
Abstract
The accurate classification of skeletal muscle fiber types according to myosin heavy chain (MyHC) polymorphism remains a difficult task in the pig. Combined myofibrillar ATPase and metabolic enzyme histochemistry, in situ hybridization, and immunocytochemistry were performed on serial transverse sections of pig longissimus (L) and rhomboideus (R) muscles at 100 kg body weight to give a new insight into muscle fiber typing in the pig. Several monoclonal antibodies (MAbs) either specific for a single MyHC (I, IIa, or IIb) or of multiple MyHCs (IIa + IIx or I + IIx + IIb) were used. No monospecific IIx antibody was available for the pig. All three adult Type II isoforms were expressed in the white L muscle, whereas no IIb was observed in the red R muscle, which was confirmed using RNase protection analysis. In most fibers, the distribution of the transcripts closely matched that of the corresponding proteins. When observed, co-expression of MyHCs mostly occured for IIx and IIb in L muscle, and was more common at the protein (11.5%) than at the mRNA (2.2%) level. A minor proportion of myofibers showed a mismatch between MyHC mRNA and protein. According to the type grouping distribution of myofibers encountered in pig muscle, MyHC isoform expression followed the rank order of I→Iia→Iix→IIb from the center to the periphery of the islets, concomitantly with a decrease in oxidative metabolism and an increase in fiber size. The developmental origin and functional significance of the type grouping distribution are discussed.
Keywords
S
Materials and Methods
Muscle Samples
A female Large White pig from the INRA (Institut National de la Recherche Agronomique) herd was slaughtered at the usual commercial body weight of 100 kg (170 days of age) by electrical stunning and exsanguination, in compliance with French national regulations applied in commercial slaughtering. Immediately after slaughter, two muscles with different locations, functions, and biochemical properties were sampled: the longissimus (L) at the last rib level, a fast-twitch glycolytic muscle involved in voluntary movements of the back, and the tubular portion of the rhomboideus (R), a postural mixed-oxydo-glycolytic muscle involved in supporting the head. Samples were mounted on flat sticks, frozen in isopentane cooled by liquid nitrogen, and stored at — 80C until further analysis.
Histochemistry, In Situ Hybridization, and Immunocytochemistry
Histochemistry, ISH, and ICC were performed on 10-μm-thick serial transverse sections cut on a cryostat at —20C (Reichert-Jung, 2800 frigocut N; Heidelberg, Germany). A section was stained using conventional mATPase histochemistry after preincubation at pH 4.35 to identify types I, IIA and IIB fibers (Brooke and Kaiser 1970). A serial section was processed for succino-dehydrogenase (SDH) to classify fibers according to their oxidative metabolism (Nachlas et al. 1957). In situ hybridizations were performed as described by Lefaucheur et al. (1998), with the following modifications. Sections were treated with 10 μg/ml proteinase K instead of 5 μg/ml. The hybridization buffer contained 60% deionized formamide and 0.36 M NaCl instead of 50% and 0.3 M, respectively. Finally, overnight hybridization temperatures were set at 40C for Types I and IIa, and 30C for Types IIx and IIb riboprobes. Exposition to Kodak NTB-2 nuclear track emulsion ranged from 10 days for the Type I probe to 30 days for the IIx probe. After photographic development, slides were analyzed using darkfield and phase-contrast optics with a Leitz Diaplan microscope (Wetzlar, Germany). Immunohistochemistry was performed on serial 10-μm-thick sections as previously described (Lefaucheur et al. 1995). Briefly, the sections were incubated overnight at 4C with the primary MAb and the specific binding was revealed by the avidin-biotin-peroxidase technique (Vectastain ABC kit; Vector Laboratories, Burlingame, CA). For each muscle, fiber type composition was measured from three fields containing a total of about 1200 fibers using tracing paper to combine information from the serial sections. Mean cross-sectional area (CSAs) of each fiber type was estimated from about 150 fibers of each type using a programmable planimeter (Hitachi; Tokyo, Japan).
Riboprobes for In Situ Hybridization
Probes against the different MyHCs were subcloned in either pBluescript KS+ (Stratagene; La Jolla, CA) or pCR 2.1 (Invitrogen; Carlsbad, CA) vectors. Vectors were linearized with appropriate restriction enzymes. Thereafter, the cRNA probes were transcribed using either T3 or T7 RNA polymerases according to the manufacturer's instructions (Promega; Madison, WI) using [35S]-UTP as the labeled nucleotide (New England Nuclear; Boston, MA). The Type I MyHC probe was a 136-base pair (bp) fragment including the last 19 bp of the translated region (TR) and the entire 3′-untranslated region (3′-UTR) (Figure 1A). The IIa MyHC probe was a 134-bp fragment comprising the full-length 3′-UTR (Figure 1B). The IIx and IIb MyHC probes correspond to the 3′ half portion of the 3′-UTR and are 43 and 53 bp fragments, respectively (Figure 1B).
Monoclonal Antibodies
Different MyHC MAbs were tested: NLC-MHCs (Novocastra; Newcastle, UK), reactive with pig slow-twitch Type I MyHC (Lefaucheur et al. 1995); SC-71, specific for Type IIa MyHC in the rat (Schiaffino et al. 1989); BF-F3, reactive with Type IIb MyHC in the rat (Schiaffino et al. 1989); and 6B8, specific for Type IIa MyHC in pig muscle (Depreux et al. 2000). Finally, two new MAbs, S5–7D4 and S5–8H2, produced by Biocytex (Marseille, France) in collaboration with INRA, were tested; they have been shown to specifically recognize MyHC proteins by Western blots in different species (Picard et al. 1998), and their specificity for each MyHC was further analyzed in the present study by ICC in the pig. Immunocytochemistry was not complemented by Western blotting for the following reasons: (a) no high-resolution SDS gel technique is available to separate IIa, IIx, and IIb isoforms in pig muscle, in contrast to what is observed in mouse, rat, and rabbit (Talmadge and Roy 1993; Janmot and d'Albis 1994; Agbulut et al. 1996); and (b) no skeletal muscle is pure in Type I, IIa, IIx, or IIb MyHC in the pig. All antibodies but 6B8 were hybridoma supernatants diluted 1:10 before use. Antibody 6B8 was produced from ascites and diluted 1:25,000.
