 para cada raza.
Los datos de todos los ensayos se analizaron con el paquete estadístico SAS (Sistema de Análisis Estadístico). Para probar la H1 se realizó una prueba X2 de independencia entre las razas de H. bacteriophora para la variable "parasitado", con modalidades si y no, y un 95% de significancia. Para la H2 se agruparon los datos en Grupo 1 (Ninfa 1 a Ninfa 4) y Grupo 2 (Ninfa 5 y Adulto) y se realizó un X2 de independencia para la variable "sano" con modalidades si y no, y un 95% de significancia. Para la H3, se tomaron las razas por separado y se realizó un Xβ de independencia para las variables "parasitado" y "sano", con modalidades si y no, bajo la hipótesis nula de que el número de individuos sanos, viniendo de diferentes concentraciones, no varía con la dosis y se utilizó un 95% de significancia. Para determinar la DL50 los datos se sometieron a análisis Probit.
Resultados y Discusión
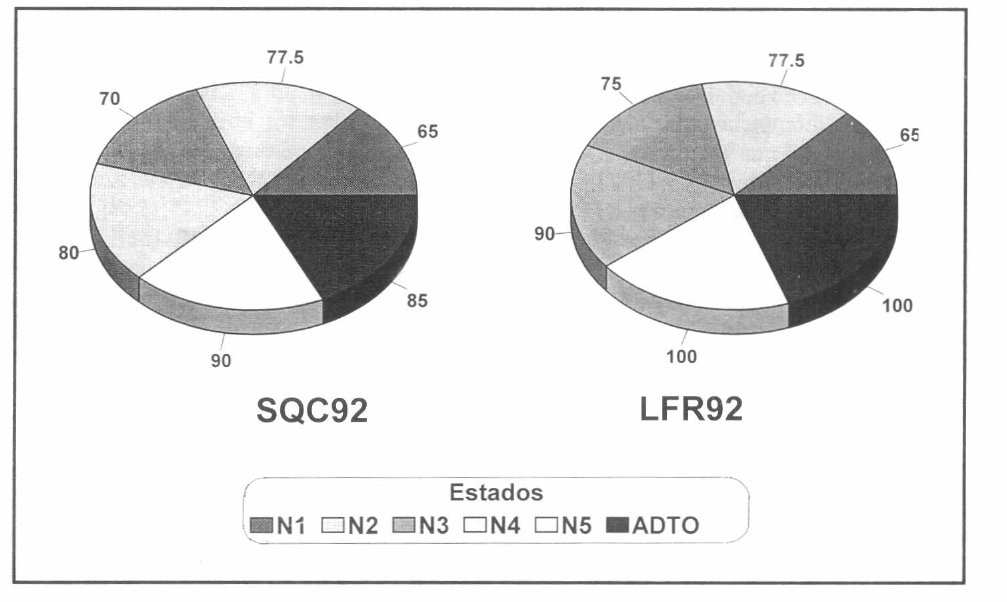
En el ensayo preliminar se vio que H. bacteriophora es capaz de parasitar todos los estados de desarrollo de C. bergi (Fig. 1). Se obtuvo un parasitismo global para SQC92 de 77.92% y para LFR92 de 84.58%. Al tomar los estados de desarrollo de forma independiente el menor parasitismo se obtuvo con ninfas de primer estado, quienes presentaron con ambas razas 65% de parasitismo; mientras que los mayores parasitismos se presentaron sobre ninfa 5 y adulto, con 90% y 85%, respectivamente, para SQC92, y 100% en ambos estados, para LFR92. Se puede decir que los individuos de ninfa 5 y adulto son más susceptibles al ataque de los nemátodos ya que son de mayor tamaño que las ninfas de primer estado; también sus aperturas naturales son mayores facilitando la penetración de los infectivos, sin embargo, para corroborar estas posibilidades es necesario desarrollar un estudio sobre el particular.
Las ninfas de segundo, tercer y cuarto estado de desarrollo presentaron niveles de parasitismo intermedios entre ninfa 1 y ninfa 5. Las ninfas de tercer estado mostraron un parasitismo menor que las de segundo y cuarto estado. Con la raza LFR92, el parasitismo en estos tres estados fue superior a los obtenidos con la raza SQC92.
