
Abstract
Helicobacter pylori (Hp) infection is associated with the development of gastric lesions including gastritis, intestinal metaplasia (IM), and gastric carcinoma. In humans, Hp is found almost exclusively in the foveolar epithelium of the gastric mucosa and rarely colonizes the deeper portions where mucous cells of the glands produce mucins with terminal α1, 4-GlcNAc O-glycans. This structure exerts antimicrobial activity against Hp. The development of IM in the stomach is characterized by Hp clearance from the metaplastic glands and by major alterations in the expression of mucins and mucin-carbohydrates. The present work evaluated whether terminal α1,4-GlcNAc and sialyl-Tn antigen are implicated in the process of Hp clearance from metaplastic glands by analyzing the expression of these antigens in different types of IM—complete (n=12) and incomplete (n=8)—and in gastric cell lines. Terminal α1,4-GlcNAc was not detected in IM except in a single foci of one case, indicating that this structure is not implicated in the clearance of Hp from IM, in contrast to what is observed in normal gastric mucosa. None of the gastric carcinoma cell lines studied showed terminal α1,4-GlcNAc, suggesting that they do not display a gastric gland mucous cell phenotype and therefore are useful models for in vitro Hp studies. Finally, sialyl-Tn antigen colocalizes with MUC2 mucin and is present in all cases of complete and incomplete IM, suggesting that either or both can be implicated in Hp clearance from IM.
Keywords
H
Hp is observed almost exclusively colonizing the gastric mucosa. It is also found in the duodenal mucosa whenever there is gastric metaplasia (Futami et al. 1999), suggesting that the colonization is dependent on the gastric microenvironment. In gastric mucosa, the exclusive adherence of Hp to surface mucous cell-type cells also indicates that the bacteria specifically recognize epithelial cell surface constituents. Two carbohydrate structures, Lewis b expressed on mucins and sialyl dimeric Lewis × expressed on glycolipids, have been described as specific ligands for Hp adhesins, BabA, and SabA, respectively (Ilver et al. 1998; Mahdavi et al. 2002).
Absence of colonization of the deeper portions of gastric mucosa has been attributed to the mucin glycosylation of gland mucous cells, which produce mucin carbohydrate chains having terminal α1,4-GlcNAc residues attached to core 2-branched O-glycans [GlcNAcα1 → 4Galβ1 → 4GlcNAcβ1 → 6(GlcNAcα1 → 4Galβ1 → 3)→GalNAcα→Ser/Thr] (Nakayama et al. 1999; Zhang et al. 2001). These terminal structures have been shown to exhibit antimicrobial activity against Hp due to the inhibition of the biosynthesis of cholesteryl-α-D-glucopyranoside, a major cell wall component of the bacteria (Kawakubo et al. 2004).
Development of gastric IM creates a microenvironment that is hostile to the bacterial colonization and generally leads to clearing of Hp from metaplastic glands (Craanen et al. 1992). Major alterations in mucin expression and their associated carbohydrate chains have been described in IM. These alterations include the aberrant expression of the MUC2 mucin and expression of the sialyl-Tn antigen, which are markers normally expressed in intestinal mucosa (David et al. 1992; Reis et al. 1998; Corfield et al. 2001).
The aim of this study was to characterize the expression pattern of the terminal α1,4-GlcNAc and the sialyl-Tn antigen in both complete IM and incomplete IM, to evaluate whether these glycans are implicated in the process of Hp clearance from metaplastic glands.
Materials and Methods
Human Tissue Samples
Gastric biopsies were obtained from individuals with non-ulcer dyspepsia. Gastric mucosas adjacent to carcinomas were obtained from individuals undergoing surgery at the Hospital S. João, Medical Faculty (Porto, Portugal). All specimens were fixed in 10% formalin and routinely embedded in paraffin wax. Serial sections were cut and used for immunohistochemistry and immunofluorescence. We evaluated 22 cases with IM that were classified as complete IM (n=14) and incomplete IM (n=8) according to the pattern of mucin expression as described below.