RNA Isolation and RNase Protection Analysis
Total RNA was isolated from the L and R muscles using the guanidium thiocyanate method (Chomczynski and Sacchi, 1987). RNA was quantified by absorbance at 260 nm and its integrity was determined by examining the 28S and 18S rRNA bands on ethidium bromide-stained agarose gels. A single antisense riboprobe was used to quantify the three adult fast-twitch Type II (IIa, IIx, IIb) mRNAs in the same sample of total RNA using RNase protection analysis (RPA). The template was a 397-bp pig Type IIx MyHC cDNA corresponding to nucleotides 2684–3080 (GenBank, AB025262). This sequence was 100% identical to the pig IIa (GenBank, AB025260) and IIb (GenBank, AB025261) over a length of 338 and 265 bp, respectively. The homology with the sequence of the pig Type I MyHC (GenBank, U75316) was only 80% and should not give a protected fragment larger than 23 bp. The cDNA was cloned into the vector pCR2.1 (Invitrogen; Groningen, The Netherlands). The antisense riboprobe was transcribed with a Promega kit using T7 polymerase in the presence of [α32P]-CTP (New England Nuclear). The RPA protocol was as described by Combes et al. (1997) after optimization for the pig IIx ribo-probe. In particular, 900,000 cpm of Type IIx MyHC ribo-probe was used to ensure that the probe was at least in fivefold molar excess. Ten μg of total RNA was hybridized overnight at 45C in a buffer containing 40 mM PIPES (pH 6.4), 1 mM EDTA, 0.2 M sodium acetate, and 80% deionized formamide. Samples were digested at 37C for 1 hr with a 1:2500 dilution of an RNAse cocktail composed of 750 U/ml RNase A and 20,000 U/ml RNase T1 (Ambion; Austin, TX). The sizes of the protected fragments were determined by comparison with [32P]-RNA fragments of 500 to 100 bp (Ambion). The radioactivity of the protected fragments was detected with a PhosphorImager (Storm; Molecular Dynamics, Sunnyvale, CA) and quantified using Im-ageQuant (Molecular Dynamics). Data were corrected to take into account the difference in the amount of [a32P]-CTP incorporated into each protected fragment. The relative amounts of Type IIa, IIx, and IIb mRNAs were expressed as a percentage of total Type II transcripts.

Nucleotide sequence of the 3′ ends of pig Type I, IIa, IIx and IIb myosin heavy chains (MyHCs). Stop codons and polyadenylation sites are singly and doubly underlined, respectively. The nucleotide sequences corresponding to the riboprobes are in bold. (
Statistical Analysis
Influence of fiber type and muscle on cross-sectional area was analyzed by a two-way ANOVA using the GLM procedure of SAS (1990). The model included the main effects of fiber type and muscle as well as their interactions. Duncan's multiple range tests were used for comparison of the means when main effects were significant.
Results
Histochemistry
The mATPase staining after acidic preincubation (pH 4.35) allowed a distinction between black Type I, unstained Type IIA, and gray Type IIB fibers in both L and R muscles (Figure 2). A type grouping distribution of fiber types was observed, consisting of islets of Type I fibers in the vicinity of which Type IIA and then Type IIB fibers are found. The percentages of Type I, IIA, and IIB fibers were 11%, 7%, and 82% in L muscle, and 68%, 12%, and 20% in R muscle. According to the SDH staining, fibers were classified as weakly, moderately, or highly oxidative (Figure 3B, fibers 3, 4 and 5, respectively). Whereas all Type I and IIA fibers were highly oxidative in both muscles, IIB fibers exhibited a heterogeneous SDH staining. Thus, IIB fibers were either weakly (fiber 3) or moderately (fiber 4) stained in L muscle (Figures 3A and 3B) and were always moderately stained (fiber 3) in R muscle (Figures 4A and 4B). About 20% and 80% of conventional IIB fibers were moderately and weakly oxidative in the L muscle, respectively.
In Situ Hybridization
The presence of positive and negative fibers within the same sample allowed a clear identification of the specific signals. Serial sections processed for ISH with cRNA probes specific for each adult MyHC isoform showed that fibers 1, 2, 3, and 4 were exclusively stained with Type I, IIa, IIb, and IIx probes in L muscle (Figures 3C–3F), showing that cRNA probes were highly specific for each MyHC isoform. An estimation carried out on 600 fibers in the L muscle showed that very few of them co-expressed two MyHC transcripts (I + IIa, 0.1%; IIa + IIx, 0.3%; IIx + IIb, 2.2%). In the L muscle, the percentages of fibers expressing Type I, IIa, IIx, and IIb transcripts were 10.9%, 6.7%, 17.5%, and 62.3%, respectively (Figure 5). In R muscle, no IIb transcript was observed (Figure 4F) and the proportions of fibers expressing Type I, IIa, and IIx mRNA were 68%, 12%, and 20%, respectively (Figure 5). Very few hybrid fibers were detected in this muscle. Based on the expression of MyHC mRNA, intensity of SDH staining was high for Types I and IIa fibers, moderate or high for Type IIx fibers, and low for Type IIb fibers (Figure 5). Influence of fiber type on myofiber CSA was different between the two muscles (Figure 6). CSA increased in the rank order of I = IIa < IIx < IIb in L muscle, and I < IIa = IIx in R muscle. A comparison between muscles showed that Types I and IIa were larger in R muscle (p<0.0001), whereas Type IIx fibers exhibited similar CSA in both muscles.
