
Abstract
Sialyl-Tn (STn), a mucin-associated disaccharide antigen carried by apomucins such as MUC2, plays an important role in tumor biology. However, little is known about the subcellular localization and compartments involved in STn synthesis. In this study we used immunoelectron microscopy to localize STn and MUC2 apomucin in human colorectal tissues. MUC2 apomucin was localized predominantly in the rough endoplasmic reticulum (RER) in normal colorectal mucosa (n=6), colorectal adenoma (n=8), and colorectal adenocarcinoma (n=10). STn, recognized by monoclonal antibody TKH2, was not readily detectable in normal colorectal mucosa but becomes manifest in both trans-Golgi apparatus and mucin droplets in colorectal adenoma. In colorectal adenocarcinoma, STn was localized not only in late but also in early Golgi compartments, and particularly in some RER lumens. Furthermore, electron microscopic in situ hybridization revealed that gold particles representing MUC2 mRNA are primarily localized over the RER. Our findings indicate that in colorectal adenoma STn sialylation takes place in the trans-Golgi apparatus, whereas in colorectal cancer STn sialylation occurs in all the Golgi compartments and in the RER.
Sialyl-Tn (STn) is a mucin-associated disaccharide (Itzkowitz 1992). Extensive studies during the past decade have demonstrated that STn is usually cryptic in normal tissues but becomes highly expressed in malignant tumors, especially those arising from the colorectal mucosa (Yonezawa et al. 1992; Kim 1993; Kim et al. 1996). In normal colorectal mucosa, the cryptic nature of STn expression has been ascribed to O-acetylation of sialic acid residues of STn antigen, but in neoplastic colorectal cells STn is synthesized as a non-O-acetylated form (Corfield et al. 1995; Jass et al. 1995; Ogata et al. 1995; Klein and Roussel 1998). Recent studies showing STn expression in premalignant lesions, such as colorectal adenomas, point to a higher tendency of colon cells towards neoplastic transformation (Itzkowitz et al. 1992; Cao et al. 1997; Matsushita et al. 1997; Yamachika et al. 1997). STn expression in colon mucosal cells has also been reported to precede dysplasia in ulcerative colitis by several years (Itzkowitz et al. 1996). Clinically, STn-positive colorectal cancers often correlate with adverse outcome (Bresalier et al. 1996; Sternberg et al. 1996; Imada et al. 1999; Nakagoe et al. 2000). In addition, mucinassociated STn has been shown to be a potent inhibitor of NK-cell cytotoxicity of target tumor cells (Ogata et al. 1992). Furthermore, STn-based cancer vaccination shows a therapeutic potential for patients with epithelial cancers (MacLean et al. 1996). These data indicate that STn is not only a unique tumor marker but also plays an important role in tumor biology, although the pathogenic significance of STn in carcinogenesis per se remains to be determined.
Biochemically, STn is formed by the sialylation of Tn antigen, i.e., GalNAc-O-Ser/Thr, which represents the common core O-glycan carbohydrate structure (Itzkowitz 1992; Kim 1993). The sialylation process for STn is carried out by a specific glycosyltransferase called CMP-NeuAc: GalNAc-peptide α2,6-sialyltransferase (STn sialyltransferase) (Itzkowitz 1992; Yang et al. 1994). Sialyltransferases are a large family, the specificity of each depending on carbohydrate sequence and anomeric linkages (Tsuji 1996; Yamada et al. 1997). Although the subcellular compartments for sialylation have been studied for several other sialylated oligosaccharides, such as NeuAcα2,3Galβ1, 3GalNAc and NeuAcα2,6Galβ1, 4GlcNAc (Roth 1987; Whitehouse et al. 1997), little is known about the subcellular localization and the compartments involved in the biosynthesis of STn antigen. In this study we examined the subcellular localization and biosynthetic pathway of STn in normal, premalignant, and malignant human colorectal tissues using high-resolution immunoelectron microscopy. By comparing the patterns of STn subcellular localization sites, we have been able to define the alterations in STn sialylation compartments in colorectal adenoma and adenocarcinoma. The data obtained in this study provide new insights for understanding the subcellular events that lead to the abnormal expression of both STn and MUC2 apomucin during neoplastic transformation of colorectal epithelial cells.
Materials and Methods
Tissue Specimens
Six normal colorectal mucosae were obtained from endoscopically resected tissues from patients who were verified to have no malignant colorectal diseases. Eight colorectal adenomas came from endoscopic polypectomy and were divided into two categories: low-grade atypia and high-grade atypia. Ten well- to moderately differentiated primary colorectal adenocarcinoma tissues were obtained from patients undergoing surgical resection. Tissue specimen collection was in accordance with human study guidelines and approval. The separation of adenomas into low and high grades and the diagnosis of adenocarcinoma were performed according to the definitions and explanatory notes by the World Health Organization (Jass and Sobin 1989). Of the 10 adenocarcinomas, four cases were found to have lymph node and distant metastasis.
Fresh tissues taken from several locations of each case were cut into fine pieces and fixed in freshly prepared 4% paraformaldehyde (Sigma Chemical; St Louis, MO) in 0.1 M phosphate buffer, pH 7.4, at 4C for 3 hr. Fixed tissues were then washed overnight in PBS, pH 7.4. On average, 10 tissue blocks from each patient were chosen and processed for embedding in LR White (Polyscience; Warrington, PA) as described by Timms (1986). In short, tissue blocks were placed in the bottoms of 00 gelatin capsules (Lilly Pharmaceuticals; Indianapolis, IN). The capsules were filled with LR White, sealed with tops, and polymerized at 50C for 24 hr. Semi-thin sections were cut for proper tissue localization and diagnostic evaluation. For the final subcellular labeling, only those blocks that were in conformity with the histopathological diagnosis of the patients were selected. For immunoelectron microscopic (IEM) labeling, thin sections (~80 nm) were collected on 300-mesh nickel grids. For EM in situ hybridization (ISH), thin sections (~100 nm) were attached onto carbon/Formvar-coated 200-mesh nickel grids.
Antibodies
The murine monoclonal antibody (MAb) TKH2 (IgG1) was obtained from Ohtsuka Pharmaceutical (Tokushima, Japan). This antibody specifically detects STn epitopes residing in the mucin glycoproteins (Kjeldsen et al. 1988). Polyclonal antibody anti-MRP (purified rabbit IgG) was used to identify the tandem repeat peptide of MUC2 apomucin. Anti-MRP was produced by using a synthetic MUC2 peptide containing the 23-amino-acid threonine rich tandem repeat peptide of MUC2 apomucin, PTTTPISTTTMVTPTPTPTGTQT (Gum et al. 1989). Purified sheep anti-digoxigenin antibody was purchased from Boehringer-Mannheim (Indianapolis, IN). Rabbit antisheep IgG antibody came from Jackson ImmunoResearch Laboratories (West Grove, PA). Colloidal gold (15-nm)-labeled goat anti-rabbit IgG (GAR-G15), goat anti-mouse IgG (GAM-G15), and protein A (PA-G15) were obtained from Amersham Life Science (Arlington Heights, IL).