Cell Lines and Cell Culture
Five gastric carcinoma cell lines (MKN45, KATOIII, GP202, GP220, and AGS) were studied. GP202 and GP220 cell lines were established in our laboratory (Gartner et al. 1996). MKN45 was obtained from the Japanese Collection of Research Bioresources (JCRB; Osaka, Japan). KATOIII and AGS are commercially available cell lines (American Type Culture Collection; Rockville, MD).
Cell lines were grown in RPMI 1640 with Glutamax and supplemented with 10% inactive FBS and 50 μg/ml gentamicin. Cultures were maintained at 37C in a humidified 5% CO2 atmosphere.
Monoclonal Antibodies, Immunohistochemistry, and Immunofluorescence
Monoclonal antibodies (MAbs) used in this study, their specificity, and references are listed in Table 1. Immunohistochemistry was performed using the avidin-biotin complex method. Paraffin sections were dewaxed and rehydrated. Samples designated for MUC2 staining were pretreated with neuraminidase for exposure of the epitope detected by antibody PMH1 (Reis et al. 1998).
Monoclonal antibodies, their specificity, and references
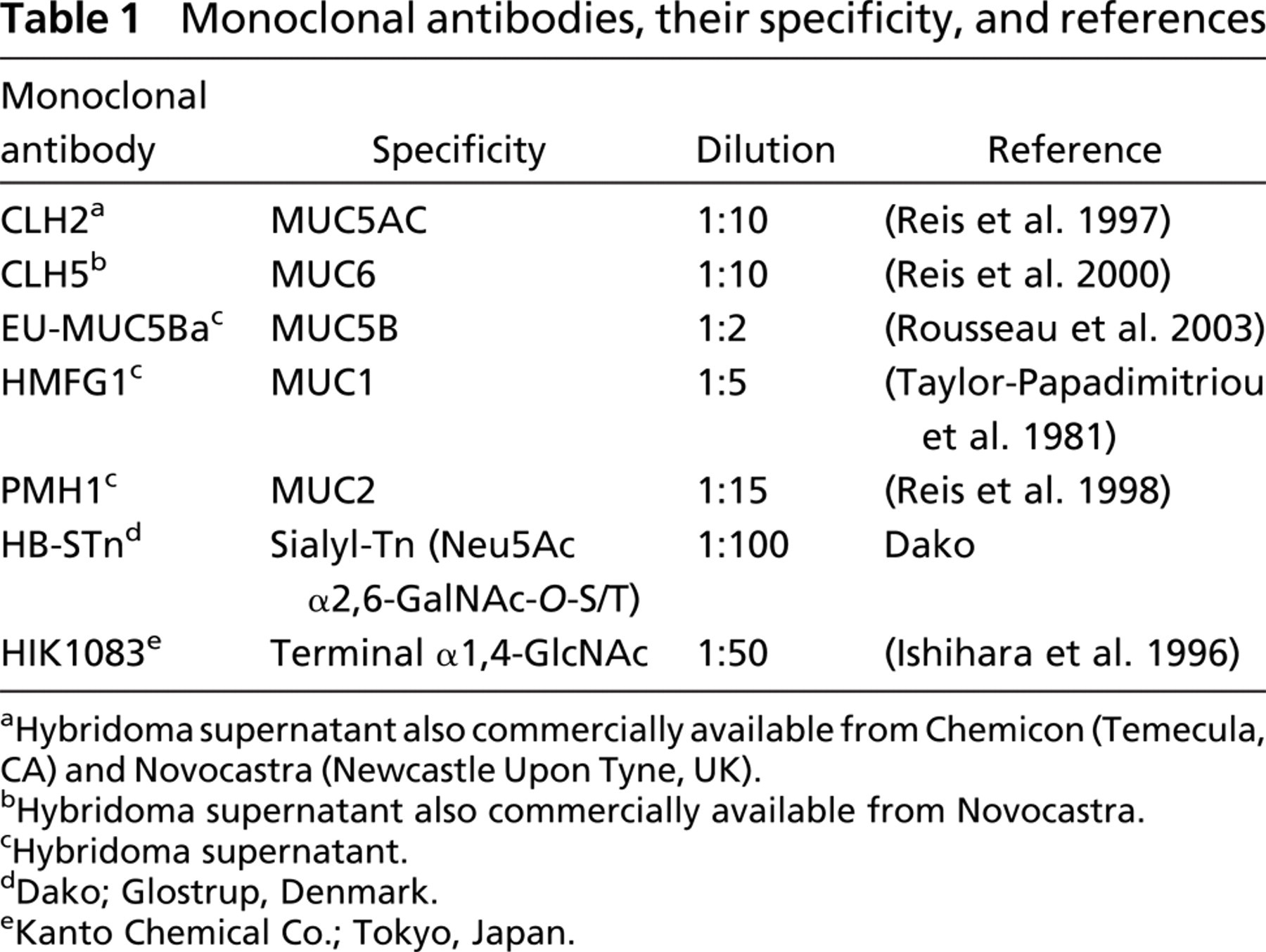
Hybridoma supernatant also commercially available from Chemicon (Temecula, CA) and Novocastra (Newcastle Upon Tyne, UK).
Hybridoma supernatant also commercially available from Novocastra.
Hybridoma supernatant.
Dako; Glostrup, Denmark.
Kanto Chemical Co.; Tokyo, Japan.
Neuraminidase from Clostridium perfringens type IV (Sigma; St Louis, MO) was used diluted in 0.1 M sodium acetate buffer (pH 5.5) to a final concentration of 0.1 U/ml. Incubation was carried out for 2 hr at 37C and was followed by three washes in cold water. Sections were treated with 0.5% hydrogen peroxide in methanol for 30 min, washed with TBS, pH 7.6, and followed by a 20-min incubation with rabbit non-immune serum diluted 1:5 in TBS containing 10% BSA. Sections were