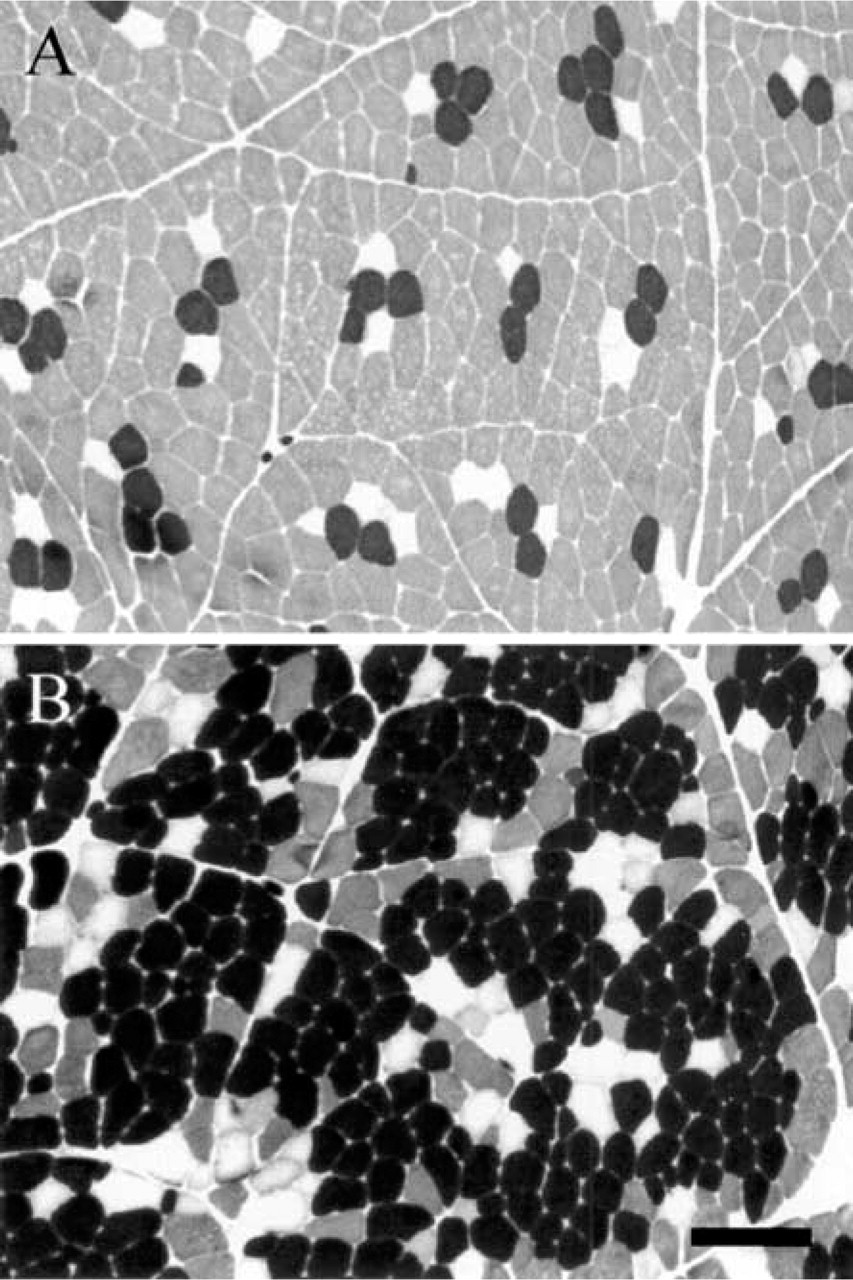
mATPase histochemistry after preincubation at pH 4.35 in longissimus (
RNase Protection Analysis
As expected, three protected fragments of 397, 338 and 265 bp, corresponding to Type IIx, IIa, and IIb mRNAs, were observed by RPA (Figure 7). All three Type II transcripts were present in the L muscle, whereas only the IIa and IIx transcripts were visible in the R muscle. A quantitative analysis of the Type II MyHCs showed that the proportions of IIa, IIx, and IIb mRNAs expressed as a percentage of total Type II transcripts were 7%, 41%, and 52% in the L muscle and 32%, 67%, and 0% in the R muscle, respectively.
Immunocytochemistry
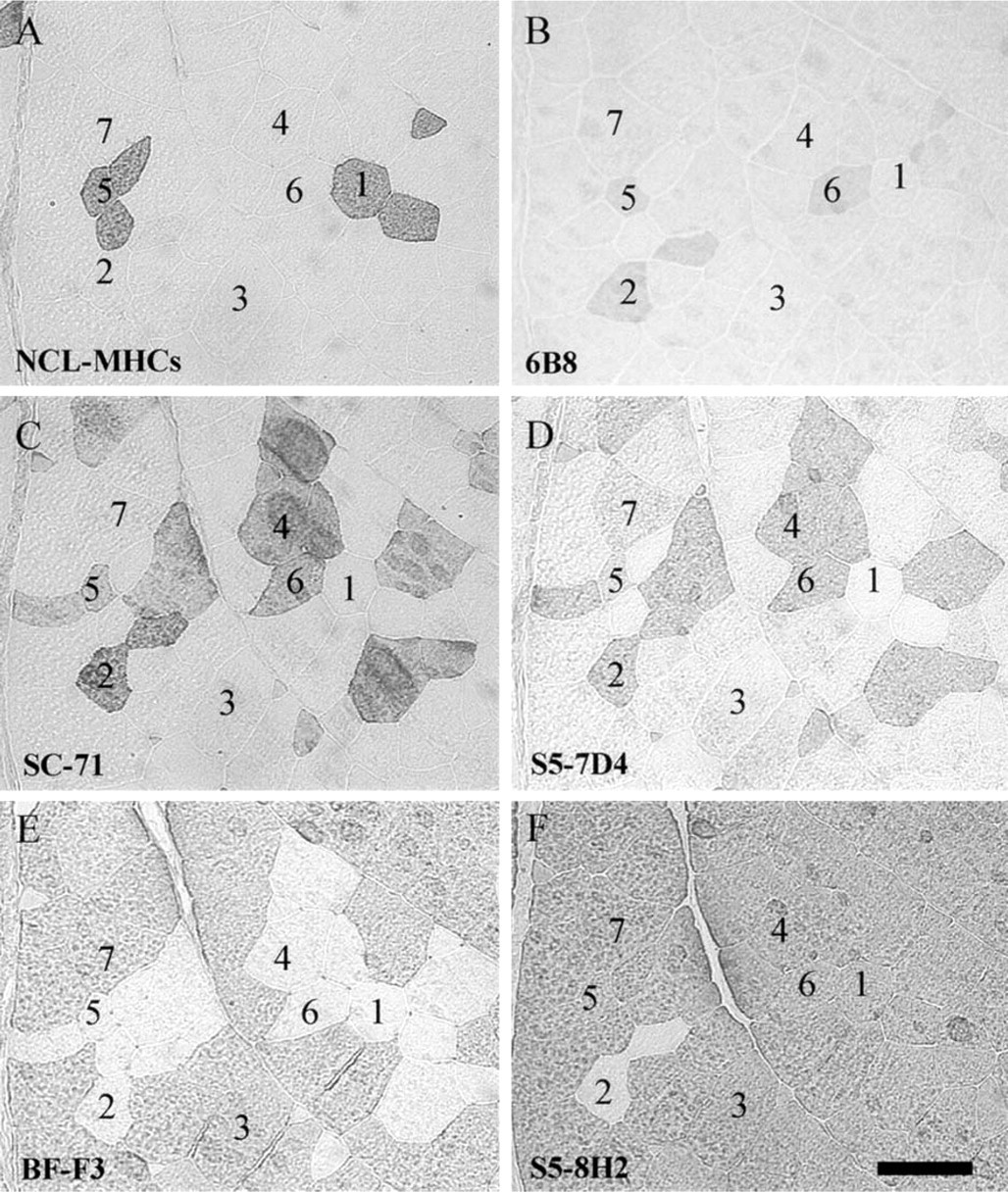
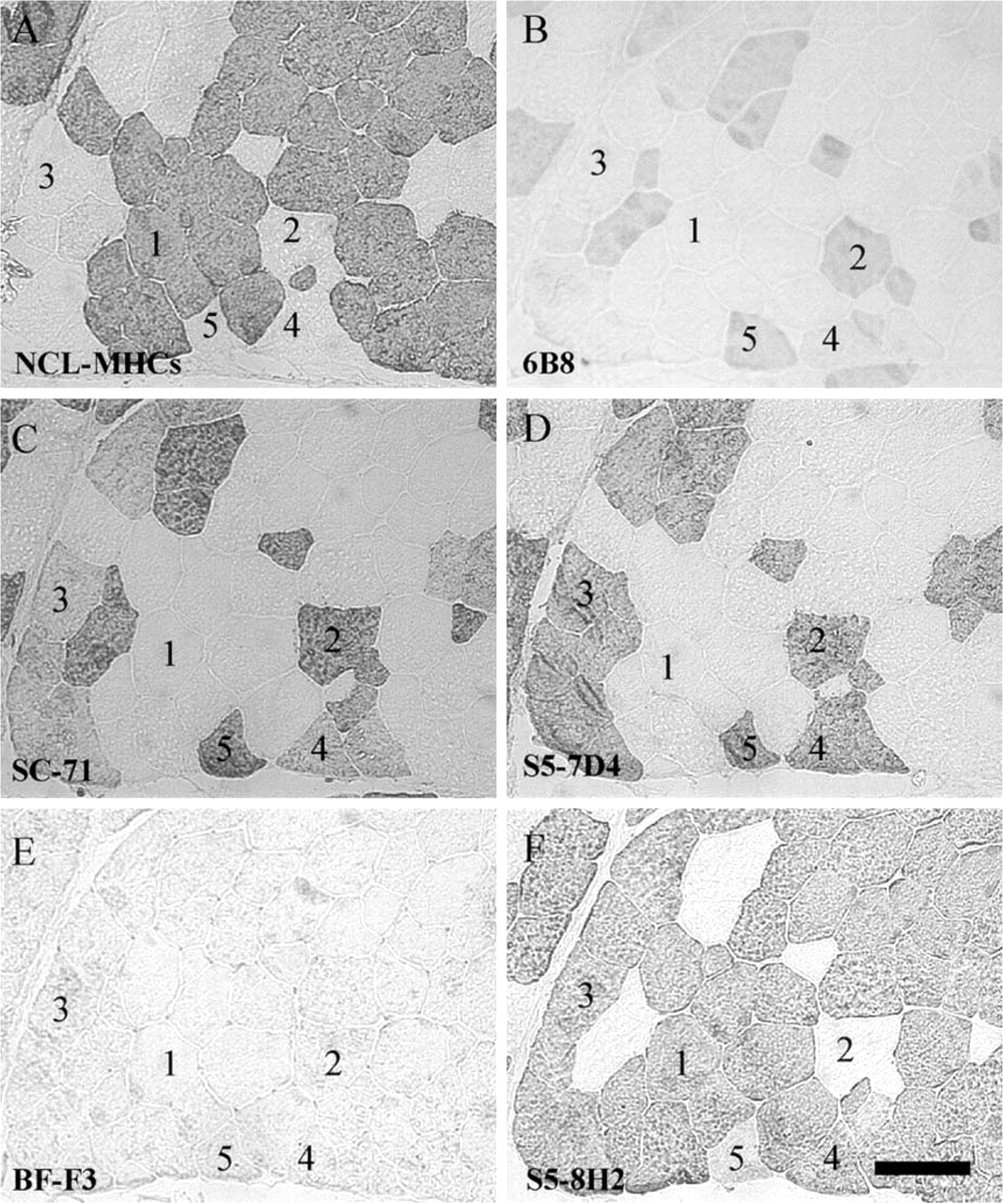
NLC-MHCs MAb. In both muscles, Type I fibers, as evidenced by the mATPase staining (Figures 3A and 4A), were reactive with NLC-MHCs, whereas remaining fibers were not (Figures 8A and 9A). We conclude that NLC-MHCs is specific for Type I MyHC in the pig. At the mRNA level, all Type I fibers were labeled with the Type I cRNA probe (Figures 3C and 4C).
6B8 MAb. All unstained fibers with the mATPase staining (Type IIA fibers) were reactive with 6B8 antibody in both muscles (Figures 8B and 9B) and were labeled with the IIa cRNA probe (Figures 3D and 4D). Most remaining fibers were unstained with 6B8 and contained either Type I, IIx, or IIb mRNA. Therefore, the present data identified 6B8 as an antibody specific for IIa MyHC in pig muscle. Interestingly, additional fibers that were stained either in black (Type I) or gray (Type IIB) after the mATPase staining were also reactive with the 6B8 antibody (Figure 8B, fibers 5 and 6; Figure 9B, fiber 5). Because these fibers were unreactive with the IIa cRNA probe (Figures 3D and 4D), they contained the IIa protein without the corresponding mRNA. At the protein level, fiber 5 in L muscle is a hybrid fiber (I + IIa) which is converting to a Type I fiber (IIa→I), as suggested by the presence of only the Type I mRNA (Figure 3C). Fibers 6 in L muscle (Figure 3A) and 5 in R muscle (Figure 4A) are hybrid fibers containing IIa + IIx proteins and are in the process of being converted to a Type IIx fiber (IIa→IIx), as suggested by the presence of only Type IIx mRNA (Figures 3E and 4E).