MUC2 cDNA Labeling Procedure
The MUC2 probe pHAM1, a 90-bp portion of the MUC2 tandem repeat inserted at the Eco RI site of Bluescript SK-, which was isolated from a tracheal cDNA library12, was kindly provided by Dr. Carol B. Basbaum (Department of Anatomy, Cardiovascular Research Institute, University of California, San Francisco). Digoxigenin-labeled pHAM1 DNA probe and digoxigenin-labeled pHAM1 RNA probes (both antisense and sense) were prepared as described previously (Yonezawa et al. 1997).
EM Immunocytochemical Labeling Procedure
For EM immunocytochemical labeling, colloidal gold was chosen on the basis of its unique properties, such as precise localization and easy differentiation without obscuring fine cell architectures (Roth et al. 1996). On the other hand, silver enhancement or intensification of gold probes provided additional advantages to the gold labeling technique by allowing double labeling using the same size gold particles without interfering with the visualization of label signals in the second sequence and, more importantly, at the same time eliminating the possible crossreactions that are usually difficult to get rid of by other methods (van den Pol 1985; Bienz et al. 1986). The present EM immunocytochemical labeling was conducted according to published protocols (Bienz et al. 1986; Egea et al. 1993) with modifications. Briefly, the grids were held vertically by snapping portions of their edges into fine slots pre-made in a silica rubber plate for convenient, time-saving manipulation. Labeling was accomplished by applying droplets of various reagents to the immobilized grids. MUC2 and STn double labeling was performed sequentially at room temperature (RT) as follows: (a) rinse in Tris-buffered saline (TBS), pH 7.4, for 10 min; (b) block in 5% normal goat serum for 15 min; (c) incubate with the anti-MRP MUC2 antibody (1:300) for 2 hr; (d) wash in TBS three times for 10 min; (e) incubate with GARG15 (1:40) for 1 hr; (f) “jet-wash” with TBS (pH 8.2) and rinse in the same buffer three times for 10 min; (g) rinse in distilled water three times for 3 min. The MUC2-labeled grids were then silver-enhanced for 6 min using the Inten-SEM kit (Amersham Life Science) according to the manufacturer's instructions. We found that 6-min enhancement could enlarge 15-nm gold particles to 30 nm on average and these could be easily differentiated from non-enhanced 15-nm gold particles. After silver enhancement, labeling for STn was performed employing the same procedure as for MUC2, except that the anti-MRP antibody and GAR-G15 were replaced by TKH2 antibody (1:150) and GAM-G15 (1:40), respectively. After washing in distilled water, the grids were air-dried and counterstained using uranyl acetate and lead citrate. The labeled thin sections were observed under an electron microscope (H-7100; Hitachi).
Labeling controls were done by substituting the primary antibodies with corresponding normal sera from the same species. In addition, omission of the first and secondary antibodies was also included. To verify the specificity of silver enhancement, sections without gold labeling were allowed to react with the enhancement reagent alone. All the control experiments showed negative labeling.
EM ISH Procedure
EM ISH was performed according to published protocols (Morey et al. 1993). Briefly, grids were floated with section face down on Tris-glycine for 15 min. After rinsing in TBS (pH 7.2), the sections were incubated with standard hybridization buffer containing denatured digoxigenin-labeled probe at a final concentration of 40 ng/ml at 37C for 4 hr. Posthybridization washes were on droplets of TBS containing 0.5% Triton and 1% BSA three times for 10 min. Thereafter, the probe detection was done by utilizing a three-step method: (a) incubate with sheep anti-digoxigenin antibody (3 μg/ml) for 2 hr; (b) incubate with rabbit anti-sheep IgG antibody (15 μg/ml) for 45 min; (c) incubate with PA-G15 (1:40) for 1 hr. All incubations were undertaken at RT and an extensive washing in TBS was conducted between each step. After washing in distilled water, the hybridized thin sections were counterstained with uranyl acetate and lead citrate and examined.
The hybridization specificity was controlled as follows: (a) ISH without the RNA probe; (b) ISH with the sense RNA probe; (c) replacement of the anti-digoxigenin antibody with normal sheep serum; (d) treatment of the sections with RNase (50 μg/ml; Amersham Life Science) before initiation of the ISH. These control experiments yielded negative results. In addition, competition using unlabeled pHAM1 DNA probe followed by digoxigenin-labeled pHAM1 DNA probe showed significantly reduced specific binding at the probe throughout the areas showing positive reaction.
Results
Normal Colorectal Mucosa
In normal colorectal mucosa, positive labeling was restricted to goblet cells. MUC2, as visualized by silver-enhanced gold particles (large), was found predominantly in the supranuclear and the perinuclear RER but was also detectable in some RER at the thin peripheral cytoplasm far away from the nucleus and in RER interspersed among the mucin droplets. The Golgi apparatus and mucin droplets were essentially devoid of MUC2 labeling except for occasional sparse background binding. STn, as visualized by non-silver-enhanced gold particles (small), could not be seen in a significant amount in any subcellular structures, such as the RER, the Golgi apparatus, and the mucin droplets (Figure 1).
Colorectal Adenoma
In colorectal adenomas, adenoma cells resembling normal goblet cells could be found and positive labeling was seen in these adenomatous goblet cells. In both low- and high-grade adenomas, MUC2 apomucin was localized predominantly in the RER. In low-grade adenoma, STn expression was observed less often than in high-grade adenoma but, when expressed, it was either only in the trans-Golgi apparatus (Figure 2), or in both the trans-Golgi apparatus and the mucin droplets. In contrast, goblet cell maturation became progressively lost in high-grade adenomas. Although typical mucin droplet aggregates, as found in mature goblet cells, were rarely seen, mucin droplets of various sizes could be found in the cytoplasm of these dysplastic cells. In such high-grade adenomatous cells, STn was expressed more frequently in both the trans-Golgi apparatus and mucin droplets (Figure 3).
