
Abstract
Mechanisms involved in skeletal myofiber differentiation during fetal development of large animals are poorly understood. Studies in small animals suggest that the calcineurin (Cn) pathway is involved in myofiber differentiation. Neural activity is a prerequisite for Cn activity, implying maintenance of sustained low intracellular Ca2+ concentrations. To study the role of Cn in fetal myofiber differentiation, we monitored the temporal and spatial distribution of Cn subunits, sarcoplasmic reticulum Ca2+ ATPase (SERCA), phospholamban (PLB), and myosin heavy chain (MyHC) isoforms in relation to ingrowing nerves in porcine semitendinosus muscle (m. semitendinosus) at 55 and 75 days of gestation (dg) and at term. Immunofluorescence analysis revealed the presence of Cn subunits and SERCA isoforms at all analyzed stages. Cn distribution was not fiber-type specific, but expression became more prominent at term. At 75 dg, differential SERCA2 expression was accompanied by perinuclear PLB in primary fibers. SERCA1 was expressed in all fiber types at all stages. No specific MyHC isoform distribution was seen in relation to neuromuscular contacts, although neuromuscular contacts were present. From these results we speculate that in porcine m. semitendinosus differential SERCA2 expression precedes differential Cn expression. The question whether the Cn pathway is involved in prenatal myofiber differentiation needs further studies.
I
In pig, primary myotubes form between 35 and 55 days of gestation (dg), followed by the second generation between 55 and 95 dg, and the third generation around birth (Lefaucheur et al. 1995; Picard et al. 2002). During gestation, not only a temporal differentiation takes place but also a spatial differentiation, ultimately leading to a unique architecture. The mature pig skeletal muscle shows a highly organized distribution pattern of fiber types mostly consisting of islets of slow-type I fibers surrounded by an internal ring of fast-type IIA and an external ring of fast-type IIB fibers (Lefaucheur et al. 1995).
The signals and the mechanisms of differentiation playing a role in early myogenesis of the primary fiber generation have recently been elucidated (Buckingham 2001,2003; Christ and Brand-Saberi 2002). Also, the evolution of fiber-type differentiation in developing muscles (of the pig) as well as postnatal muscles has been described (da Costa et al. 2002; Lefaucheur et al. 2002; Picard et al. 2002). However, the mechanisms underlying the differentiation of the second and third generation of myofibers during fetal development remain poorly understood. For mice, regulation of sarcoplasmic reticulum gene expression during skeletal muscle development has been described (Arai et al. 1992). In addition, the signaling pathway of calcineurin (Cn) has been implicated in controlling slow- vs fast-fiber phenotypes as described for mouse, cell cultures (primary myoblasts, C2C12 myogenic cells), and for fiber-type switching after regeneration in rat (Chin et al. 1998; Friday et al. 2000; Buckingham 2001; McCullagh et al. 2004; Schulz and Yutzey 2004).
Two mechanisms have been implicated in the control of the activity of Cn. First, tonic motor nerve activity, characteristic of nerves innervating slow muscles, has been described to sustain [Ca2+]i at levels sufficient to activate Cn via activation of Ca2+-sensitive calmodulin. It has been shown that Cn activation selectively establishes the slow muscle phenotype, including expression of slow MyHC and slow sarcoplasmic reticulum ATP-ase type 2 (SERCA2) (Bigard et al. 2000; Schulz and Yutzey 2004). Second, as Cn is a heterodimeric protein phosphatase composed of a catalytic subunit A and a regulatory subunit B (Rusnak and Mertz 2000), regulation of Cn activity has been proposed through regulation of CnB protein levels (Mitchell and Pavlath 2002).
A role of the calcineurin pathway in myofiber differentiation during fetal development of large animal muscles has not yet been explored. Myofibers in large mammals such as the pig do not run from tendon to tendon and show multiple innervation zones (Paul 2001). As Cn activation requires electrical nerve activity providing the sustained [Ca2+]I, we postulate that this pathway is involved in the achievement of the highly organized patterns of distributed fiber types. We therefore chose to perform this investigation on porcine muscle starting at 55 dg when the formation of the second generation of myofibers had started. Formation of secondary myotubes initiates at the innervation sites of the primary myotubes (Duxson and Sheard 1995), supporting a role of neurogenic control in secondary myotube differentiation. We have chosen the semitendinosus muscle (m. semitendinosus), which has multiple innervation zones and a characteristic rosette myofiber-type distribution. In an earlier study we showed that the porcine m. semitendinosus is already fully innervated at 44 dg when only primary myotubes have developed (De Jonge et al. 2005). From this work, however, we cannot deduce if the neuromuscular contacts are already functional. In the underlying study we have investigated a possible influence of ingrowing nerves and/or neuromuscular contacts on spatial and temporal myofiber differentiation. We monitored at several ages of gestation the distribution of a) expression of MyHC myofiber isoforms and b) the potential regulatory factors of myofiber differentiation: Cn catalytic unit A and regulatory unit B, potential regulators of [Ca2+]i SERCA1 and 2 (committed, respectively, to fast- and slow-type myofibers) and phospholamban (PLB, regulator of SERCA2 activity). The distributions are monitored in relation to axonal outgrowth, indicated by the presence of growth-associated protein 43 (GAP43) and neuromuscular contacts, indicated by the presence of acetylcholine receptor (AChR) subunits α and γ. The γ subunit of the ACh receptor is specific for fetal motor nerve endings, whereas the α subunit is present in both pre- and postnatal isoforms of the mammalian receptor (Mishina et al. 1986; Takahashi et al. 2002).