SC-71 MAb. In the L muscle, SC-71 recognized Type IIA and a subpopulation of histochemically defined Type IIB fibers (Figure 8C, fiber 4) that were moderately oxidative (Figure 3B). These IIB fibers were positive with the IIx riboprobe (Figure 3E) and negative with the IIb riboprobe (Figure 3F). In the R muscle, this antibody recognized all Type IIA and IIB fibers (Figure 9C). All conventional Type IIB fibers (fiber 3) were reactive with the IIx probe (Figure 4E) and unreactive with the IIb probe (Figure 4F) in the R muscle. Therefore, our results show that SC-71 recognized both IIa and IIx MyHC in pig skeletal muscle, and that labeled conventional IIB fibers are in fact IIx fibers. It is noteworthy that Type IIA fibers (Figure 9C, fiber 2) were more intensely labeled with SC-71 than conventional Type IIB fibers (Figure 9C, fiber 3), suggesting that the affinity of SC-71 was higher for IIa than for IIx MyHC.

Identification of myofiber types by histochemistry and distribution of the different myosin heavy chain (MyHC) mRNA in longissimus muscle. Histochemical demonstration of mATPase after preincubation at pH 4.35 (
BF-F3 MAb. In the L muscle, BF-F3 recognized all histochemically defined Type IIB fibers that were unreactive with SC-71 (Figure 8E, fiber 3). These fibers were strongly reactive with the IIb riboprobe (Figure 3F), whereas they were unstained with the IIx probe (Figure 3E). In the R muscle, all conventional IIB fibers (fiber 3) were negative with the BF-F3 antibody (Figure 9E) and the IIb probe (Figure 4F), whereas they were positive with the IIx probe (Figure 4E). Therefore, the present data identified BF-F3 as being specific for IIb MyHC in pig skeletal muscle.
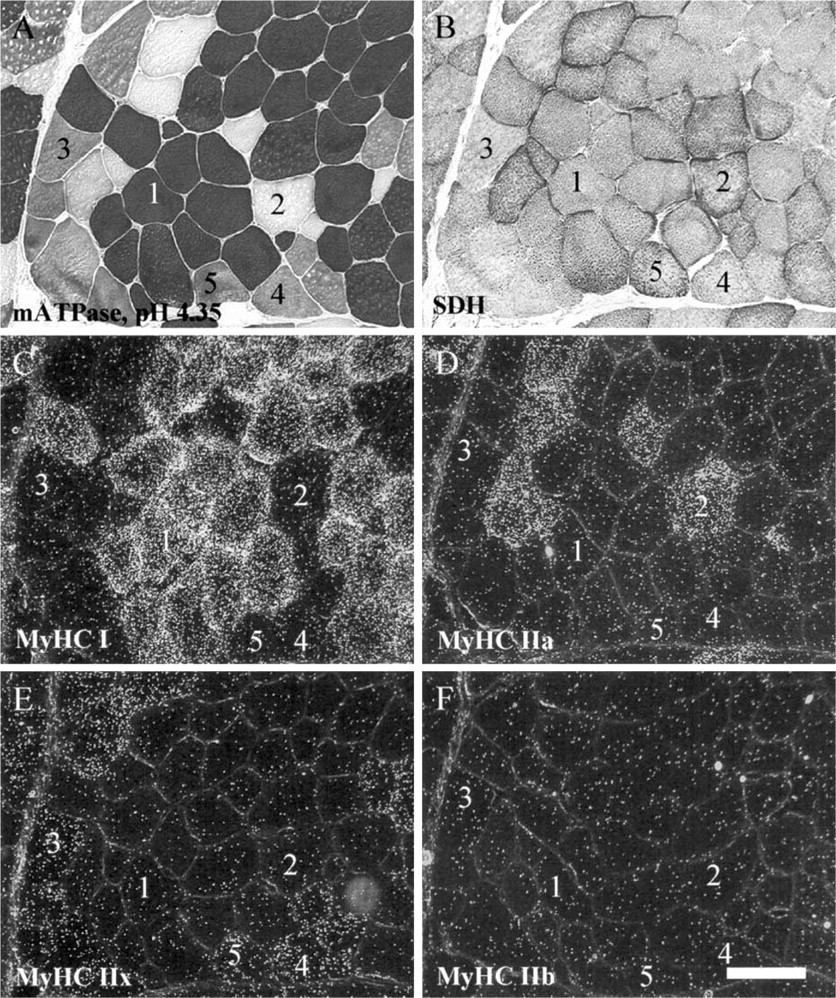
Identification of myofiber types by histochemistry and distribution of the different myosin heavy chain (MyHC) mRNA in rhomboideus muscle. Serial sections were processed as described in Figure 3. Arabic numerals denote corresponding fibers on the serial sections. Bar = 100 μm.
S5–7D4 MAb. In the R muscle where no IIb MyHC was expressed, S5–7D4 (Figure 9D) recognized the same fibers as did SC-71 (Figure 9C). However, in contrast to SC-71, staining intensities were homogeneous between fibers with S5–7D4. Consequently, we conclude that S5–7D4 recognized IIa and IIx MyHC with the same affinity in pig muscle. In L muscle, S5–7D4 labeled the same fibers as did SC-71, as well as additional conventional IIB fibers with a lower intensity (Figure 8D, fiber 7). These fibers were also highly reactive with BF-F3, showing that they contained the IIb MyHC protein (Figure 8E). Interestingly, they were unreactive with the IIx probe (Figure 3E) and strongly labeled with the IIb probe (Figure 3F). Thus, at the protein level, our data identified fiber 7 as a hybrid (IIx + IIb) fiber that was converting to a IIb fiber (Iix→IIb), as suggested by the presence of only the Type IIb mRNA.