washed and incubated overnight at 4C with MAbs (Table 1) diluted in TBS containing 5% of BSA. Sections were washed three times in TBS, incubated with a biotin-labeled rabbit anti-mouse secondary antibody diluted 1:200 in TBS for 30 min, washed in TBS, and incubated with avidin-biotin-peroxidase complex for 1 hr. Sections were washed and stained for 7 min with 0.05% DAB freshly prepared in 0.05 M Tris/hydroxymethylaminomethane buffer, pH 7.6, containing 0.1% hydrogen peroxide. Negative controls were performed using conjugated secondary antibody alone. Control for neuraminidase treatment was performed by incubating the section without enzyme.
Single immunofluorescence labeling was performed in cell lines fixed in acetone for 5 min. Samples were washed twice in TBS and incubated for 20 min with rabbit non-immune serum diluted 1:5 in TBS containing 10% BSA. Samples were washed in TBS and incubated overnight at 4C with MAbs diluted in TBS containing 5% BSA. Sections were washed three times for 5 min in TBS and incubated with FITC-conjugated rabbit anti-mouse immunoglobulin (Code F-261; Dako, Glostrup, Denmark) diluted 1:70 in TBS.
Samples designated for double-labeling immunofluorescence were treated as described above for single immunofluorescence labeling, followed by washing twice for 5 min in TBS and blocked with non-immune goat serum diluted 1:10 in TBS. Sections were incubated with MAbs PMH1 or HIK1083 (mouse IgM; Table 1) overnight at 4C. Sections were washed three times for 5 min with TBS and incubated for 45 min with Texas red-conjugated goat anti-mouse IgM (Jackson Immunoresearch Laboratories; West Grove, PA) diluted 1:50 in TBS. Sections were washed three times for 5 min in TBS and with DAPI, 20 min in the dark. Samples were washed three times for 5 min in TBS and mounted in Vectashield (Vector Laboratories; Burlingame, CA).