Materials and Methods
Animals
Fetal pigs were collected from slaughtered sows (the standard Dutch commercial hybrid race) in a commercial slaughterhouse. Fetal age was estimated from the crown-rump length of the litter, using the growth curves of Evans and Sack (1973). We obtained litters from the prenatal ages of 55 and 75 days postconception, at term, and 112-116 days postconception. Experiments were approved by the Animal Ethical Committee for Animal Experimentation of Utrecht University.
Tissue Collection
Directly after death of the fetuses, the hind leg attached to the hemisected pelvis was fixed in toto by immersion in phosphate-buffered 4% formaldehyde for 24 hr at 4C. Dermal incisions of the oldest fetuses increased the exposure to fixation fluid. Thereafter, tissues were stored in 70% alcohol at 4C until further preparation.
Vibratome Sections and Free-floating Immunoassay
The semitendinosus muscle was dissected from the hind legs, embedded in 30% gelatin, and subsequently cut longitudinally on the vibratome (Micro-cut H1200; Bio-Rad, Hercules, CA) in 80-μm sections. For any gestational age, the left and right muscles were taken from two to three fetuses for immunoanalysis. Sections were analyzed by double- or triplefluorescent labeling in free-floating immunoassays. Briefly, sections were rinsed in PBS, permeabilized in 100% methanol for 10 min at −20C, rinsed in PBS, blocked in 0.1% BSA (fraction V, BDH, UK) in PBS, and incubated with a mixture of first antibodies in 0.1% BSA in PBS for 1 hr at room temperature followed by 40 hr at 4C. The sections were then rinsed in large volumes of PBS, followed by the incubation with the mixture of secondary fluorescent conjugated antibodies in 0.1% BSA in PBS for 1.5 hr at room temperature. Finally, the sections were rinsed in large volumes of PBS, followed with a quick rinse in dH2O, mounted in Fluorsave Reagent (Calbiochem; EMD Biosciences, Inc., Darmstadt, Germany) on SuperFrost Plus (Menzel-Glazer; Braunschweig, Germany) glass slides and left to dry at 37C.
To control the performance of the resulting assay, for example, in background staining in every free-floating immunoassay, all incubations were paralleled by negative controls treated similarly with the exception of the first antibody.
Antibodies
Sections were stained using the following first antibodies: anti-skeletal muscle slow MyHC (NOQ7.5.4D), mouse monoclonal IgG, dilution 1:2000 and anti-skeletal fast MyHC (MY-32), mouse monoclonal IgG, dilution 1:2000 (both from Sigma-Aldrich; StLouis, MO); anti-SERCA1 ATPase (IIH11), mouse monoclonal IgG, dilution 1:500, and anti-SERCA2 ATPase (IID8), mouse monoclonal IgG, dilution 1:500 (both from Abcam; Cambridge, UK); anti-calcineurin A (PP2B-A, clone H-209), rabbit polyclonal IgG, dilution 1:100 (Santa Cruz Biotechnology, Santa Cruz, CA), anti-calcineurin B, rabbit polyclonal, dilution 1:100 (Affinity BioReagents; Golden, CO); anti-AChRα2 (H-111), rabbit polyclonal IgG, dilution 1:100 and anti-AChRγ (H-172), rabbit polyclonal IgG, dilution 1:100 (both from Santa Cruz Biotechnology; tebubio b.v., The Netherlands); anti-GAP43 (H-100), rabbit polyclonal IgG, dilution 1:100 (Santa Cruz Biotechnology). Anti-phospholamban (clone A1), mouse monoclonal IgG, dilution 1:100, was purchased from Upstate (Lake Placid, NY). Secondary monoclonal antibodies used were goat anti-mouse or goat anti-rabbit, highly cross-adsorbed whole antibody conjugates Alexa Fluor 488 or 568 at a dilution of 1:100, and nuclear staining was performed with TO-PRO-3 iodide, all purchased from Molecular Probes (Eugene, OR).