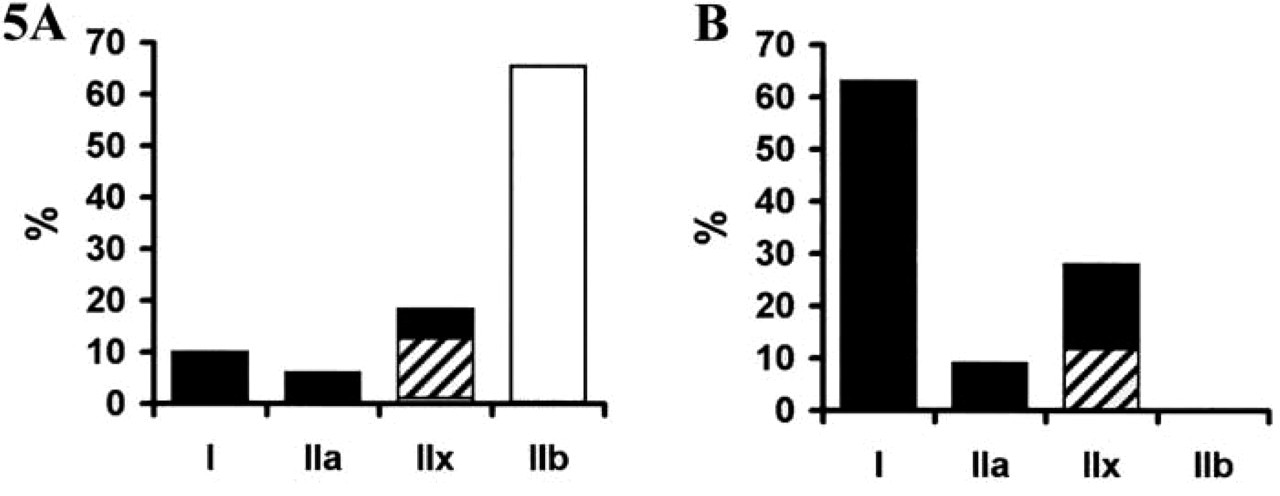
Myofiber type composition of longissimus (
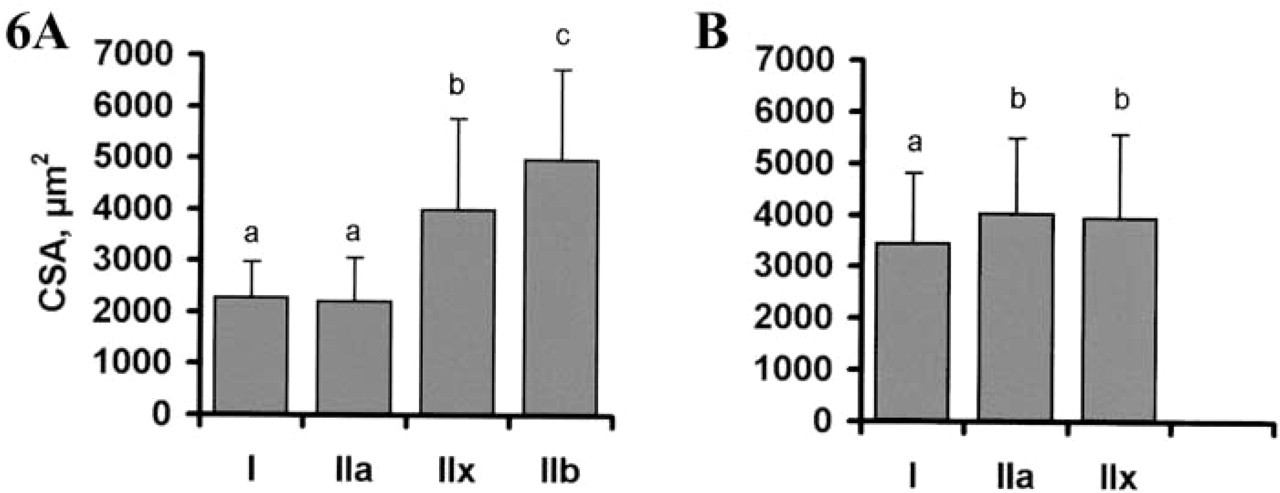
Cross-sectional areas (CSAs) of myofibers in longissimus (
S5–8H2 MAb. In the L muscle, S5–8H2 uniformly labeled all except two fibers (Figure 8F) that were typed IIA by histochemistry (Figure 3A). Similar results were observed in R muscle (Figure 9F). Unstained fibers were reactive with the IIa probe and unreactive with either I, IIx, or IIb probe in both muscles. We conclude that S5–8H2 recognized all but Type IIa MyHC in pig muscle, i.e., I + IIx + IIb MyHCs. Fiber 5 in R muscle deserves special attention. This fiber was stained with the anti-IIa MyHC 6B8 antibody (Figure 9B), whereas no IIa mRNA was present (Figure 4D). In addition, it was moderately stained with S5–8H2 (Figure 9F), suggesting that an MyHC other than IIa was present. At the mRNA level, this fiber expressed only IIx mRNA (Figure 4E), suggesting that it was a hybrid IIa + IIx fiber at the protein level, converting to IIx (IIa→IIx).
Discussion
Pigs have been aggressively selected for rapid, large, and efficient accretion of muscle, which is believed to have deteriorated meat quality. Muscle fibers, quantitatively the most important compartment of skeletal muscle, have long been thought to be important factors influencing meat quality. However, identifying a superior fiber type for meat production remains a tough issue. Because a lack in the accuracy of myofiber typing can be a limiting factor in these studies, the present work was undertaken to bring new insights into muscle fiber types in the pig.
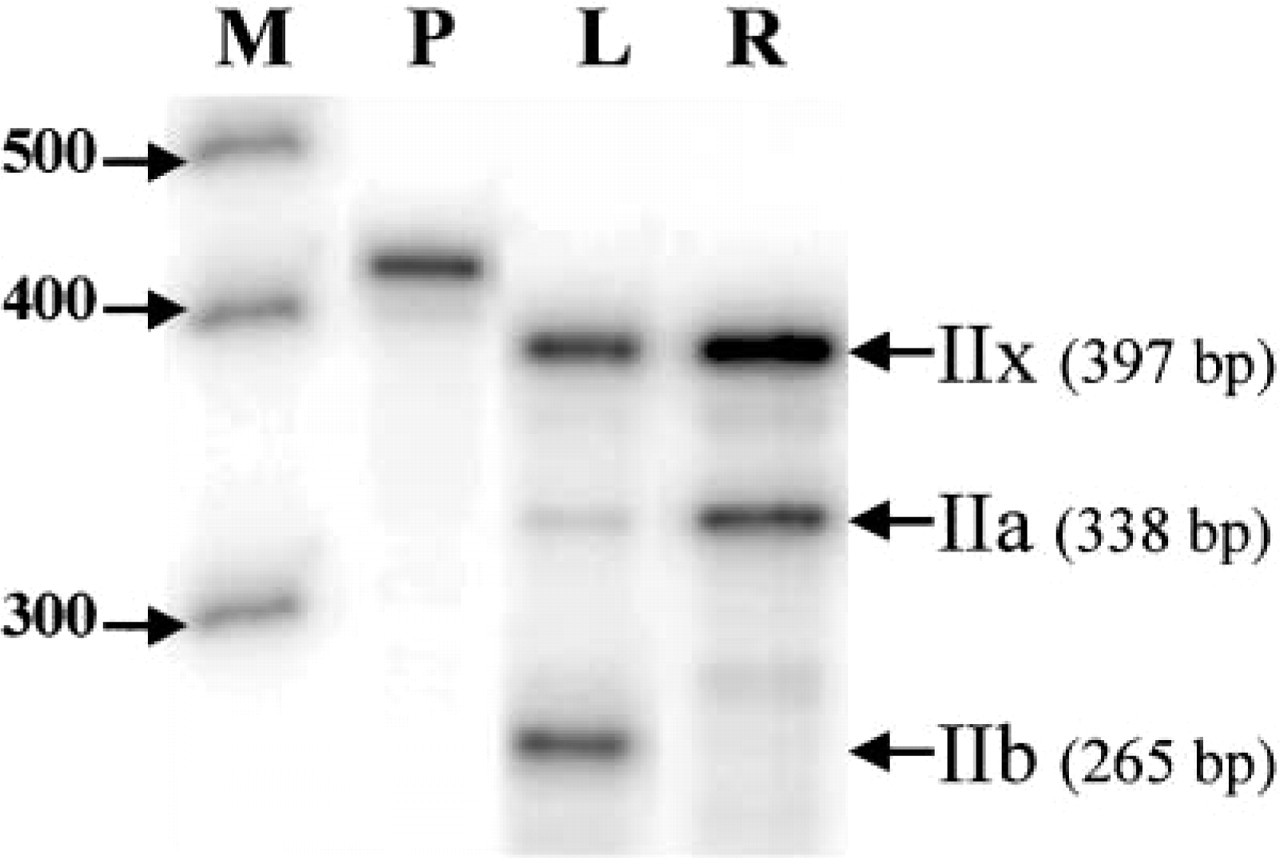
Simultaneous RNase protection analysis of adult fast-twitch Type IIa, IIx, and IIb mRNAs in pig longissimus (L) and rhomboideus (R) muscles. M, RNA size marker; P, IIx probe.