Esta capacidad de parasitar todos los estados de desarrollo de C. bergi permite potencializar el nemátodo como un posible agente de control del insecto, que está causando un daño constante al cultivo pues su forma de alimentación facilita la entrada de fitopatógenos a través de la herida dejada por el estilete.
En la prueba X2 (Xβ=3.50; p=0.061) se obtuvo que las razas no presentan una capacidad parasítica significativamente diferente; aunque al observar los estados de desarrollo, de forma individual, se encontró que el parasitismo fue diferencial sobre ninfas de 5 estado (p=0.04) y adulto (p=0.011).
Parasitismo (%) por estado de desarrollo de C. bergi, de las razas SQC92 y LFR92 de H. bacteriophora, con una dosificación de 8000 n/ml, en laboratorio.
Las diferencias en cuanto a parasitismo no sólo dependen de las características de las razas/especies de nemátodos, sino también de la susceptibilidad del hospedero, así las diferencias aquí obtenidas serían el resultado de la interacción raza del nemátodo/insecto.
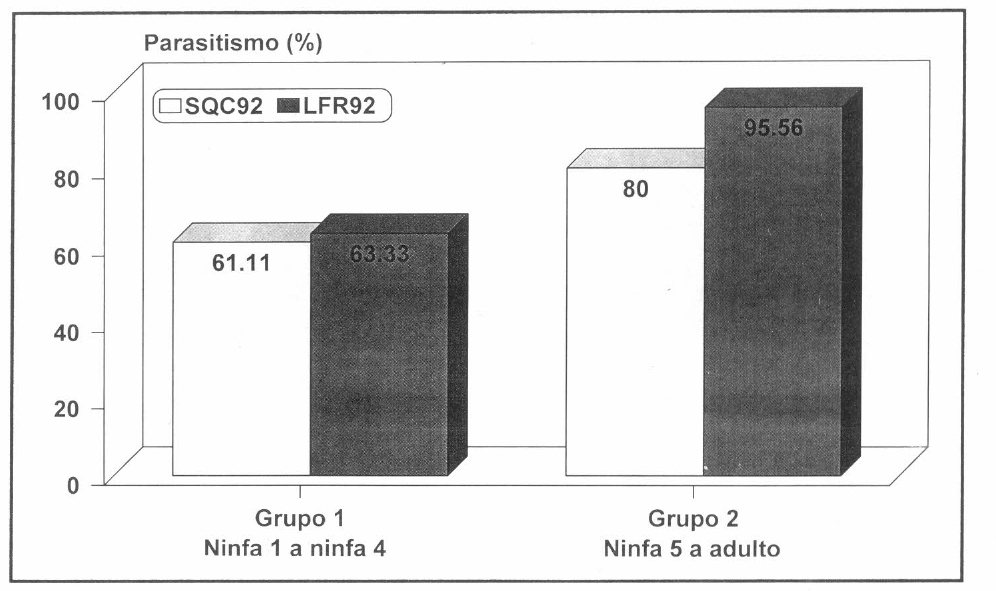
En el agrupamiento realizado para determinar susceptibilidad, se encontró que en el grupo 1 (N1 a N4) se presentó un parasitismo de 61% con SQC92 y de 63% con LFR92, y en el grupo 2 (N5 y Adulto) se vio un parasitismo de 80% con SQC92 mientras que con LFR92 fué del 95% (Fig. 2). En la prueba estadística (95% de significancia) se determinó que existen diferencias en el parasitismo entre el grupo 1 y el grupo 2 (Xβ=6.323; p=0.012), es decir, entre individuos pequeños e individuos grandes; por lo tanto, se puede plantear que los nemátodos de las razas LFR92 y SQC92 tienen una marcada preferencia por parasitar los individuos de tamaño mayor (Fig. 2).
Parasitismo (%) de SQC92 y LFR92 de H. bacteriophora, por grupo ninfal de C. bergi, con una dosificación de 8000 n/ml después de diez días de inoculado.