Colorectal Adenocarcinoma
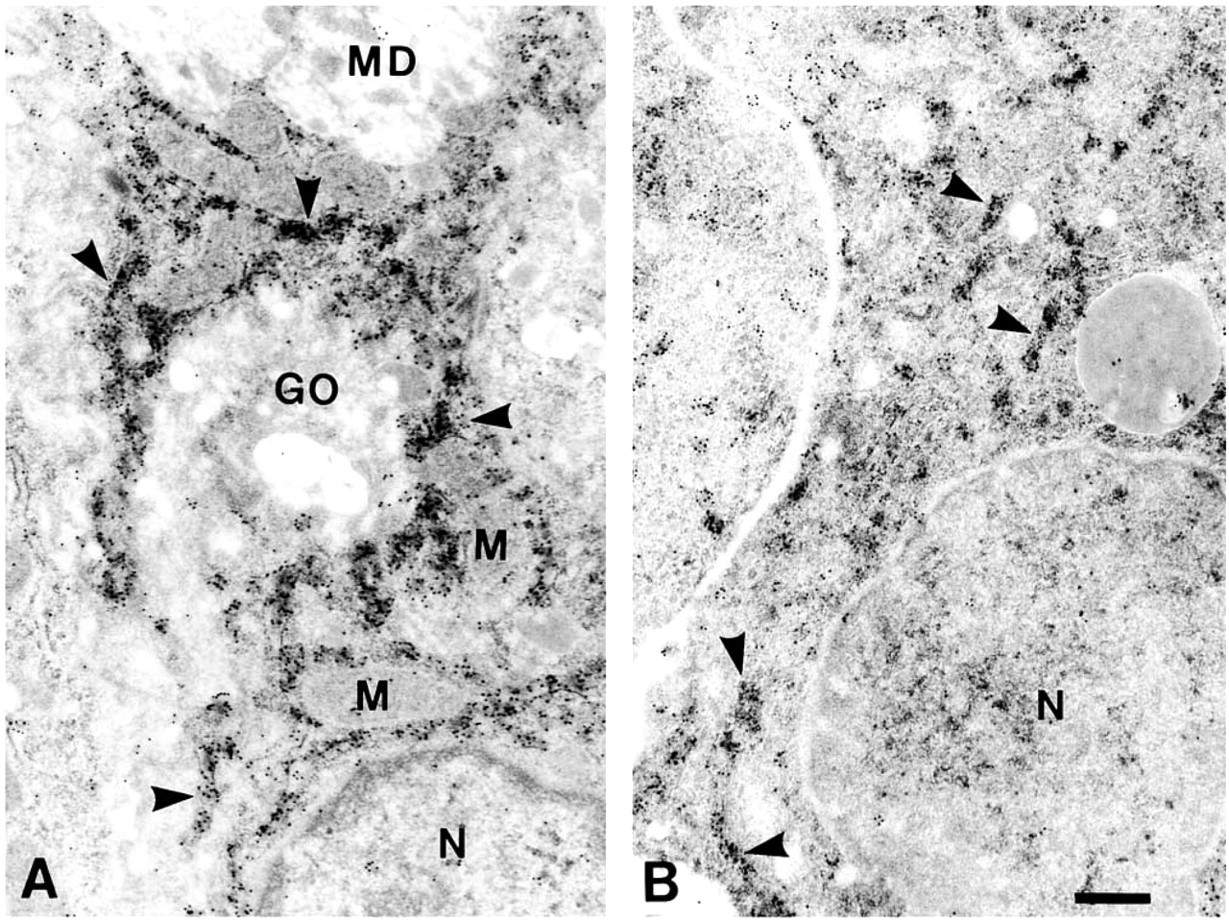
In adenocarcinoma, MUC2-STn double-positive cells were often observed. In these MUC2-expressing cancer cells, STn antigen expression showed several different subcellular localization profiles irrespective of the tumor differentiation level. Extracellularly, STn was often localized in the luminal contents, cell surface membranes, and microvilli (Figure 4). Intracellularly, STn was frequently found in mucin droplets of various sizes and electron densities (Figure 4). Another localization feature was that STn was heavily distributed over the entire Golgi apparatus, more notably in the cis-Golgi and trans-Golgi apparatus (Figure 5). Furthermore, STn was clearly detected in some RER lumens (Figure 6). In all of the above-mentioned situations, MUC2 apomucin was localized predominantly in the RER but could also be observed in some Golgi apparatus (Figure 5). Nevertheless, MUC2 apomucin was not seen in the cell surface or in the microvilli, nor in the glandular luminal contents in any appreciable amount (Figure 4).
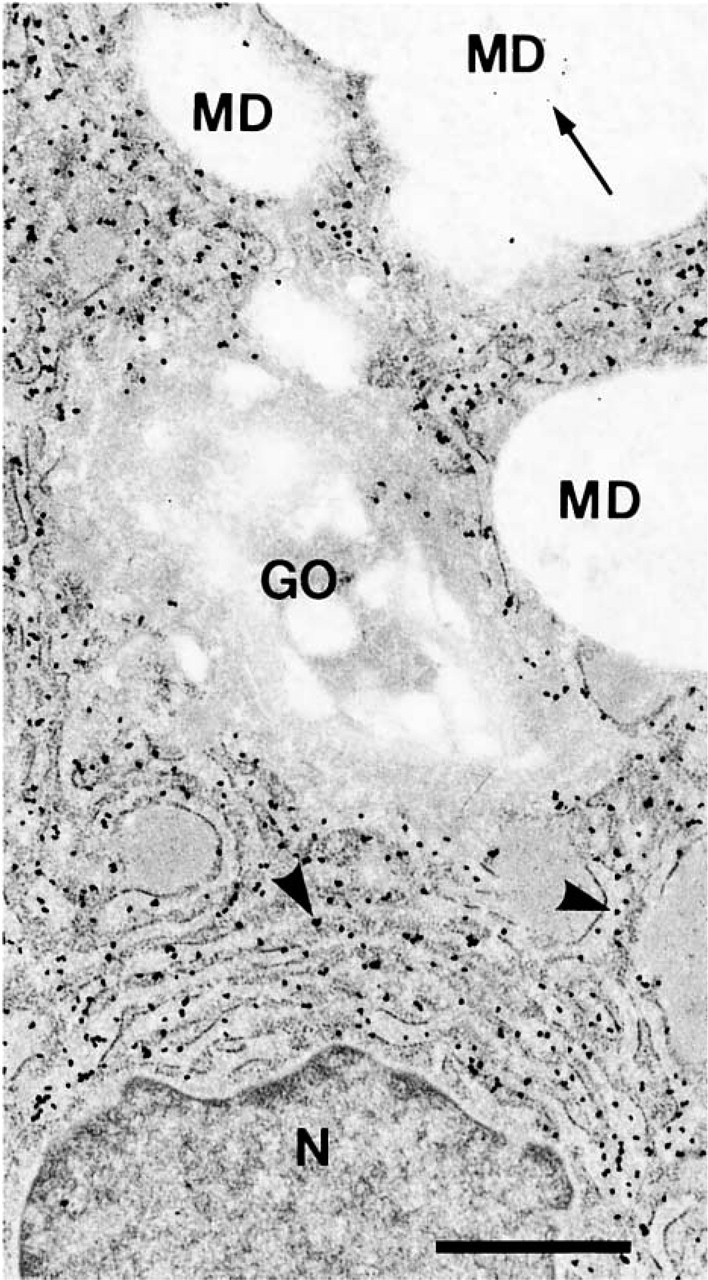
MUC2 apomucin and STn double labeling in normal human colorectal epithelial cells. Shown here is the predominant localization of MUC2 apomucin (arrowheads, large particles) in the RER. Note that only sparse background labeling of STn (arrow, small particles) can be seen in the mucin droplets (MD). The Golgi apparatus (GO) is not labeled either by MUC2 apomucin or by STn. N, nucleus. Bar = 1 μm.