Analysis
Vibratome sections were analyzed on a Leica TCS Sp (Wetzlar, Germany) confocal laser scanning microscope using laser lines 488, 568, and 633 nm, and images shown are composed of these sequential scannings. Negative controls were set on the background settings of the positive controls taken on the confocal scanning microscope.
Results
MyHC Expression and Neuromuscular Contacts
We monitored the spatial distribution of neuronspecific proteins in relation to the fiber-type pattern in the porcine fetal m. semitendinosus. To study the presence of the neuromuscular junctions, antibodies against ACh-receptor subunits α and γ were used. Figure 1 illustrates the expression pattern of ACh-receptor subunit α (Figures 1A-1C and γ (Figures 1D-1F) in combination with the expression of slow MyHC (Figures 1A,1C-1F) or fast MyHC (Figure 1B) at three different stages of development. Both receptor type units were already present at 55 dg.
The presence of positive staining for GAP43 at all presented gestation ages (Figures 1G-1I) confirmed our earlier findings that the porcine m. semitendinosus was already fully innervated as early as 44 dg with a finemeshed neural network supplying the whole muscle (De Jonge et al. 2005). In the latter study we applied the classical silver staining method to visualize the ingrowing fetal nerves, which revealed the same patterns found by immunohistochemistry presented in the present study. In addition, staining fetal m. semitendinosus with Alexa 488-conjugated α-bungarotoxin presented similar patterns (data not shown). Figures 1G-1I illustrate the combination of GAP 43 with the expression of slow MyHC (Figures 1H and 1I) and fast MyHC (Figures 1B and 1G). We could not reveal a specific expression pattern directly related to the presence of neuromuscular junctions or ingrowing nerves at any of the analyzed gestational ages.
Expression of SERCA and PLB
Activity of the calcineurin pathway is determined by [Ca2+]I, which in turn is controlled by many proteins located in the sarcoplasmic membrane. One of these proteins is PLB-regulating SERCA. Differences in expression patterns of SERCA1 and 2 and PLB were found (Figure 2 and Figure 3). Because antibodies used against slow-type MyHC proteins and SERCA were both raised in mouse, we tried to use another antibody against slow MyHC, also raised in mouse but of type IgM (A4.840; Alexis Biochemicals, Lausen, Switzerland). Unfortunately, this antibody did not give any positive staining, making it impossible to achieve double staining of slow MyHC and SERCA. Using CnA (red) as a counterstaining, we could detect SERCA1 expressed in sarcomeric banding patterns in all fibers at 55 and 75 dg (Figures 2A and 2B). At term (Figure 2C), SERCA1 is still expressed at the sarcomeres as well as in clusters throughout all myofibers, as can clearly be observed in Figures 3A and 3B. SERCA2 expression was irregular at 55 dg (Figure 2D). At 75 dg it was restricted to large fibers, assumed to be primary fibers, in banding patterns but also concentrated around the nucleus (Figure 2E). At term, SERCA2 expression was located near the sarcomeres of large, central fibers (Figure 2F). In Figures 3D-3F, cross-sections of m. semitendinosus at term are presented, clearly showing the expression of SERCA2 in central fibers. The expression of the SERCA2 regulator PLB was similar to SERCA2 expressed in the large central fibers. This was first seen at 75 dg (Figure 2H). Distribution of PLB within each fiber was remarkable. As revealed by the use of a nuclear staining with TO-PRO-3 iodide (Figures 2H ∗ and 2I ∗), it was solely concentrated at the periphery of the nucleus. At 55 dg, PLB expression was undetectable.
Calcineurin Expression
We have analyzed the expression of both CnA and B at different gestational ages. Both subunits were expressed in all fibers at all ages (Figures 4A-4F), and both subunits became more prominent at term (Figures 4C and 4F). At 55 dg, CnA expression is irregular (Figure 2G and Figure 3A), whereas at 75 dg it is more ubiquitous; at term, expression of CnA is found to be in small concentrates clearly seen in Figure 2I and Figures 3A, 3C, 3D, and 3F. It is, however, difficult to discern whether it is colocalized or localized in between sarcomeric proteins (Figure 4C). CnB at 55 dg is expressed more diffusely, although the image obtained with the 488-nm laser line revealed expression near the sarcomeres (Figure 4D). This diffuse expression of CnB was also seen at 75 dg (Figure 4E), whereas at term CnB expression seemed to reveal a banding pattern (Figure 4F).