The specificity of each antibody is summarized in Table 1. They were either specific for a single MyHC (I, IIa, or IIb) or reacting with multiple MyHC isoforms (IIa + IIx or I + IIx + IIb). The main limiting factor in the pig remains the lack of a monospecific Type IIx antibody. A specific IIx MAb has been recently described in the mouse, guinea pig, rabbit, cat, and baboon (Lucas et al. 2000) but is not yet available. The specificity of NLC-MHCs as anti-Type I MyHC confirms previous results in pig skeletal muscle (Lefaucheur et al. 1995) and in that of many other species, suggesting that Type I MyHC is highly conserved among species. Antibody 6B8 was previously identified as IIa-specific in pig skeletal muscle (Depreux et al. 2000). The present data confirm those results. Antibody SC-71 recognized both IIa and IIx MyHC, but the staining intensity was higher for IIa than for IIx fibers, suggesting a higher affinity for IIa than for IIx MyHC. Similar differential stainings have also been reported in monkey and human (Fitts et al. 1998). However, these data are different from those reported in rats, in which SC-71 was IIa specific (Schiaffino et al. 1989), showing an interspecies variability in the specificity of SC-71. Antibody S5–7D4 also recognized IIa and IIx MyHC in pig muscle but, in contrast to SC-71, the staining intensities were similar for both IIa and IIx isoforms. The specificity of S5–7D4 in other species remains to be established. BF-F3 was found to specifically react with IIb MyHC in pig muscle, as previously reported in rat muscle (Schiaffino et al. 1989). However, BF-F3 has been shown to be either unreactive or to recognize several fast isoforms in other species such as guinea pig, horse, and cattle (Gorza 1990; Rivero et al. 1996; Duris et al. 2000), still showing a variability of the specificity among species. Finally, S5–8H2 recognized all but Type IIa My-HCs in pig muscle, i.e., I + IIx + IIb. Consequently, S5–8H2 can be used to identify the pure IIa fibers, which will be the only ones to be unstained. In bovine muscle, in which no expression of the IIb MyHC gene has been reported (Tanabe et al. 1998), S5–8H2 also recognizes all but Type IIa fibers, i.e., I + IIx (Duris et al. 2000), which is consistent with reactions in the pig.
Until recently, the consensus among most researchers was that Type IIb MyHC, the isoform exhibiting the most rapid speed of contraction, was expressed only in small mammals, in relation to the relative higher speed of contraction of their muscles (Smerdu et al. 1994; Tanabe et al. 1998; Rivero et al. 1999; Pette and Staron 2000). However, the present study reports unequivocally a very high level of expression of the IIb isoform in 100-kg pig L muscle by combining conventional histochemistry, ICC, ISH, and RPA. A recent study combining conventional histochemistry with ICC suggests that IIb MyHC is also expressed in the llama (Graziotti et al. 2001). Therefore, it can be concluded that the general assumption stating that the very fast Type IIb MyHC isoform is expressed only in small mammals can be rejected. The reasons leading to the high expression of IIb MyHC in pig L muscle are not known. However, because the pig is a meat-producing animal which has been selected, fed, and housed to increase muscle development and growth efficiency, one can speculate that the high expression of IIb MyHC is related to selection and breeding conditions, but further research is needed to test this hypothesis.
Summary of the specificities of the anti-MyHC MAbs determined by immunocytochemistry in pig skeletal muscle a

aStaining intensity: -, negative; +, moderate; ++, strong.
The four isoforms (I, IIa, IIx, IIb) were expressed at the mRNA and protein levels in the L muscle, a fast-twitch glycolytic muscle, whereas no IIb was observed in the R muscle, a mixed highly oxidative muscle. The absence of any IIb in R muscle was confirmed by RPA. This is in accordance with previous results showing no IIb expression in the deep red portion of pig semitendinosus muscle using ISH (Lefaucheur et al. 1998). The different expression of MyHC among muscles is likely related to their function. Indeed, the L muscle is a fast-twitch glycolytic muscle involved in voluntary movements of the back, whereas the rhomboideus is a postural mixed-oxydo-glycolytic muscle involved in posture by supporting the head. To our knowledge, no similar study has compared the MyHC pattern in longissimus and rhomboideus muscles of small mammals. However, a study carried out in the mouse showed that IIb MyHC is the predominating isoform in tibialis anterior, extensor digitorum longus, gastrocnemius, plantaris, quadriceps, pectoralis, and biceps, whereas IIb is absent in the soleus, a postural muscle, and is present in a small amount in the diaphragm, a muscle that is continually in use (Agbulut et al. 1996). The prevalence of IIb MyHC in most limb muscles of the mouse is assumed to be related to its small size and consequently rapid movements. In humans, we are not aware of any study that has compared the MyHC composition of these two muscles. Finally, because myofiber contractile properties are predominantly influenced by innervation, a difference of innervation between L and R muscles is likely involved, in relation to their different locations and functions, but no data are available.
