
Abstract
TREK-1 is one of the important potassium channels for regulating membrane excitability. To examine the distribution of TREK-1 in the rat carotid body, we performed RT-PCR for mRNA expression and in situ hybridization and immunohistochemistry for tissue distribution of TREK-1. RT-PCR detected mRNA expression of TREK-1 in the carotid body. Furthermore, in situ hybridization revealed the localization of TREK-1 mRNA in the glomus cells. TREK-1 immunoreactivity was mainly distributed in the glomus cells and nerve fibers in the carotid body. TREK-1 may modulate potassium current of glomus cells and/or afferent nerve endings in the rat carotid body.
T
Several potassium channels show an oxygen-sensing property: TASK-1 in cerebellar granule neurons (Plant et al. 2002) and in glomus cells (Buckler et al. 2000); TASK1 and TASK-3 in H146 cells (Hartness et al. 2001); THIK-1 in glossopharyngeal neurons (Campanucci et al. 2003); Kv3.4 and Kv4.3 in rabbit glomus cells (Sanchez et al. 2002). However, TREK-1 sensitivity for oxygen is argued. Miller et al. (2003) reported that acute hypoxia occluded human TREK-1 expressed in the HEK293 cells under ischemic and/or acidotic conditions.
Furthermore, it is reported that inhibited TREK-1 current by intracellular alkanization was further suppressed by hypoxia (Miller et al. 2004). On the contrary, other reports demonstrated that TREK-1 was not oxygen sensitive (Buckler and Honore 2005; Caley et al. 2005). Furthermore, it has been demonstrated that several oxygen-sensing potassium channels are present in the peripheral chemosensor, the carotid body. Voltage-gated potassium channels Kv3.4 and 4.3 were demonstrated in the rabbit glomus cells (Sanchez et al. 2002). TASK-1 and TASK-3 immunoreactivities were observed in the rat glomus cells (Yamamoto et al. 2002).
To examine the distribution of TREK-1 in the rat carotid body, we performed RT-PCR for mRNA expression and in situ hybridization and immunohistochemistry for tissue distribution of TREK-1. Furthermore, double immunofluorescence by use of several cell markers was performed to identify cellular distribution of TREK-1.
Materials and Methods
Animals
Wistar rats of both sexes (n = 10) were used in our studies, which were approved by the local Ethics Committee. For RT-PCR analysis, four rats were sacrificed in the chamber filled with diethyl ether gas, and the bifurcations of carotid arteries were corrected. Whole carotid body was further dissected out with dissecting microscope and moved into RNA stabilization reagent (RNAlater; Qiagen, Tokyo, Japan). For immunohistochemistry and in situ hybridization, six rats were anesthetized intraperitoneally with pentobarbital (15 mg/kg) and transcardially perfused with Ringer's solution (500 ml) followed by Zamboni's fixative (4% paraformaldehyde, 0.5% picric acid in 0.1 M phosphate buffer; pH 7.4, 500 ml). The bifurcations of carotid arteries were dissected out and further fixed in the same fixative for 5 hr. The tissues were then soaked in 30% sucrose in phosphate-buffered saline (PBS; pH 7.4) and frozen. They were serially sectioned at 8 μm and mounted on glass slides coated with chrome alum-gelatin.
Primers for RT-PCR

RT-PCR Amplification
Two carotid bodies were mechanically homogenized for 5 min, and RNA was then extracted by commercial kit (RNeasy Fibrous tissue kit; Qiagen). mRNA templates were incubated with DNAase I (Takara; Tokyo, Japan) for 30 min at 37C before use. RT-PCR was performed by use of Qiagen OneStep RT-PCR kit with specific primers for TREK-1. For positive control, primers for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were also used to detect mRNA expression of housekeeping gene. Primers for TREK-1 and GAPDH are shown in Table 1. Reverse transcription was done for 30 min at 50C, and initial PCR activation was then performed for 15 min at 95C. PCR amplification was performed for 40 times as follows: 30 sec at 94C for denaturation, 30 sec at 55C for annealing, 1 min at 72C for extension. After PCR amplification, samples were applied for 10 min at 72C for final extension. PCR end products were visualized on 2% agarose gels with ethidium bromide. For negative control experiment, mRNA template was omitted.