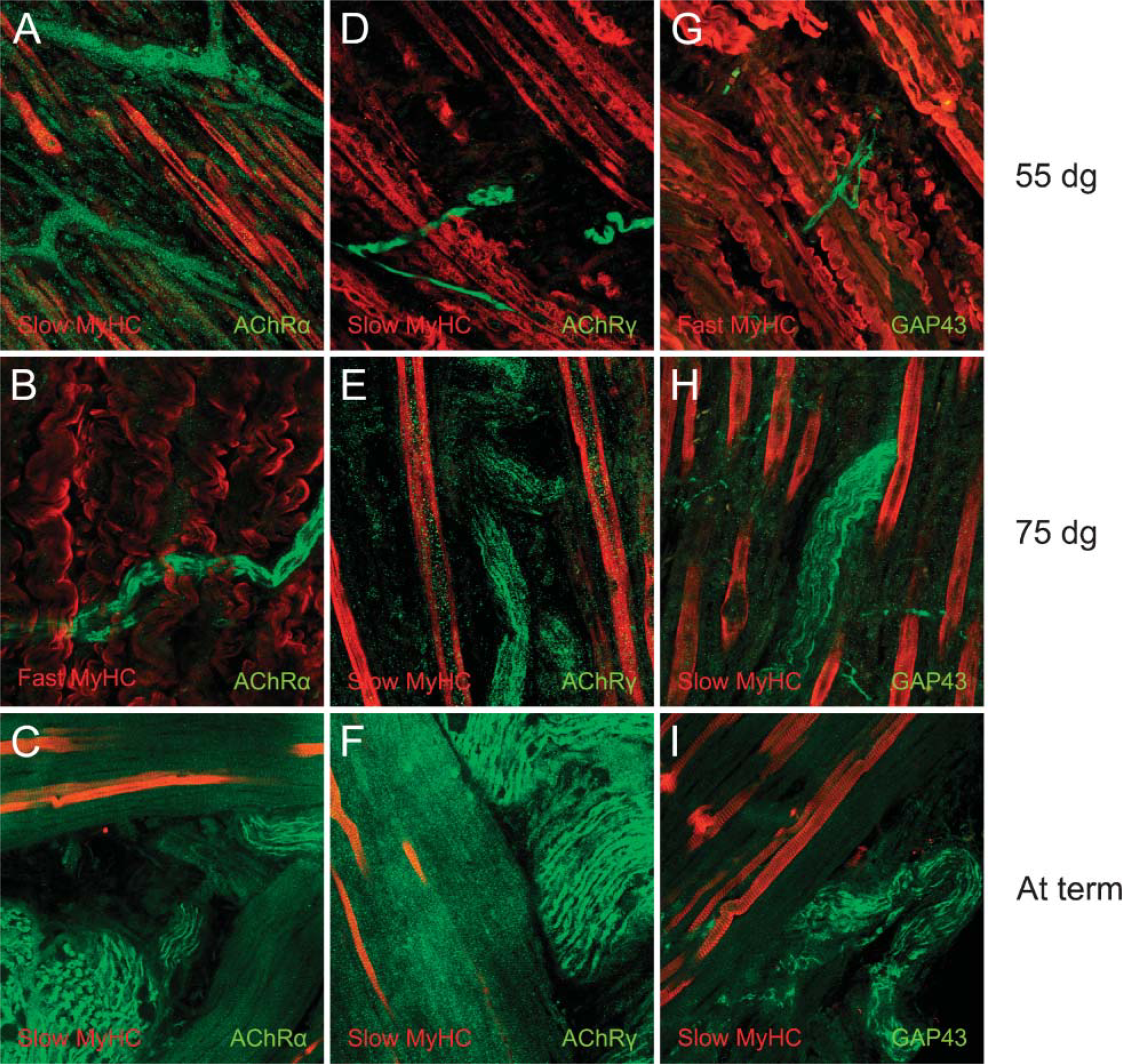
Immunoanalysis of the expression of myosin heavy chain (MyHC) isoforms slow or fast in relation to the presence of acetylcholine receptor (AchR) subunits α or γ or growth-associated protein 43 (GAP43) at 55 or 75 dg or at term in the porcine semitendinosus muscle (m. semitendinosus). In green: AChR subunits α (
Discussion
The general process of myogenesis is similar in different species. However, the kinetics of muscle fiber development appears to be different among species depending on their maturity at birth (Picard et al. 2002). For small laboratory animals it has been described that skeletal muscles are highly undifferentiated at birth and then undergo a marked process of differentiation in attaining the adult phenotype (Baldwin and Haddad 2001), whereas fiber-type differentiation in large farm-animal species has its onset before birth (Picard et al. 2002). The phenotypic differentiation leading to the mature fiber identity is influenced by innervation. The neural initiation site on the primary myofibers correlates with the onset of secondary myofiber formation (Gunning and Hardeman 1991; Zhang and McLennan 1998).