Se puede observar que entre los individuos del grupo 2, las ninfas de 5o. estado presentan porcentajes de mortalidad mayores que los individuos adultos (Fig. 3); lo que indica que los individuos del 5o. estado de desarrollo son más susceptibles al ataque de los nemátodos de H. bacteriophora. El quinto estado de desarrollo reúne características tales como tamaño adecuado para producir progenie de nemátodos, diámetro de sus aberturas naturales correcto para permitir el paso de los estados infectivos; además, el abdomen es voluminoso por tener gran contenido de líquido y por consiguiente la cutícula es más suave. Diezmar la población de C. bergi sobre ninfas de 5º estado de desarrollo, presenta varias ventajas, ya que son individuos que tienen una alta sobrevivencia (mortalidad natural sobre maíz es del 2%, Riis 1990) es decir, que casi en su totalidad alcanzan el estado adulto, el cual es el de mayor longevidad. De esta forma, se cortaría el ciclo de vida, ya que no habría oviposición, lo que se traduciría en una disminución de individuos en poblaciones futuras. Por lo tanto, la obtención de la DL50 para cada raza se realizó con individuos de este estado de desarrollo.
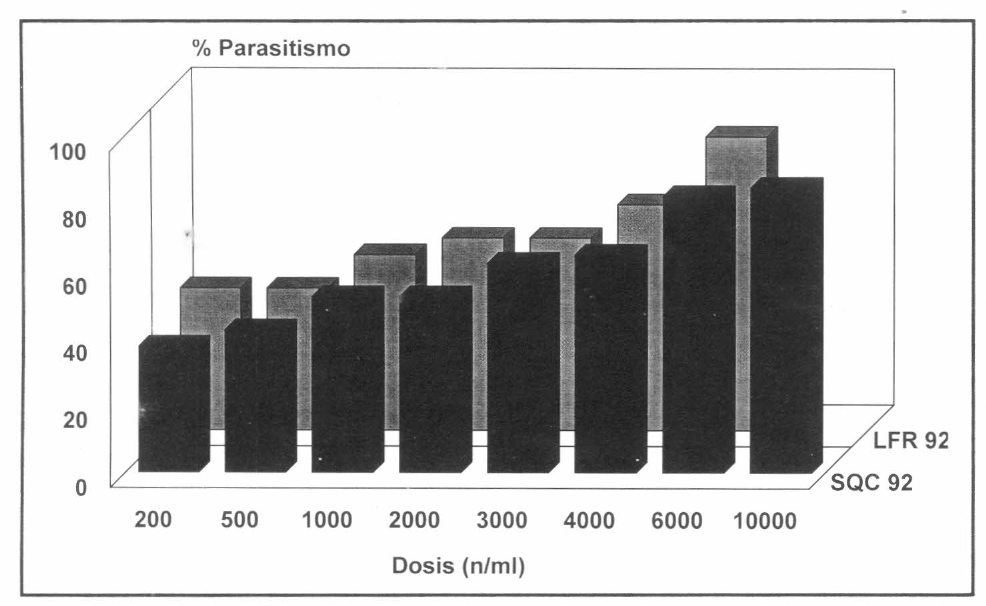
En la determinación de si existe o no respuesta diferencial a varias dosis de n/ml, se encontró que para LFR92 el parasitismo varió de 42.5% a 87.5% mientras que para SQC92 de 37.5% a 85% (Fig. 4). En la prueba estadística (al 95% de significancia) se apreció, que el número de individuos sanos varió con la dosis, disminuyendo a medida que la dosis aumentaba (SQC92: X2=84.978, p=0.00; LFR92: X2=71.012, p=0.00).
Mortalidad (%) por estado de desarrollo de C. bergi, SQC92 y LFR92 de H. bacteriophora, con una dosificación de 8000 n/ml y evaluado a los diez días.
Para la determinación de la DL50 se utilizaron los datos de parasitismo, aunque el modelo Probit trabaja con mortalidades, ya que dichos datos son más significativos pues un insecto parasitado vivo sólo requiere tiempo para alcanzar la muerte y, además, al momento de la evaluación no todos los insectos parasitados habían muerto. Debido a esto, los datos se sometieron a una prueba de bondad de ajuste con respecto al modelo Probit (Tabla 1), con lo que se estableció que las DL50 y sus límites de confianza debían ser calculados utilizando factores de corrección, que para SQC92 fue de 1.96 y para LFR92 de 2.57.