Results
Expression of Mucins and Carbohydrate Antigens (Terminal α1,4-GlcNAc and Sialyl-Tn) in Normal Gastric Mucosa
Normal gastric mucosa showed a consistent expression of MUC5AC in superficial foveolar cells and MUC6 in mucous neck cells of the body and deeper glands of the antrum (Figure 1A). MUC1 was observed in the superficial foveolar cells of the antrum and in oxynthic glands of the body. MUC2 and MUC5B were not detected in normal gastric mucosa (Figure 1A).
Expression of terminal α1,4-GlcNAc was restricted to mucous neck cells of the body and deeper glands of the antrum (Figure 1A and Figure 2A). Expression of terminal α1,4-GlcNAc did not fully colocalize with MUC6. Immunostaining of terminal α1,4-GlcNAc was observed throughout the cytoplasm and membrane, whereas MUC6 immunostaining was restricted to the perinuclear area of the cytoplasm. Expression of sialyl-Tn antigen was not detected in normal gastric mucosa.
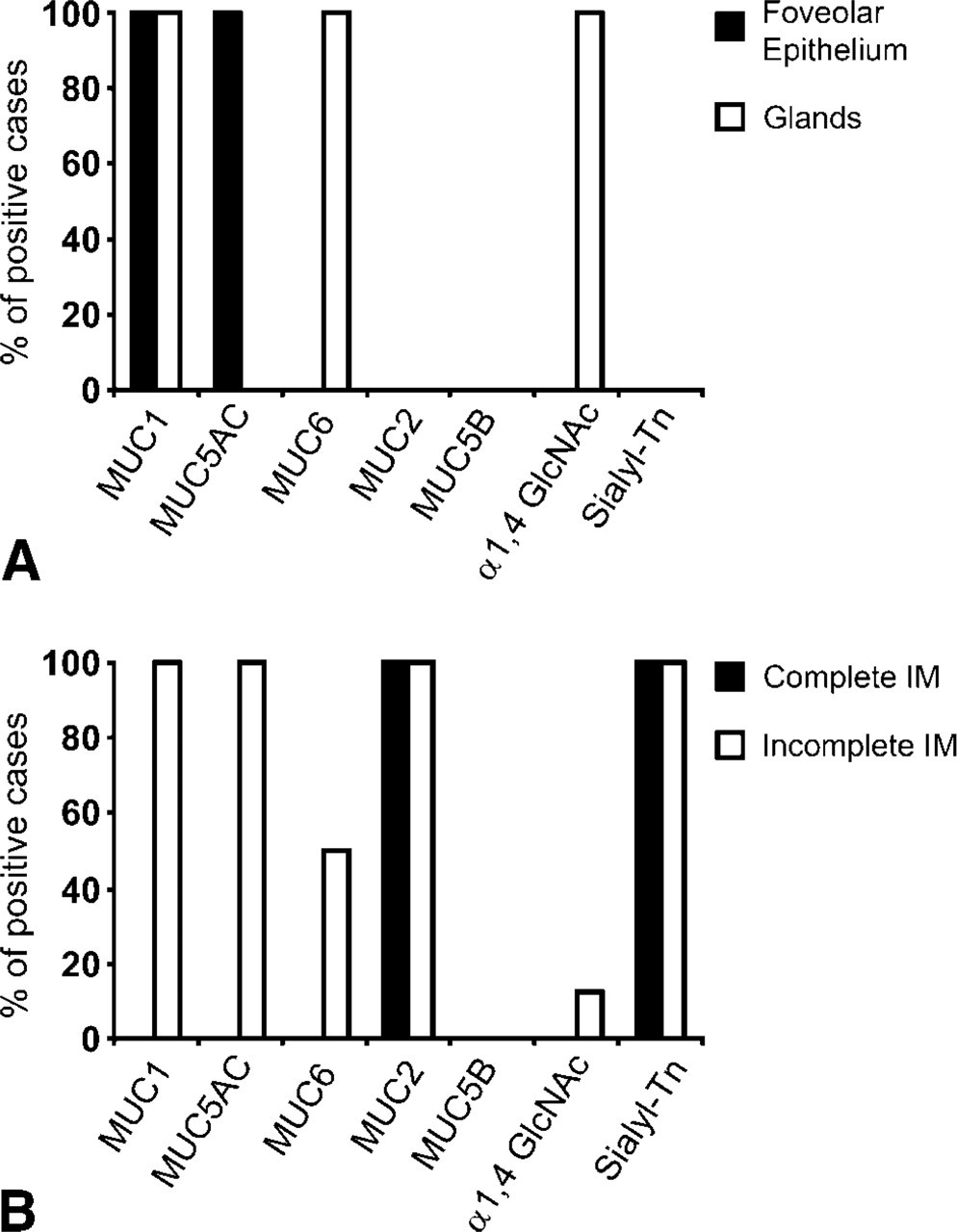
Expression profile of mucins (MUC1, MUC2, MUC5AC, MUC5B, and MUC6) and carbohydrate antigens (terminal α1,4-GlcNAc and sialyl-Tn). (
Expression of Mucins and Carbohydrate Antigens (Terminal α1,4-GlcNAc and Sialyl-Tn) in Complete and Incomplete Intestinal Metaplasia
The 22 cases of IM were classified as complete IM (n=14) or incomplete IM (n=8) according to the pattern of expression of mucins (Reis et al. 1999). The cases with complete IM showed absence of expression of gastric mucins MUC5AC, MUC6, and MUC1 and expression of the intestinal mucin MUC2 (Figure 1B and Figure 2C). The cases with incomplete IM showed coexpression of “gastric mucins” MUC5AC, MUC6, and MUC1 together with the intestinal mucin MUC2 (Figure 1B and Figure 2F). Both complete and incomplete IM showed absence of expression of MUC5B (Figure 1B).
Results of the expression of terminal α1,4-GlcNAc and sialyl-Tn antigen are summarized in Figure 1B. The 11 foci of complete IM showed no expression of terminal α1,4-GlcNAc as detected by HIK1083. Expression of sialyl-Tn was detected in all cases of incomplete IM.