A considerable number of MUC2-negative tumor cells were seen interspersed among MUC2-positive cells. In these MUC2-negative cancer cells, STn was localized in similar structures to those described above, except that the intracellular mucin droplets were usually smaller compared to their counterparts seen in MUC2-positive cells (Figure 4). Malignant goblet cell and columnar cell phenotypes are the two major lineages in colorectal adenocarcinoma. Because MUC2 expression is related to the goblet cell lineage in colorectal cancer (Ajioka et al. 1996), these STn single-positive cells identified in the MUC2-STn double labeling could presumably be of columnar cell lineage.
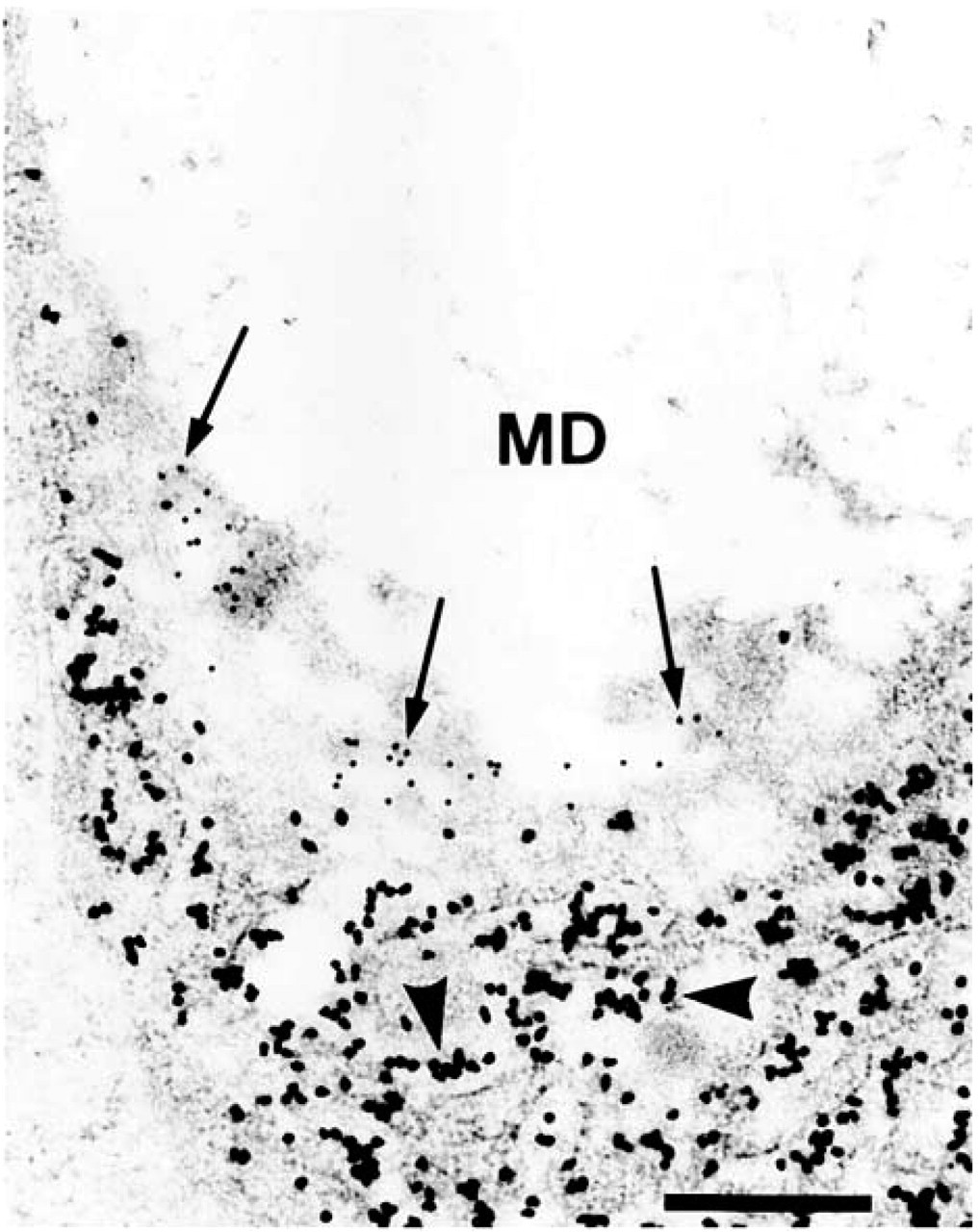
MUC2 apomucin and STn double labeling in human colorectal adenoma. Shown here is the predominant localization of MUC2 apomucin (arrowheads, large particles) in the RER of low-grade adenoma. Note that STn (arrows, small particles) is localized in the trans-Golgi apparatus while the mucin droplets (MD) are essentially devoid of STn labeling. Bar = 0.5 μm.
Although the above-mentioned localization patterns could be seen in all adenocarcinoma cases examined here, MUC2-STn double-positive cancer cells with abundant STn expression in the cell surface and in the RER/entire Golgi compartments were encountered more frequently in adenocarcinoma with lymph node and distant metastasis.