En el análisis Probit (Tabla 2) se obtuvo que la ecuación de la recta que describe el comportamiento de SQC92 es
donde X es la concentración utilizada y Y el valor de probit. Así, se obtuvo una DL50 de 826 n/ml, con límites [545,1137]. Para la raza LFR92 la ecuación es
y la DL50 es 702 n/ml, con límites [81,1551].
Prueba de bondad de ajuste al modelo Probit, del parasitismo presentado por las razas SQC92 y LFR92 de H. bacteriophora
Raza
Estadístico
Valor
GL
Probabilidad
Fact Correc.
SQC92
X2
9.66
6
0.1398
1.96
LFR92
X2
13.96
5
0.0159
2.57
Procedimiento Probit de las razas SQC92 y LFR92 de H. bacteriophora
Raza
Variable
GL
Estimado
Error Stand.
X2
Probab.
SQC92
INTERCEPTO
1
-2.255
0.323
48.63
0.0001
Log10 DOSIS
1
0.773
0.099
61.39
0.0001
LFR92
INTERCEPTO
1
-1.945
0.598
10.58
0.0011
Log10 DOSIS
1
0.683
0.188
13.21
0.0003
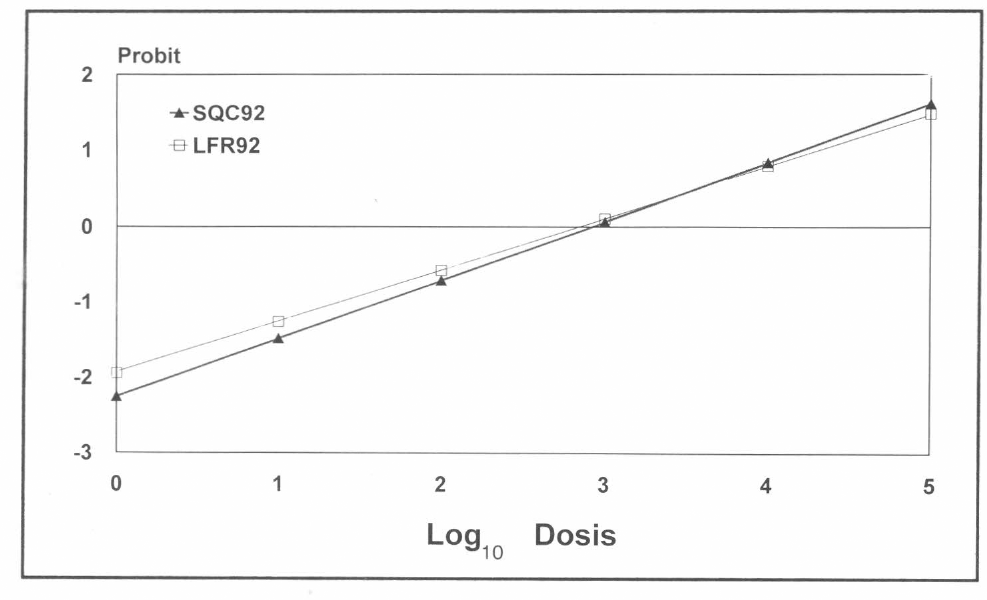
En la figura 5 se observa que la recta de SQC92 tiene un crecimiento más rápido que la de LFR92; bajo el eje X se puede interpretar que con LFR92 se requieren dosis menores para lograr los mismos resultados que si se utiliza SQC92; al alcanzar un valor X=4.00, es decir, una concentración de 10000 n/ml se puede ver que la raza SQC92 es la que tiene mayor impacto, pues, con las mismas dosis, arroja porcentajes mayores de parasitismo.
Parasitismo de SQC92 y LFR92 de H. bacteriophora, sobre el 5º estado de desarrollo de C. bergi, con ocho dosificaciones diferentes, a los diez días.