In incomplete IM, most foci were negative for terminal α1,4-GlcNAc, and only one case showed a single metaplastic gland of the incomplete IM type with expression of terminal α1,4-GlcNAc. Expression of sialyl-Tn was detected in all cases with incomplete IM.
Expression of Mucins and Carbohydrate Antigens (Terminal α1,4-GlcNAc and Sialyl-Tn) in Gastric Carcinoma Cell Lines
Expression of carbohydrate antigens is summarized in Table 2. Terminal α1,4-GlcNAc was absent in all gastric cell lines studied. Low levels of expression of sialyl-Tn were detected in MKN45 cells.
Expression of MUC5AC was detected in <25% of MKN45, KATOIII, and GP220 cells. A diffuse cytoplasmic staining pattern was observed. None of the gastric cell lines studied showed expression of the intestinal mucin MUC2 (Table 2).
Discussion
Development of IM in human stomach creates a microenvironment that is hostile to the bacterial colonization leading to the clearing of Hp from metaplastic glands (Craanen et al. 1992; Genta et al. 1996). Given the role that carbohydrate structures play in the adhesion and/or colonization of Hp, we characterized the alteration of two carbohydrate structures, terminal α1,4-GlcNAc and sialyl-Tn antigen, in both complete and incomplete IM to evaluate whether these glycans are implicated in the process of Hp clearance from metaplastic glands.
Evaluation of terminal α1,4-GlcNAc in normal gastric mucosa confirmed previous studies (Nakamura et al. 1998,1999; Zhang et al. 2001) showing that terminal α1,4-GlcNAc was restricted to the mucous cells of the glands of the gastric mucosa with absence of expression in the foveolar superficial gastric epithelium. These observations are in agreement to the hypothesis that the expression of terminal α1,4-GlcNAc in gastric glands creates hostile conditions for Hp colonization in deep gastric mucous cells of the glands. This antimicrobial activity of terminal α1,4-GlcNAc against Hp has been shown to stem from the inhibition of the biosynthesis of cholesteryl-α-D-glucopyranoside, a major cell wall component of the bacteria (Kawakubo et al. 2004).