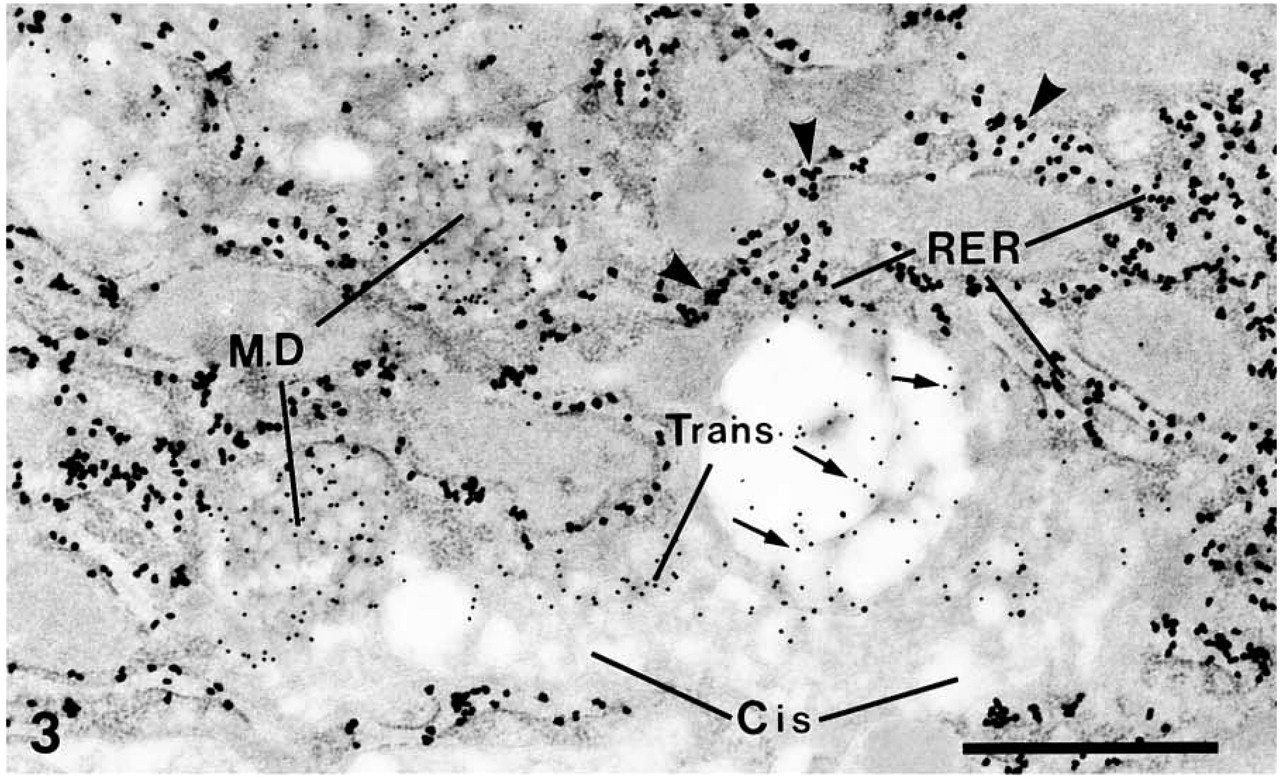
MUC2 apomucin and STn double labeling in human colorectal adenoma. Shown here is the localization of MUC2 apomucin (arrowheads, large particles) in the RER of high-grade adenoma. STn (arrows, small particles) is localized not only in the trans-Golgi apparatus (Trans), but also in the mucin droplets (MD). Note that the cis-Golgi apparatus (Cis) is devoid of STn labeling. Bar = 1 μm.
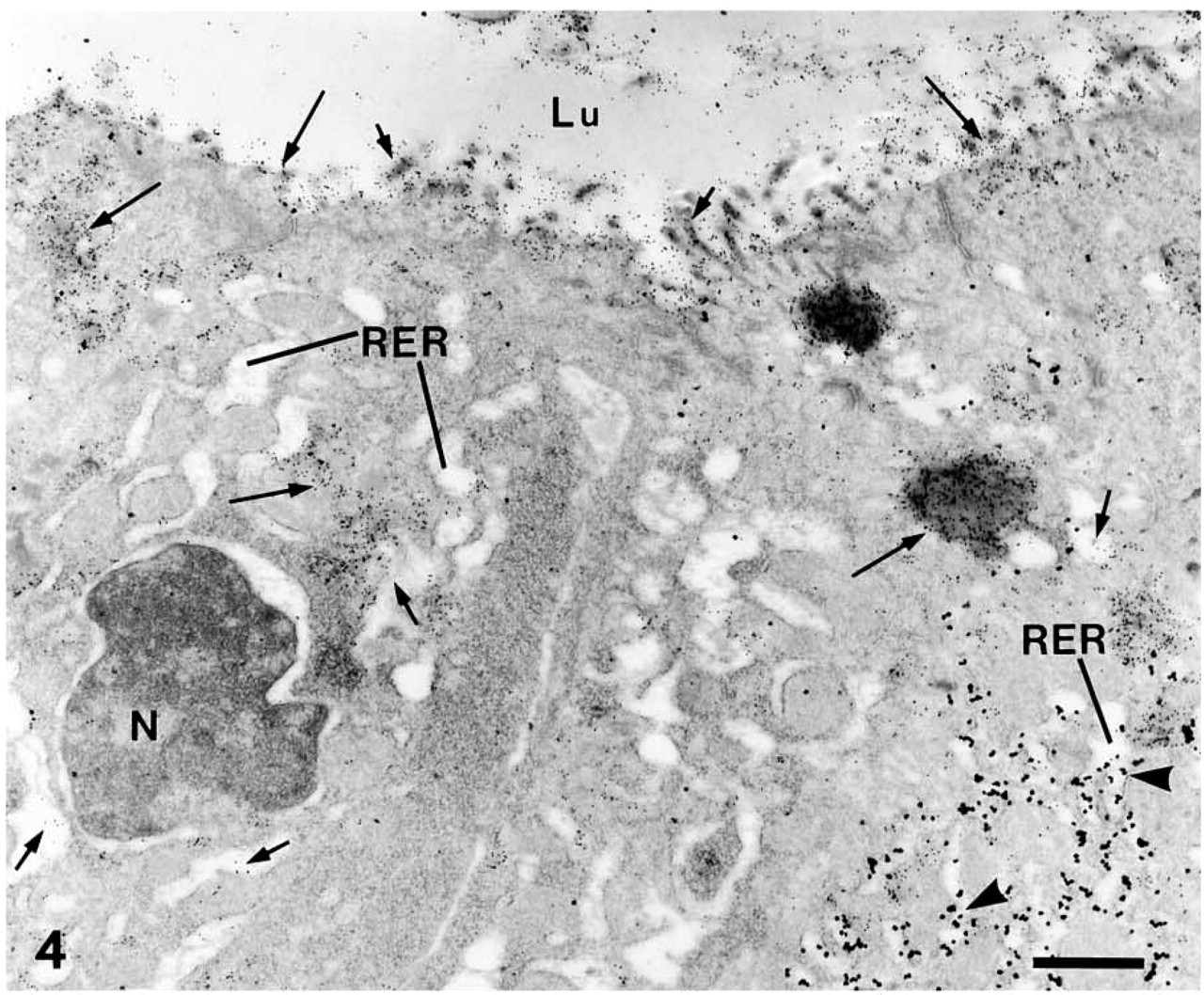
MUC2 apomucin and STn double labeling in human colorectal adenocarcinoma. Shown here are one MUC2-STn double-positive cell (right) and one STn single-positive cell (left). Extracellularly, STn (small particles) is localized in the cell surface (long arrows), microvilli (short arrows) and glandular luminal content (Lu). Intracellularly, STn is seen in mucin droplets (long arrows) and in some RER lumens (short arrows). Note that MUC2 apomucin (arrowhead, large particles) is localized in the RER in the MUC2-expressing cell but not in the cell surface or in the luminal content. N, nucleus. Bar = 1 μm.