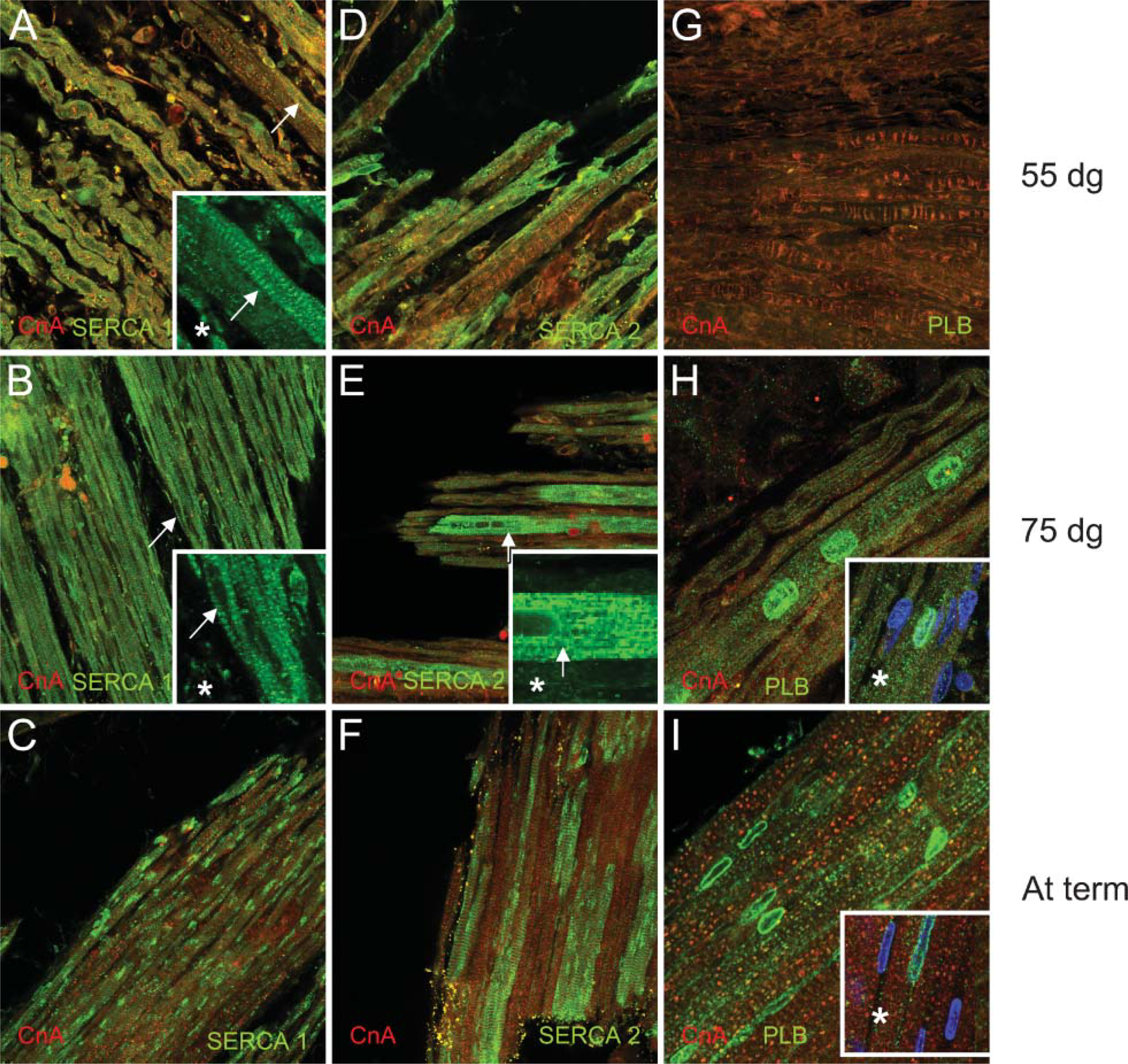
Immunoanalysis of the expression of sarcoplasmic reticulum Ca2+ ATPase 1(SERCA1), SERCA2, or phospholamban (PLB) in the porcine m. semitendinosus at 55 or 75 dg or at term. In green: SERCA1 at 55 dg (
MyHC Expression
In large animals such as the pig, results on myofiber typing of fetal and adult muscle based on histochemistry and immunohistochemistry using non-commercial antibodies may differ (Lefaucheur et al. 2002) due to the lack of reliable antibodies specific for pig, as well as the presence of a high proportion of hybrid fibers (Quiroz-Rothe and Rivero 2004; Toniolo et al. 2004). Lack of specific antibodies against fast MyHC isoforms seriously hampered our study. Although we did not find clear differences in distribution of slow or fast MyHC in the area of neuromuscular contacts, we cannot rule out a specific distribution of the several fast MyHC isoforms. Because we were interested in the spatial distribution of myofiber types and other factors, we used relatively thick formalin-fixed vibratome sections and free-floating immunoassays, further complicating specific antibody binding.