In Situ Hybridization
For in situ hybridization, sections were washed with PBS and then rinsed with 5x SSC. The sections were incubated with 50% formamide in 5x SSC including 1 μg/ml digoxygenin-labeled oligonucleotide probe for TREK-1 (accession number AF385402; position 1264-1308; 5î-CACCGACAGGGTCCTCCTACATGGAGTCAGTTCCTGGTTATGGTT-3î) with 40 μg/ml fragmented salmon sperm DNA for 15 hr at 50C. Sections were then washed in 5x SSC, 2x SSC, 0.1x SSC, and buffer 1 (constitute; 100 mM Tris, 150 mM NaCl, pH 7.5) and incubated with 1% normal goat serum in buffer 1 for 1 hr at room temperature. Sections were then washed with buffer 1 and reacted with alkaline phosphatase-labeled sheep anti-digoxygenin antibody Fab fragment (1:2000; Roche Diagnostics, Mannheim, Germany) for 2 hr at 40C. Sections were washed with buffer 1 and buffer 3 (100 mM Tris, 150 mM NaCl, 50 mM MgCl2, pH 9.5) and incubated with 5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazolium solution for visualization of reacted sites. For positive control, antisense dioxygenin-labeled oligonucleotide probe for poly-A was used. For negative control, randomized digoxygenin-labeled oligonucleotide probes were used.
Immunohistochemistry
Sections were incubated for 60 min with non-immune donkey serum (1:50) and rinsed with phosphate-buffered saline (PBS; pH 7.4). Sections were then incubated overnight at 4C with goat polyclonal antisera against TREK-1. After incubation, sections were washed again with PBS and treated with TRITC-labeled donkey antibody against goat IgG for 2 hr at room temperature. After washing with PBS, sections were mounted in glycerol-PBS and examined with an epifluorescence microscope (E-600; Nikon, Tokyo, Japan). Negative controls were incubated with preabsorbed antibody using blocking peptide (SC-11556P; Santa Cruz Biotechnology, Santa Cruz, CA), 1 μg antigen/1 μg antibody, or PBS. Details of antibodies are shown in Table 2.
Double Immunofluorescence
Cryostat sections were also used for double immunofluorescence for TREK-1 with synaptophysin, glial fibrillary acidic protein (GFAP), vimentin, or α-smooth muscle actin (ASMA). After incubation with normal donkey serum, sections were incubated with goat polyclonal TREK-1 together with antibody against mouse monoclonal antibody against synaptophysin, rabbit polyclonal antibody against GFAP, mouse monoclonal antibody against vimentin, or mouse monoclonal antibody against ASMA for 12 hr at 4C. Sections were then incubated with a mixture of TRITC-labeled donkey anti-goat IgG with FITC-labeled donkey anti-mouse IgG or with FITC-labeled donkey anti-rabbit IgG for 2 hr at 25C. Sections were coverslipped with glycerol-PBS and examined under an epifluorescence microscope. Details of antibodies are shown in Table 2.
Antibodies used in the present study

RT-PCR for TREK-1 mRNA in the rat carotid body. The appropriate-sized PCR product is visualized. 1, 100-bp molecular weight marker; 2, carotid body; 3, glyceraldehyde-3-phosphate dehydrogenase (GAPDH); 4, negative control for TREK-1.
Results
RT-PCR
Using RT-PCR, mRNA expressions of TREK-1 and GAPDH were detected at 125 and 401 bp, respectively (Figure 1, Lanes 2 and 3). In the negative control experiment, no product was found (Figure 1, Lane 4).