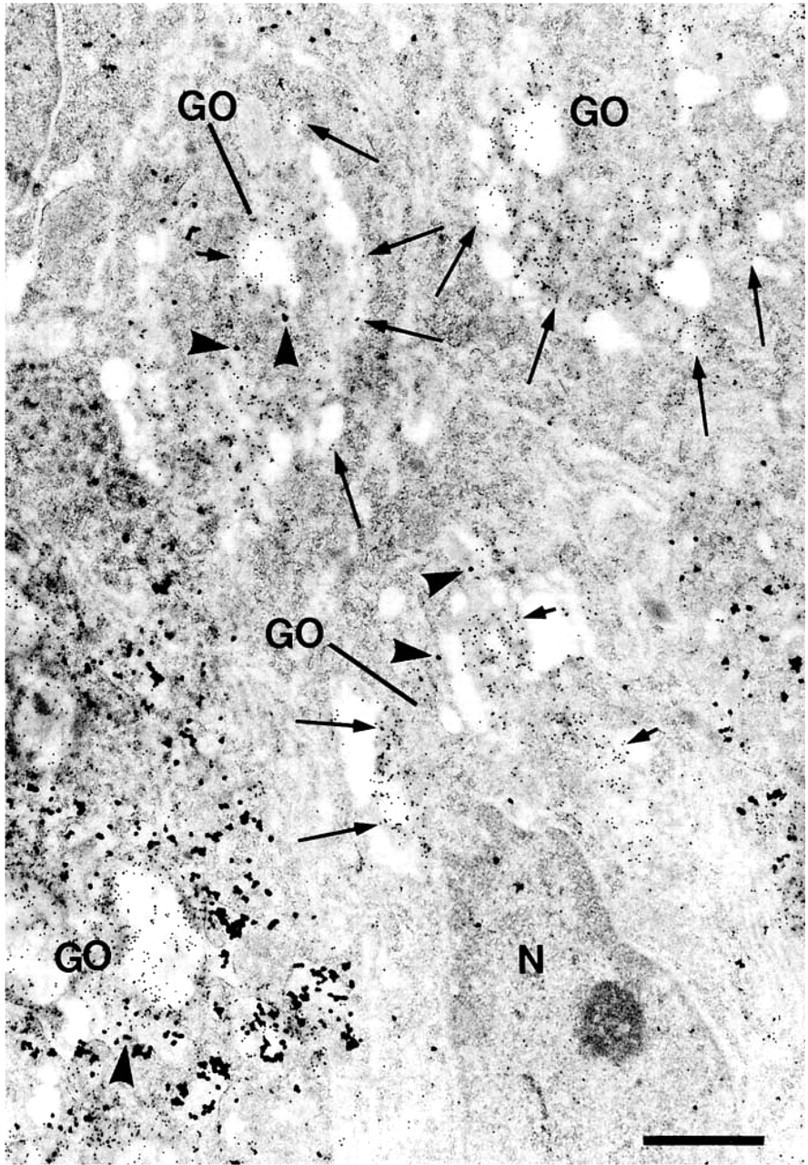
MUC2 apomucin and STn double labeling in human colorectal adenocarcinoma. Shown here is the abundant STn (small particles) localization in the cis- (long arrows) and trans- (short arrows) Golgi apparatus (GO). Note that MUC2 apomucin (arrowheads, large particles) can be seen in the Golgi apparatus. N, nucleus. Bar = 1 μm.
MUC2 mRNA Localization
EM ISH yielded clear hybridization signals without severely compromising the ultrastructural morphology. In all the specimens from normal colorectal mucosa, colorectal adenoma, and colorectal adenocarcinoma examined, gold particles representing MUC2 mRNA were consistently found to be closely associated with the RER (Figure 7). Other fine structures, such as the Golgi apparatus, mucin droplets, and mitochondria were not hybridized to any significant level.
The data on EM subcellular localization patterns of STn, MUC2 apomucin, and MUC2 apomucin mRNA described above are summarized in Table 1.
Discussion
Abnormal sialylation is a glycoconjugate change frequently seen in the neoplastic transformation of epithelial cells in a number of tumors, including colorectal cancer (Ørntoft et al. 1990; Cao et al. 1997). It has been well documented that this aberrant process can produce a number of different sialylated oligosaccharides, such as sialyl LewisA, sialyl LewisX, sialyl T, and STn (Hoff et al. 1989; Hanski et al. 1995; Matsushita et al. 1997). Because STn is involved in a number of important biological properties of cancer cells, such as immunological functions, tumor progression, and metastasis (Ogata et al. 1992; Bresalier et al. 1996; Itzkowitz et al. 1996; MacLean et al. 1996; Sternberg et al. 1996; Imada et al. 1999; Nakagoe et al. 2000), this tumor-associated antigen has attracted particular attention.