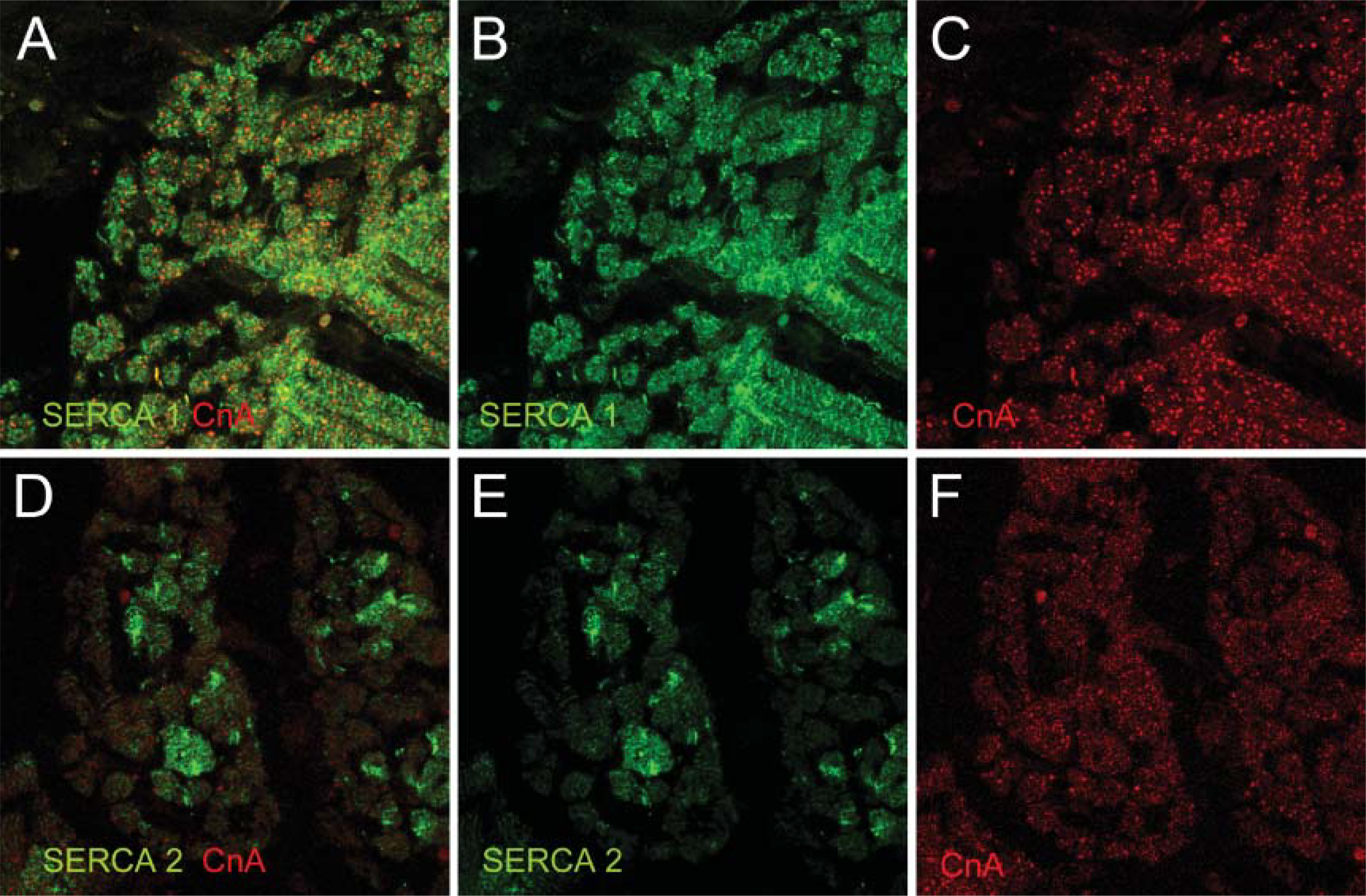
Immunoanalysis of the expression of SERCA1 (green,
SERCA and PLB Expression
In fetal porcine m. semitendinosus, SERCA1 appears in all fiber types as seen in the cross-sections at term, although a reduced expression at primary fibers cannot be ruled out with certainty because of the impossibility of double staining with anti-type-I antibody. At term, SERCA1 expression in condensates, as well as in a banding pattern near the sarcomeres, may imply reorganization in quantity or localization of protein expression. Expression of SERCA2 in conjunction with PLB in assumed primary fibers is supported by the study of Quiroz-Rothe and Rivero (2004). These authors found a nearly unique correspondence in the distribution of sarcoplasmic reticulum-related proteins, such as slow isoforms of SERCA and slow MyHC, PLB, and slow MyHC but also fast MyHC and fast SERCA isoforms in adult porcine m. longissimus lumborum, which is similar to findings reported for fetal mice (Arai et al. 1992). In fetal porcine m. semitendinosus, PLB at 75 dg was convincingly found to be sublocalized at the periphery of the nucleus possibly colocalized with SERCA2 as also SERCA2 density was increased around the nucleus (Figures 2E and 2H). This peculiar expression pattern was consistent and was found in several specimens at 75 dg, as well as at term. Recently, it has become clear that nuclear calcium signals are also regulated at the nuclear envelope and nuclear reticulum (Echevarria et al. 2003) by, e.g., SERCA-type Ca-ATPase (Bachs et al. 1994; Zhao et al. 2004). Because it is generally accepted that PLB is coexpressed with the SERCA2 gene in slow-twitch skeletal muscle, PLB may be (temporally) expressed in the nuclear envelope in a similar manner, e.g., X,K-ATPase β-subunit of which a unique temporospatial distribution in the nuclear envelopes of perinatal myocytes has been described (Zhao et al. 2004).
Calcineurin Expression