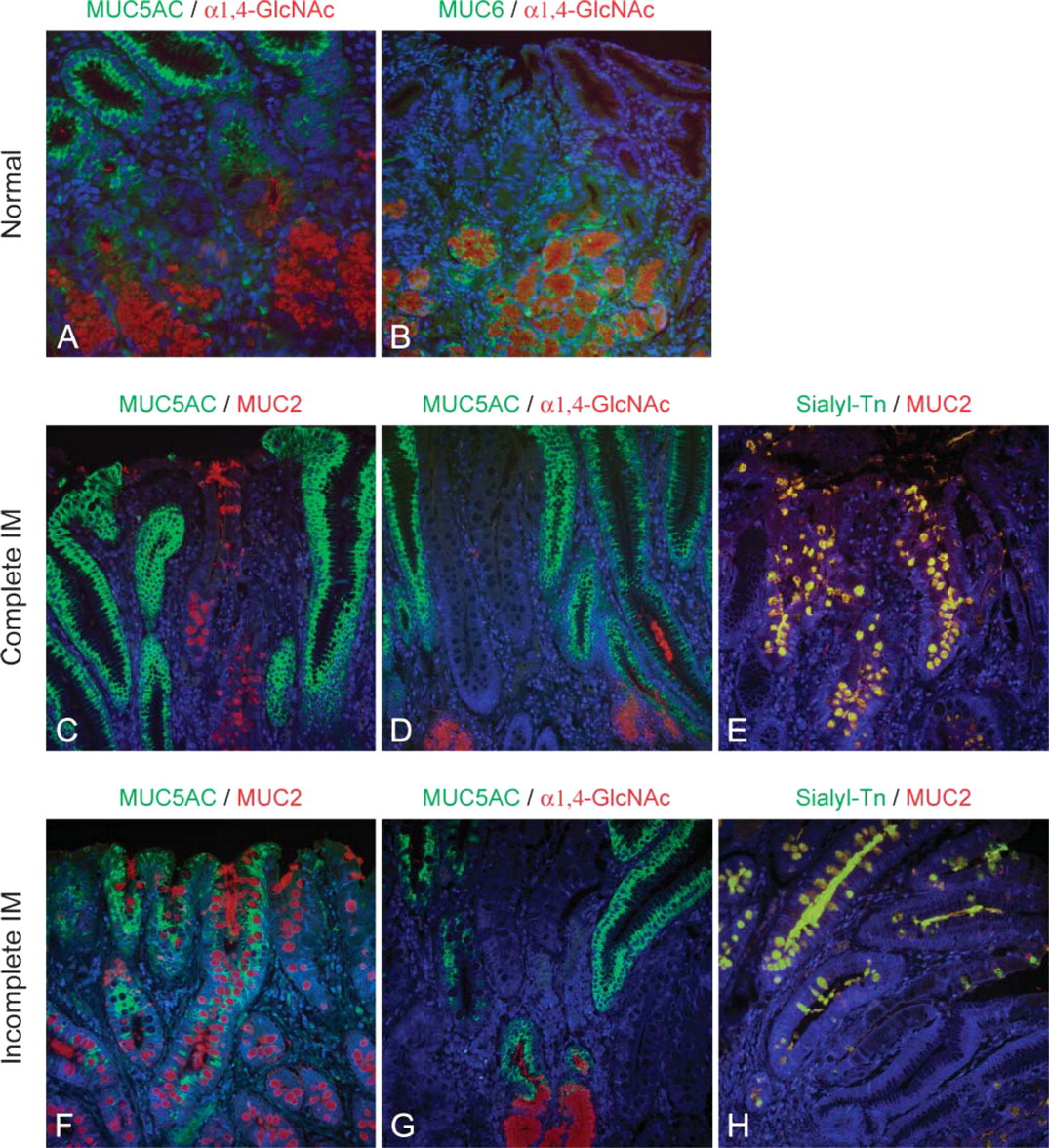
Immunofluorescence of the expression of terminal α1,4-GlcNAc, sialyl-Tn, and mucins MUC5AC and MUC2 on normal gastric mucosa, complete IM, and incomplete IM. (
Expression of terminal α1,4-GlcNAc, sialyl-Tn, and mucins MUC5AC, MUC6, and MUC2 in gastric cell lines

Scoring of labeling intensity: -negative; +weakly positive; ++moderately positive; +++strongly positive.