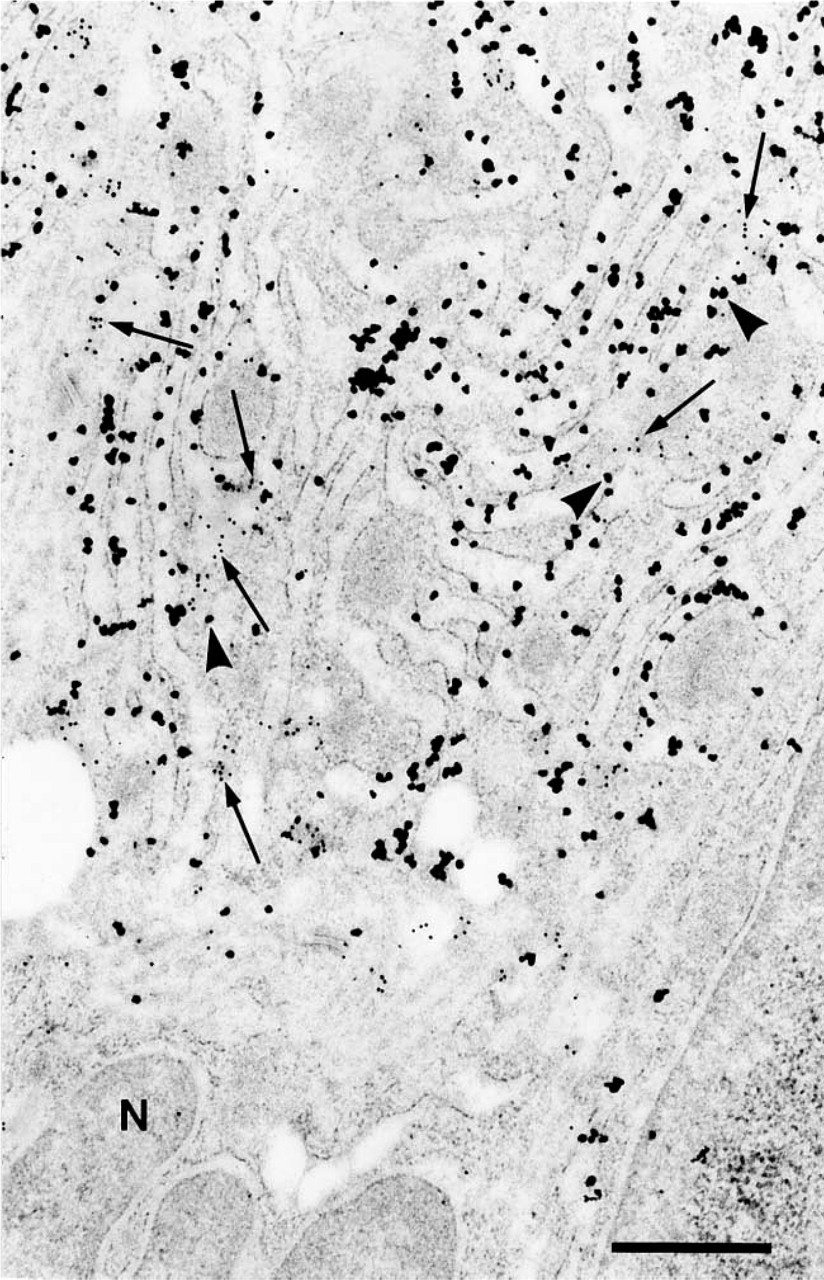
MUC2 apomucin and STn double labeling in human colorectal adenocarcinoma. Shown here is the localization of the STn (arrows, small particles) in the RER. Note that MUC2 apomucin (arrowheads, large particles) is also localized in the RER. N, nucleus. Bar = 0.5 μm.
In this study we were able to examine the subcellular localization patterns of STn in the normal colorectal mucosa, colorectal adenoma, and adenocarcinoma by utilizing the high-resolution IEM labeling method. In agreement with previous STn light microscopic immunohistochemical reports (Itzkowitz et al. 1989; Ørntoft et al. 1990; Yonezawa et al. 1992; Yamachika et al. 1997), STn was not detected in any significant amount in normal goblet cells in the present study. However, distinct STn subcellular localization profiles were observed in both colorectal adenoma and cancer cells in MUC2-STn double-positive cells. Because MUC2 has been regarded as a goblet cell mucin, the colorectal cancer cells that express this mucin are considered to be of goblet cell lineage (Ajioka et al. 1996). In low-grade adenoma, STn was localized in some adenomatous goblet cells either confined to the trans-Golgi apparatus, or in both the trans-Golgi apparatus and mucin droplets. In high-grade adenoma, STn tended to be localized more often in both the trans-Golgi compartment and mucin droplets. In MUC2-expressing colorectal cancer cells, however, the prominent localization features are that STn was found to be distributed throughout the entire Golgi apparatus and localized abundantly in some RER lumens (Figures 5 and 6; Table 1). STn is a mucin-associated disaccharide antigen which resides exclusively on mucin glyco-proteins (Hakomori 1989; Itzkowitz et al. 1992). The biosynthetic pathway for the oligosaccharide side chains of mucin glycoproteins in normal epithelial cells has been shown to progress from RER→Golgi apparatus→secretory granules (Griffths and Simons 1986). Our observation of STn subcellular localization sites along the biosynthetic direction in both colorectal adenoma and adenocarcinoma strongly indicates that, in colorectal adenoma, STn sialylation takes place in the trans-Golgi apparatus whereas in colorectal cancer cells STn sialylation occurs both in the entire Golgi apparatus and in the RER.
EM ISH of MUC2 apomucin mRNA in normal human colorectal mucosa (
In relation to the subcellular localization of the Golgi enzymes in the colorectal cancer, Egea et al. (1993) demonstrated that the enzyme molecules normally resident in the Golgi apparatus could be distributed retrogradely to the RER during malignant transformation of colon epithelial cells. Tn antigen, the precursor of STn, has been detected in the RER of colon cancer cells (Egea et al. 1993). These studies lend support to our observations.
In recent years it has been shown that normal colorectal mucosa constitutively produces STn (Corfield et al. 1995; Jass et al. 1995; Ogata et al. 1995; Klein and Roussel 1998). The negative immunostaining for STn in normal goblet cells has been demonstrated to be due to O-acetylation of sialic acids of STn antigen, which prevents binding by STn-detecting antibodies such as TKH2 (Ogata et al. 1995) used in this study. Because O-acetylated STn could not be detected by TKH2, the present data do not enable us to draw direct conclusions about the STn sialylation compartment in normal colorectal goblet cells. Nevertheless, a number of studies (Roth 1984; Roth et al. 1994; Whitehouse et al. 1997) have demonstrated that Tn is formed in the cis-Golgi apparatus in normal cells. Therefore, it is likely that in normal mucosal cells, no sialylation of any O-glycans will happen before the cis-Golgi compartment. In addition, the trans-Golgi apparatus has been found to be the subcellular compartment for O-acetylation of sialic acids (Butor et al. 1993; Chammas et al. 1996; Vandamme-Feldhaus and Schauer 1998). Because no significant STn localization was seen either in the trans-Golgi or the post-Golgi elements in the normal colorectal cells, it seems reasonable to speculate that, in normal colorectal tissue, STn sialylation and O-acetylation take place in the same subcellular compartment, i.e., the trans-Golgi apparatus, although it should be emphasized that further studies on the immunolocalization of the specific O-acetyltransferase for STn are required to verify this hypothesis.