In rabbit soleus muscle, CnB was immunocytochemically found to be expressed in a diffuse way in both fast and slow fiber types and to be arranged in cross-striated bands in functioning muscles (Sacchetto et al. 2001). Our findings on Cn subunit distribution in porcine m. semitendinosus corroborate these findings. In developing muscle, CnB revealed a similar pattern of diffused expression but in a cross-striated pattern at term. CnA expression pattern in concentrates, as we found in fetal porcine muscle, was not described as such by Sacchetto et al. (2001). However, Torgan and Daniels (2006) showed a similar expression pattern in cultured rat myotubes. They showed that calcineurin staining became more prominent in mature myotubes and that it began to exhibit a cytoplasmic pattern that included rows of punctua to the developing myofibrils, supposedly on either side of the Z-line in a pattern consistent with that of triads (Torgan and Daniels 2006). Differences in expression pattern or quantity may serve a regulatory role. An unequal distribution of the Cn subunits over different muscles leads to differences in activity levels, as has been described for the plantaris and soleus muscle in mouse (Mitchell et al. 2002).
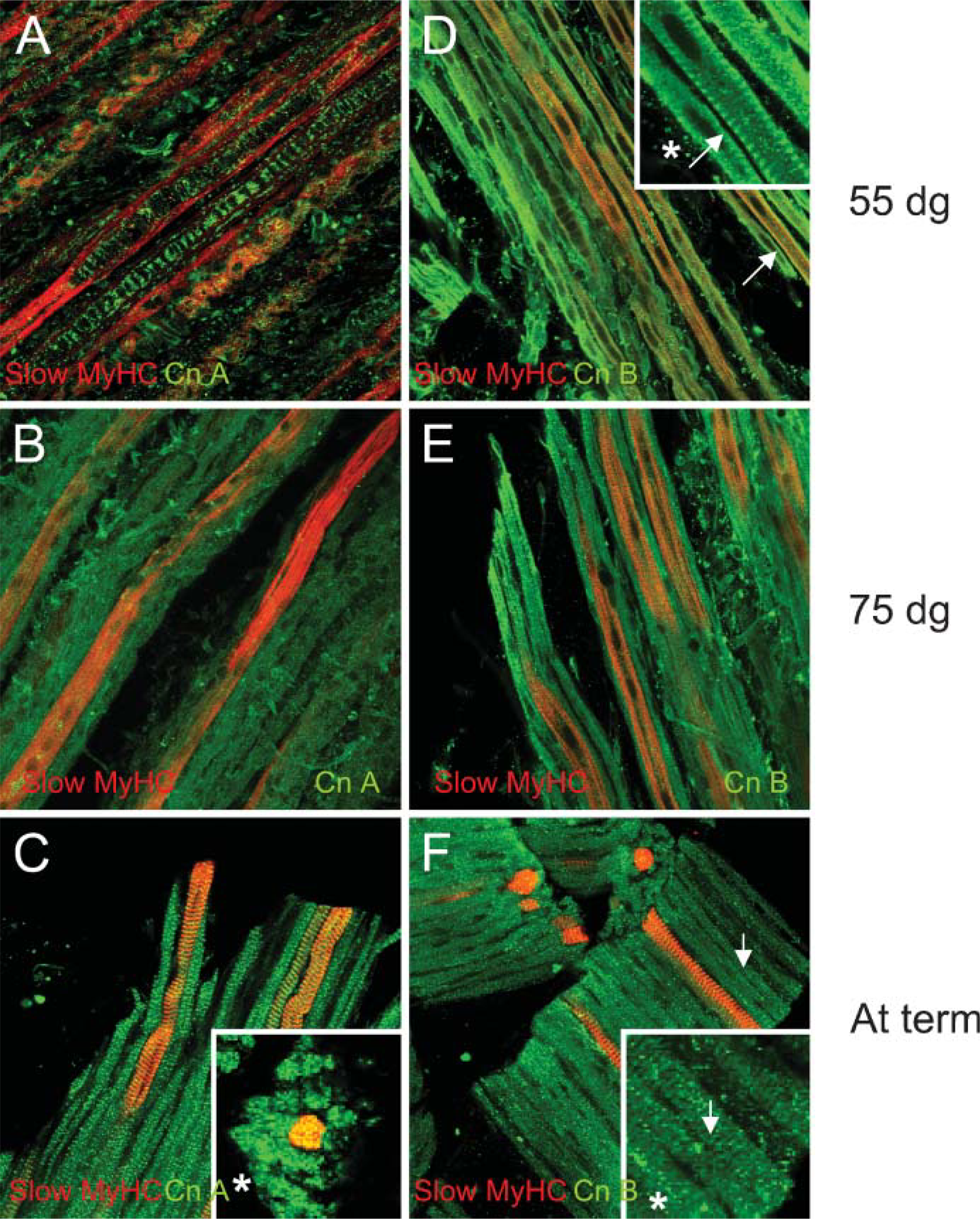
Immunoanalysis of the expression of subunits CnA and CnB in the porcine m. semitendinosus at 55 dg, 75 dg, or at term. In green: CnA at 55 dg (
In developing porcine m. semitendinosus, distribution of the Cn subunits is not associated with the spatial or temporal distribution of the [Ca2+]i regulator SERCA. In the pig, the temporal distribution of SERCA runs, in turn, parallel to myofiber type differentiation, e.g., concerning MyHC isoform expression, which starts at ∼70-75 dg (Picard et al. 2002). Cn activity is not only responsible for myofiber differentiation but is also implicated in maintenance of muscle calcium homeostasis (Mitchell et al. 2002), which may