NA, not applicable.
Comparison of expression of mucins and carbohydrate antigens in normal gastric mucosa shows that both MUC6 and terminal α1,4-GlcNAc are expressed in mucous cells of the glands of the gastric mucosa but do not fully colocalize at the tissue and cellular level. Lack of overlap in the immunocytolocalization pattern may be due to variability of the type and rate of glycosylation of MUC6 mucin, which insomecases displays glycoforms that may not be detected by the MAb CLH5 (Reis et al. 2000) but still express terminal α1,4-GlcNAc.
We did not observe expression of the simple mucin-type carbohydrate antigen sialyl-Tn in normal gastric mucosa in agreement with what was previously described (David et al. 1992). This is in keeping with data showing that sialyl-Tn is a carbohydrate structure rarely expressed in normal tissues, despite its ability to be detected in normal intestinal mucosa where it shows restricted expression in the goblet cells (Itzkowitz et al. 1990).
The almost exclusive colonization of Hp of the gastric mucosa and the localized adherence restricted to the surface foveolar cells (Hidaka et al. 2001; Teixeira et al. 2002) suggest that Hp colonization is dependent on the gastric microenvironment. Development of IM in the stomach is characterized by extensive modifications in the profile of mucins (Reis et al. 1999) and mucin-glycosylation (Torrado et al. 1990; Mullen et al. 1995; Silva et al. 2002; Bodger et al. 2003). In the present study we characterized the expression of mucins MUC1, MUC2, MUC5AC, MUC5B, and MUC6 using specific MAbs. Detection of MUC2 was performed using the MAb PMH1, which detects MUC2-GalNAc (Reis et al. 1998). To ensure the exposure of the PMH1 epitope and its detection, we performed the removal of NeuAc by submitting the tissue sections to neuraminidase treatment as previously described (Reis et al. 1998,1999). Neuraminidase treatment did not affect the immunostaining obtained with the antibodies directed to MUC1, MUC5AC, MUC5B, MUC6, or α1,4-GlcNAc; therefore, neuraminidase treatment was unnecessary with these antibodies (data not shown).
Characterization of the pattern of mucin expression in IM disclosed two major types: (a) the complete IM showing absence of expression of gastric mucins (MUC5AC, MUC6, and MUC1) and de novo expression of the intestinal mucin MUC2 and (b) the incomplete IM displaying a coexpression of “gastric mucins” MUC5AC, MUC6, and MUC1 together with the intestinal mucin MUC2 (Reis et al. 1999). The present study also confirmed previous observations showing that MUC5B is not expressed in normal gastric mucosa or in intestinal metaplasia (Perrais et al. 2001; Pinto-de-Sousa et al. 2004).
We, and others, have shown that most areas of IM show no Hp colonization (Genta et al. 1996; Bravo and Correa 1999; Teixeira et al. 2002), and we have previously characterized that the frequency of Hp colonization in complete and incomplete IM is 0% and 25%, respectively (Teixeira et al. 2002). We also observed that, in these rare positive cases, Hp colonization was restricted to small IM foci lacking MUC2 expression. These observations, together with evidence of the role that carbohydrate structures play in the colonization by Hp, led us to the characterization of complete and incomplete IM with respect to carbohydrate structures: terminal α1,4-GlcNAc and sialyl-Tn antigen.