En ambos casos, los límites de confianza de las DL50 obtenidas son muy amplios, es decir, que con la raza SQC92, al utilizar tanto 545 n/ml como 1137 n/ml, se puede obtener un 50%, lo mismo que con LFR92. Esto revela un desajuste con respecto al modelo probit. Epsky y Capinera (1994) plantean corregirlo evaluando la eficiencia de invasión de los nemátodos sobre el insecto hospedante y no el parasitismo.
Al comparar estos límites de confianza, entre las razas, se observa que los de LFR92 son más amplios que los de SQC92, es decir, que esta última tiene un comportamiento que se ajusta mejor al descrito por el modelo Probit, lo cual le permite tener un comportamiento más predecible que LFR92. Así, si se busca proyectar estos resultados a campo; con SQC92, los datos obtenidos van a ser más similares a los esperados que si se utiliza LFR92; al mismo tiempo fue con esta raza con la que se alcanzaron porcentajes de parasitismo de 100% con una dosis de 8000 n/ml, mientras que con la raza SQC92 no se superó el 90% de parasitismo, aún con dosis tan altas como 10000 n/ml. Por consiguiente, la raza LFR92 es más agresiva y por lo tanto más deseable para el control de C. bergi pero su comportamiento es más impredecible que el de SQC92, que, a pesar de necesitar dosis mayores para alcanzar niveles similares de parasitismo, presenta mayor predicción en su comportamiento.
Caicedo y Bellotti (1994) registraron para Steinernema carpocapsae Weiser (Rhabditida: Steinernematidae) una DL50 de 193 n/ml, sobre C. bergi; ésta es baja al compararla con las obtenidas con H. bacteriophora, pero los autores no señalaron mortalidades al momento de la evaluación; este aspecto se convertiría en una ventaja de las razas de H. bacteriophora ya que interrumpen el daño que los individuos del insecto están causando a las raíces de la planta; además, estas razas son nativas y por lo tanto están adaptadas a las condiciones ambientales donde se encuentra C. bergi; por otra parte Bedding (1981) menciona que las especies de Heterorhabditis son más virulentas que las de Steinernema.
El mismo autor y sus colaboradores (Bedding et al. 1983) mencionan que una asociación prolongada de un nemátodo con un insecto lleva a que la población parásita busque hacer el menor daño posible a su población hospedera, por lo tanto, un control mejor de una población plaga se obtendría al introducir un parásito foráneo. Esta posibilidad se plantearía con S. carpocapsae pero las condiciones ambientales donde este nemátodo ha evolucionado difieren notablemente de aquellas donde se encuentra este insecto plaga, pues la mejor sobrevivencia de este nemátodo se obtiene en rangos de temperatura que oscilan entre 5°C y 15°C; así, al implementar un control con esta especie de nemátodo se haría necesario realizar liberaciones sucesivas ya que al presentarse un ecosistema de suelos cálidos, tendría como limitante la temperatura, lo que incrementaría los costos de control, mientras que las razas de H. bacteriophora están adaptadas a las condiciones donde se encuentra C. bergi, por lo tanto se pueden establecer en campo, con lo cual se buscaría provocar una epizootia que disminuya la población hospedera y el control sería más prolongado y menos costoso.
Conclusiones
Las razas LFR92 y SQC92 de Heterorhabditis bacteriophora son capaces de parasitar todos los estados de desarrollo de Cyrtomenus bergi.
Las razas LFR92 y SQC92 de H. bacteriophora no presentaron una capacidad parasítica diferencial sobre los cuatro primeros estados de desarrollo de C. bergi; por el contrario sobre ninfas de 5º estadío y adultos el parasitismo sí fue diferencial.
Rectas de comportamiento para las razas SQC92 y LFR92 de H. bacteriophora, sobre el 5º estado de desarrollo de C. bergi según análisis probit.
El 5º estado de desarrollo de C. bergi es el más susceptible al ataque de ambas razas de H. bacteriophora.
C. bergi presentó una respuesta diferencial a varias dosis de los nemátodos de ambas razas de H. bacteriophora, haciéndose el parasitismo mayor al aumentar la dosificación.
Para la raza LFR92 de H. bacteriophora se obtuvo una DL50 de 702 n/ml, con límites de confianza 81 y 1551 y para SQC92 de 826 n/ml con límites 545 y 1137.