be the prenatal role for Cn. The maintenance of the appropriate [Ca2+]i in slow fibers by SERCA2 and PLB may, therefore, be a prerequisite for maintenance of the slow (primary) fiber type. According to the fiber-type differentiation evolution scheme given by Picard et al. (2002), although starting at 70-75 dg, the major differentiation of porcine secondary fibers occurs postnatally. As the kinetics of myofiber development is dependent on the maturity at birth, we may be able to explain our results in pig based on our laboratory findings in horse. Elevated levels of CnA and CnB were associated with SERCA2 expression in slow MyHC fibers in the equine m. gluteus medius at 28 days before birth (Eizema K, unpublished data). In pig, at birth less mature than horse, we found that CnA is clearly present, and CnB reveals a banding pattern. However, the distribution is not slow-fiber-type specific. A contribution of Cn in porcine secondary myofiber differentiation might be postponed and occur postnatally.
In summary, to study fetal myofiber differentiation, we have analyzed the temporal and spatial distribution of MyHC isoforms, Cn, and SERCA 1/2 and PLB in developing porcine m. semitendinosus in relation to neuromuscular contacts. Although we could not distinguish a specific MyHC isoform distribution, from 75 dg onward, SERCA2 was specifically expressed in large, presumably primary fibers, concomitant with a peculiar but consistent expression of PLB. SERCA1 was expressed in all fibers. Cn subunit expression was analyzed, and both subunits appeared more prominent at term. We speculate that in correspondence to evolution of porcine myofiber differentiation, in the m. semitendinosus, calcineurin may be involved in myofiber differentiation after birth.
Footnotes
Acknowledgements
We thank Ing. Anko de Graaff and Dr. Richard Wubbolts of the Center for Cell Imaging (Faculty of Veterinary Medicine, Utrecht University) for technical advice.