Contrary to the normal gastric mucosa, which showed a consistent expression of terminal α1,4-GlcNAc in the gastric glands, we observed no expression of this glycan in IM. Both complete and incomplete IM were negative for terminal α1,4-GlcNAc. One case of incomplete IM was an exception showing a single metaplastic gland with expression of terminal α1,4-GlcNAc among several metaplastic glands showing no expression of terminal α1,4-GlcNAc. The general absence of expression of terminal α1,4-GlcNAc in IM observed in the present study demonstrates that this carbohydrate structure is not responsible for the absence of colonization of Hp observed in metaplastic glands (Craanen et al. 1992; Teixeira et al. 2002). Other mechanisms, such as absence of ligands for Hp adhesins, together with the aberrant expression of MUC2 and sialyl-Tn, may determine major modifications in IM contributing to the creation of a hostile microenvironment for Hp colonization of metaplastic glands.
The sialyl-Tn antigen, a carbohydrate structure normally expressed in intestinal mucosa, was observed in all cases of both complete and incomplete IM. Sialyl-Tn expression was localized in goblet cells, and double-labeling immunofluorescence demonstrated a consistent overlapping between sialyl-Tn and MUC2 expression (Figures 2E and 2H). The consistent expression of sialyl-Tn and MUC2 in all IM cases suggests that either or both can be implicated in Hp clearance from the gastric mucosa with IM lesions.
Given the antimicrobial effect that terminal α1,4-GlcNAc may exert (Kawakubo et al. 2004), it is important to characterize the expression of this glycan in cell lines used for coculture experiments with Hp. We have observed that Hp can adhere and be cocultured with gastric cell lines MKN45, AGS, and GP202 (unpublished data), which do not express terminal α1,4-GlcNAc. However, Kawakubo et al. (2004) demonstrated that when Hp was cocultured with the cell line AGS transfected with α4GlcNAc-transferase and expressing α1,4-GlcNAc, Hp growth was markedly suppressed, and the cellular damage induced by Hp was barely detected. These observations demonstrate that gastric cells expressing α1,4-GlcNAc-capped O-glycans protect themselves against Hp infection. Our results showed no expression of terminal α1,4-GlcNAc with the gastric carcinoma cell lines studied (Table 2), demonstrating that the cell lines used in the present and in previous studies (Lee et al. 2004; Keates et al. 2005) are suitable models for Hp coculture/adhesion experiments.
Our results are identical to previous studies with the same cell lines except for slight discrepancies regarding levels of mucin expression in the cell lines when compared with previous studies (Carvalho et al. 1999; Basque et al. 2001). This variability among studies may stem from the different confluence levels of the cell culture at the time of immunocytology sampling.
In conclusion, terminal α1,4-GlcNAc was not detected in IM except in a single foci of one case, indicating that this structure is not implicated in the clearance of Hp from IM, in contrast to what is observed in normal gastric mucosa. None of the gastric carcinoma cell lines studied showed terminal α1,4-GlcNAc, suggesting they do not display a gastric gland mucous cell phenotype and therefore are useful models for in vitro Hp studies. Finally, sialyl-Tn antigen colocalizes with MUC2 mucin and is present in all cases of complete and incomplete IM, suggesting that either or both can be implicated in Hp clearance from IM.
Footnotes
Acknowledgments
This work was supported by Fundação para a Ciěncia e a Tecnologia (FCT, POCI/SAU-OBS/56686/2004) and Association for International Cancer Research (Grant 05-088). N.M. acknowledges Fundação para a Ciěncia e a Tecnologia (FCT, Ref. SFRH/BD/11764/2003) for financial support.
We thank Nuno Mendes and Paula Silva for technical assistance. We thank Prof. Dallas Swallow for providing antibody EU-MUC5Ba and Dr. Joy Burchell and Prof. Joyce Taylor-Papadimitriou for providing antibody HMFG1.