Expression patterns of STn, MUC2, and MUC2 mRNA in human colorectal tissues a
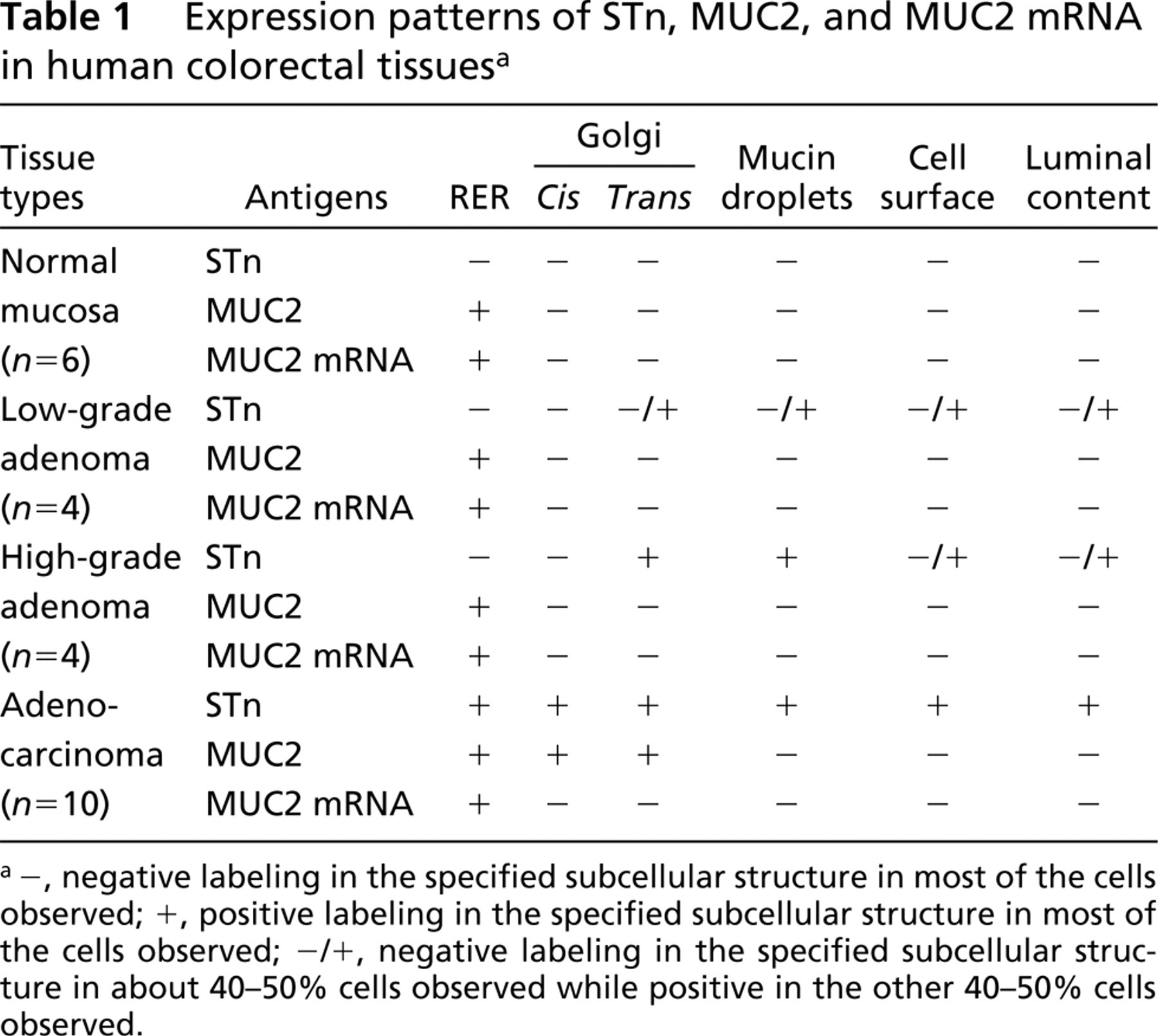
-, negative labeling in the specified subcellular structure in most of the cells observed; +, positive labeling in the specified subcellular structure in most of the cells observed; -/+, negative labeling in the specified subcellular structure in about 40–50% cells observed while positive in the other 40–50% cells observed.
In human colorectal tissue, MUC2 has been found to be a major mucin type (Kim 1993; Tytgat et al. 1994). Recent studies of Sternberg et al. (1996) have further confirmed that MUC2 serves as a major carrier for STn, although STn can be borne by other mucin proteins as well (Cao et al. 1997; Matsushita et al. 1997). Although the EM ISH profile of MUC2 mRNA expression shows that MUC2 apomucin is synthesized in the RER in both normal and malignant colorectal tissues, our subcellular immunolocalization study indicates that, unlike sole RER localization in the normal colorectal cells, MUC2 apomucin is expressed in both the RER and the Golgi apparatus in neoplastic colorectal cells. This finding strongly suggests that defective and sparse glycosylation of MUC2 apomucin, resulting in the exposure of apomucin peptide and/or altered compartmentalization of STn sialylation, constitutes the basis for the aberrant STn and MUC2 apomucin expression patterns observed in neoplastic colorectal epithelial cells.
In animal models, human colon cancer cells selected for high metastatic potential produce high levels of MUC2 mucin (Bresalier et al. 1996). Antisense inhibition of MUC2 mucin synthesis resulted in almost complete loss of STn expression and dramatically reduced liver colonization by human colon cancer cells (Sternberg et al. 1996). In the present study, we found that colorectal cancer with lymph node and distant metastasis showed more MUC2-STn co-localization, with STn being abundantly expressed in the cell surface and in the RER/entire Golgi compartments. This appears to suggest that neoplastic MUC2 mucin and MUC2-borne STn might play a role in clinical colorectal cancer metastasis.
Methodologically, studies on the subcellular glycosylation compartments are usually conducted through localization of the glycosylated products or the enzyme proteins of corresponding glycosyltransferases (Roth 1984; Roth et al. 1994; Chammas et al. 1996). Due to unavailability of a detection reagent specific for STn sialyltransferase, we have not been able to conduct IEM localization of this enzyme. However, as demonstrated recently by Chammas et al. (1996), EM localization of the glycosylated product provides valuable information on elucidation of the subcellular glycosylation compartments because glycosylation is subject to control by a number of complex factors, such as availability of the substrate, acceptor, and endogenous transporter. Studies by Hayes et al. (1993) indicate that the presence of the enzyme per se does not necessarily define a functional subcellular compartment. In a detailed study, Chammas and colleagues (1996) have further shown that the distribution of a sialyltransferase with lactose-4 MU specificity has broader distribution than its sialylated product. In addition, some glycosyltransferases may simply act like lectins, capturing the substrate without glycosylation (Ma and Colley 1996; Ma et al. 1997). Therefore, examination of the subcellular localization of glycosylated products provides important information on biosynthetic pathways of mucin glycosylation in normal and neoplastic cells.
Footnotes
Acknowledgments
Supported in part by Grants-in-Aid from the Ministry of Education, Science, Sports and Culture, Japan (12218233, 12218234, 12470046, and 13220016) and also by a USPHS Grant from the National Cancer Institute (CA24321), the Department of Veterans Affairs Medical Research Service, and the Theodore Betz Foundation (USA).