References
1.
BeddingR. A. (1981). Low cost in vitro mass production of Neoaplectana and Heterorhabditis species (Nematoda) for field control of insect pests. Nematologica, 27, 109–114.
2.
BeddingR. A. (1984). Large scale production, storage and transport of the insect parasitic nematodes Neoaplectana spp and Heterorhabditis spp. Annals of Applied Biology, 104, 117–120.
3.
BeddingR. A.MolyneuxA. S.ArhurstR. J. (1983). Heterorhabditis spp., Neoaplectana spp., and Steinernema kraussei: interspecific and intraspecific differences in infectivity for insects. Experimental Parasitology, 55, 249–257.
4.
CaicedoA. M.BellottiA. C. (1994). Evaluación del potencial del nematodo entomógeno Steinernema carpocapsae Weiser (Rhabditida: Steinernematidae) para el control de Cyrtomenus bergi Froeschner (Hemiptera: Cydnidae) en condiciones de laboratorio. Revista Colombiana de Entomología, 20(4), 241–246.
5.
EpskyN. D.CapineraJ. L. (1994). Invasion efficiency as a measure of efficacy of the entomogenous nematode Steinernema carpocapsae (Rhabditida: Steinernematidae). Journal of Economic Entomology, 87(2), 366–370.
6.
GarciaO.BellottiA. C. (1980). Estudio preliminar de la biología y morfología de Cyrtomenus bergi Froeschner nueva plaga de la yuca. Revista Colombiana de Entomología, 6, 3–4.
7.
GeorgisR. (1990). Commercialization of Steinernematid and Heterorhabditid entomopathogenic nematodes. Brighton Crop Protection Conference. Pests and Diseases: 275–280.
8.
GeorgisR.GauglerR.KayaH. K. (1991). Effect of Steinernematid and Heterorhabditid nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) on nontarget arthropods. Environmental Entomology, 20(3), 815–822.
9.
HomA. (1994). Current status of entomopathogenic nematodes. The IPM practitioner, XVI(3), 1–12.
10.
JanssonR. K.LecloneS. H.GauglerR. (1993). Field efficacy and persistence of entomopathogenic nematodes (Rhabditida: Steinernematidae, Heterorhabditidae) for control of sweetpotato weevil (Coleoptera: Apionidae) in southern Florida. Journal of Economic Entomology, 86(4), 1055–1063.
11.
KleinM. G. (1990). Efficacy against soil-inhabiting insect-pests. In Entomopathogenic nematodes in biological control. Ed. CRC PressBoca Raton. p. 195–215.
12.
LindegrenJ. E.WongT. T.McInnisD. O. (1990). Response of mediterranean fruit fly (Diptera: Tephritidae) to the entomogenous nematode Steinernema feltiae in field tests in Hawaii. Environmental Entomology, 19(2), 383–386.
13.
MannionC. M.JanssonR. K. (1992). Movement and postinfection emergence og entomopathogenic nematodes from sweetpotato weevil, Cylas formicarus (F.) (Coleoptera: Apionidae). Biological Control, 2, 297–305.
14.
PoinarG. O.Jr. (1990). Biology and taxonomy of Steinernematidae and Heterorhabditidae. In Entomopathogenic nematodes in biological control. Ed. CRC PressBoca Raton p. 23–63.
15.
RiisL. (1990). La chinche subterránea Cyrtomenus bergi, una plaga de importancia creciente en América Latina: Estudios de comportamiento, fluctuación poblacional, control botánico con referencia especial a yuca [M. Sc. Thesis]. Copenhagen, Denmark: Institute of ecology and molecular biology. The Royal Veterinary and Agricultural University.
16.
SelvanS.GauglerR.CampbellJ. F. (1993). Efficacy of entomopathogenic nemtadode strains against Popillia japonica (Coleoptera: Scarabaeidae) larvae. Journal of Economic Entomology, 86(2), 353–359.
17.
WoodringJ. LKayaH. K. (1988). Steinernematid and Heterorhabditid nematodes: A handbook of techniques. Southern Cooperative Series Bulletin 331. Arkansas Agricultural Experiment Station, Fayeteville.