Within the typical islets, MyHC expression was spacially regulated from the center to the periphery with a gradual expression in the rank order of I→IIa→Iix→IIb in the L muscle and I→IIa→IIx in the R muscle. In L muscle, a corresponding gradual decrease in oxidative metabolism (I = IIa > IIx > IIb), and increase in CSA (I = IIa < IIx < IIb) occurred, showing that metabolic and size properties of muscle fibers varied according to their MyHC content. The gradient in MyHC expression (I→IIa→Iix→IIb) corresponds to increasing mATPase activity, contractile velocity, tension cost, fatigability, and excitation threshold (Bottinelli et al. 1996; Hilber et al. 1999; Kernell et al. 1999; He et al. 2000), suggesting that the fibers could be gradually recruited from the center to the periphery of the islets in relation to the intensity of the movement. Therefore, the highly organized type grouping arrangement of myofibers could also be functionally relevant within a muscle. In particular, Type I fibers would be those most easily recruited for long-lasting weak contractions, whereas Type IIb fibers would be mainly employed in short-lasting bouts of forceful contractions. However, even though this type grouping distribution appears to be functionally and physiologically relevant, it is not a prerequisite for the functionality of a muscle because most mammals exhibit a mosaic pattern and are nevertheless fully functional. Recently, a similar type grouping distribution has been described in the llama (Graziotti et al. 2001), but the reason for such a peculiar distribution is not fully known. Developmentally, the type grouping is tightly related to primary and secondary myogenesis in the pig (Lefaucheur et al. 1995). Indeed, the central fiber of each islet of slow-twitch Type I fibers originates from a primary myotube that serves as a scaffold for secondary fibers. Around birth, some secondary fibers in the direct vicinity of each primary myotube mature to Type I fibers, whereas remaining secondary fibers mature to Type IIa, IIx, or IIb fibers.
In most fibers, the distribution of Type I, IIa, IIx, and IIb transcripts closely matched that of the corresponding proteins, which confirms previous results showing that the expression of MyHC isoforms is mostly transcriptionally regulated (Cox and Buckingham 1992). This is the case for fibers 1 (I), 2 (IIa), 3 (IIb), and 4 (IIx) in L muscle and fibers 1 (I), 2 (IIa), and 3 (IIx) in R muscle. However, some hybrid fibers were also observed, in particular IIx + IIb hybrid fibers. Thus, 11.5% of the fibers reacted with both S5–7D4 and BF-F3 in L muscle. Because these fibers did not react with 6B8, they probably expressed IIx and IIb proteins. Interestingly, co-expression was much less common at the mRNA level (2.2% for IIx + IIb in L muscle), which could be due to the shorter half-life of mRNAs than of proteins. The low level of co-expression at the mRNA level suggests that a fiber usually synthesizes only one isoform at a time, even though several protein isoforms can be present in the same fiber. Therefore, fibers that contained two My-HCs at the protein level, but only one transcript, are most probably fibers that are converting to the type corresponding to the expressed mRNA, whereas the MyHC isoform lacking its corresponding mRNA is no longer being synthesized and will progressively disappear as a result of protein turnover. Some fibers containing an MyHC protein but not the corresponding transcript have also been found in human muscle (Andersen and Schiaffino 1997). These authors also proposed that these fibers represent transitional fibers and that the relative content of each isoprotein and mRNA gives a clue to the direction of change in MyHC expression. Interestingly, because the myofiber is a multinucleated syncytium, an alternative explanation for the occasional appearance of a protein without its mRNA is that the fiber in question contains mRNA for more than one MyHC isoform but that each nucleus expresses only one mRNA or expresses it in pulses (Newlands et al. 1998). In this case, transverse sections would give a limited sampling of mRNA expression in a muscle fiber. Because protein is longer-lived than the mRNA and/or because the protein may diffuse more than its mRNAs, a mismatch between the presence of the protein and its corresponding mRNA can occur. In further experiments, the existence of a second mRNA could be addressed experimentally by looking at longitudinal muscle sections. The mismatch between proteins and transcripts illustrates the plasticity of muscle fibers. All of the transitions we observed were either I↔IIa, IIa↔IIx or IIx↔IIb, supporting the general idea of an obligatory pathway of MyHC gene expression in the rank order of I↔IIa↔IIx↔IIb under normal physiological conditions (Schiaffino and Reggiani 1994; Pette and Staron 2000).
In conclusion, the present integrated data confirm that fast-twitch Type IIB skeletal muscle fibers identified on the basis of their mATPase activities have been misclassified in the pig in most previous studies. The combination of ISH with ICC shows they should be subdivided into Types IIx and IIb, as well as hybrid IIx + IIb fibers. At the protein level, a monospecific Type IIx antibody is still not available to accurately type myofibers according to their MyHC composition in pig muscle. Interestingly, IIx fibers exhibited high or moderate SDH staining, whereas pure IIb fibers were weakly stained, showing that only weakly stained IIB fibers did correspond to real IIb fibers. However, the continuum between SDH-positive and -negative stainings makes the use of this classification not always very accurate. Recently, Gil et al. (2001) identified four fiber types in pig longissimus muscle by combining acid and alkaline preincubations on serial sections before mATPase staining. However, the correspondence with Types I, IIa, IIx, and IIb fibers remains to be definitely established. Co-expression of MyHCs was more common at the protein than the mRNA level and was mostly observed for IIx and IIb MyHCs, suggesting a fine tuning of these two genes and a strong influence of their expression on myofiber plasticity. The present study also shows that only the combination of immunocytochemistry with in situ hybridization allows a clear understanding of the dynamic process involved in fiber type transitions by giving a clue to the direction of change in MyHC gene expression. Finally, because MyHC is the most abundant protein in muscle and because of the influence of its polymorphism on contractile, metabolic, and size properties of myofibers, further research is still needed to establish and understand the importance of MyHC polymorphism in muscle growth efficiency and meat quality in the pig.
Footnotes
Acknowledgements
We are grateful to Dr David Gerrard for his generous gift of the monoclonal antibody 6B8, to Dr Gillian S. Butler-Browne for critical reading of the manuscript, and to Véronique Corbière for her contribution in cloning the probes.