In Situ Hybridization
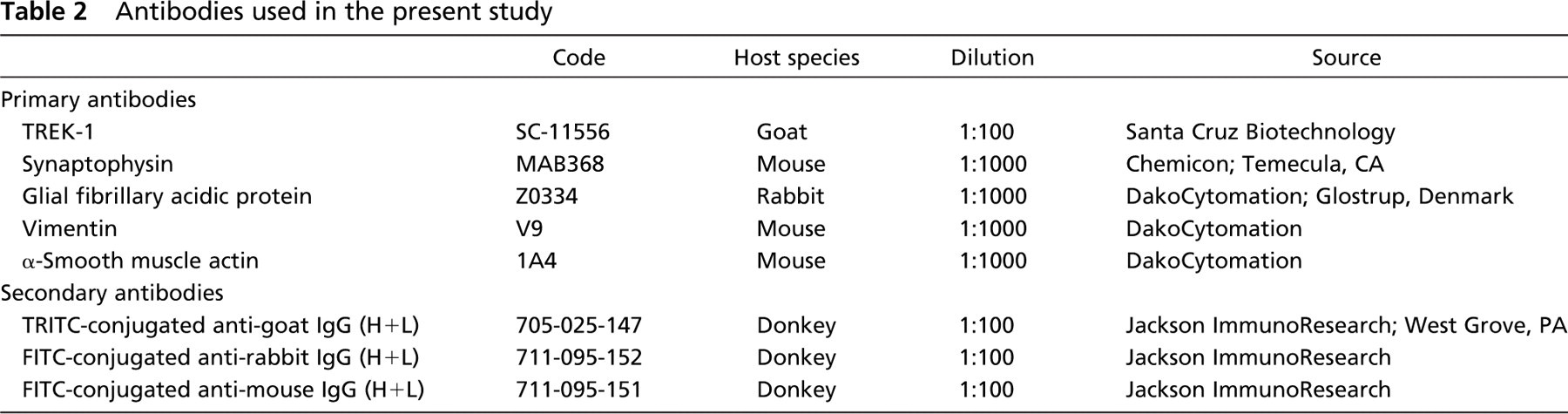
Intense signals for TREK-1 mRNA were observed in the cytoplasm of the glomus cells by in situ hybridization (Figures 2A and 2B). Ubiquitous distribution of mRNA signals was observed in positive control section that stained with probe for poly-A sequence (Figure 2C). No signal was observed in the negative control (Figure 2D).
Immunohistochemistry
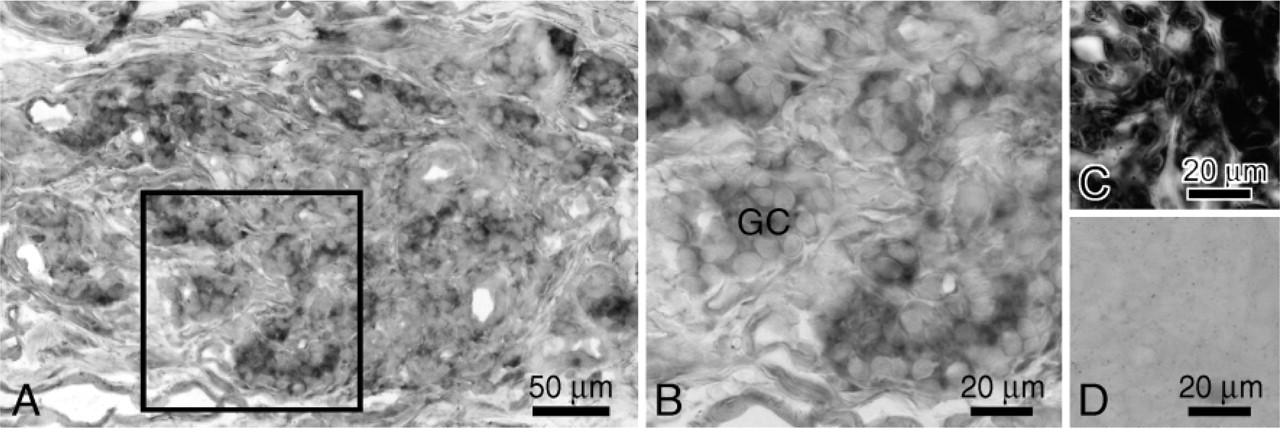
Immunoreactivity for TREK-1 was observed in the rat carotid body (Figures 3A and 3B). Sections incubated with either PBS or preabsorbed antibody instead of primary antibody showed no immunoreactivity (Figure 3C). Glomus cells and nerve fibers with various thicknesses were strongly immunoreactive for TREK-1. Furthermore, dot-like structures with immunoreactivity for TREK-1 were observed around the clusters of glomus cells. Vascular endothelial cells were also immunoreactive, but fibroblasts were not immunoreactive for TREK-1.
Double Immunofluorescence
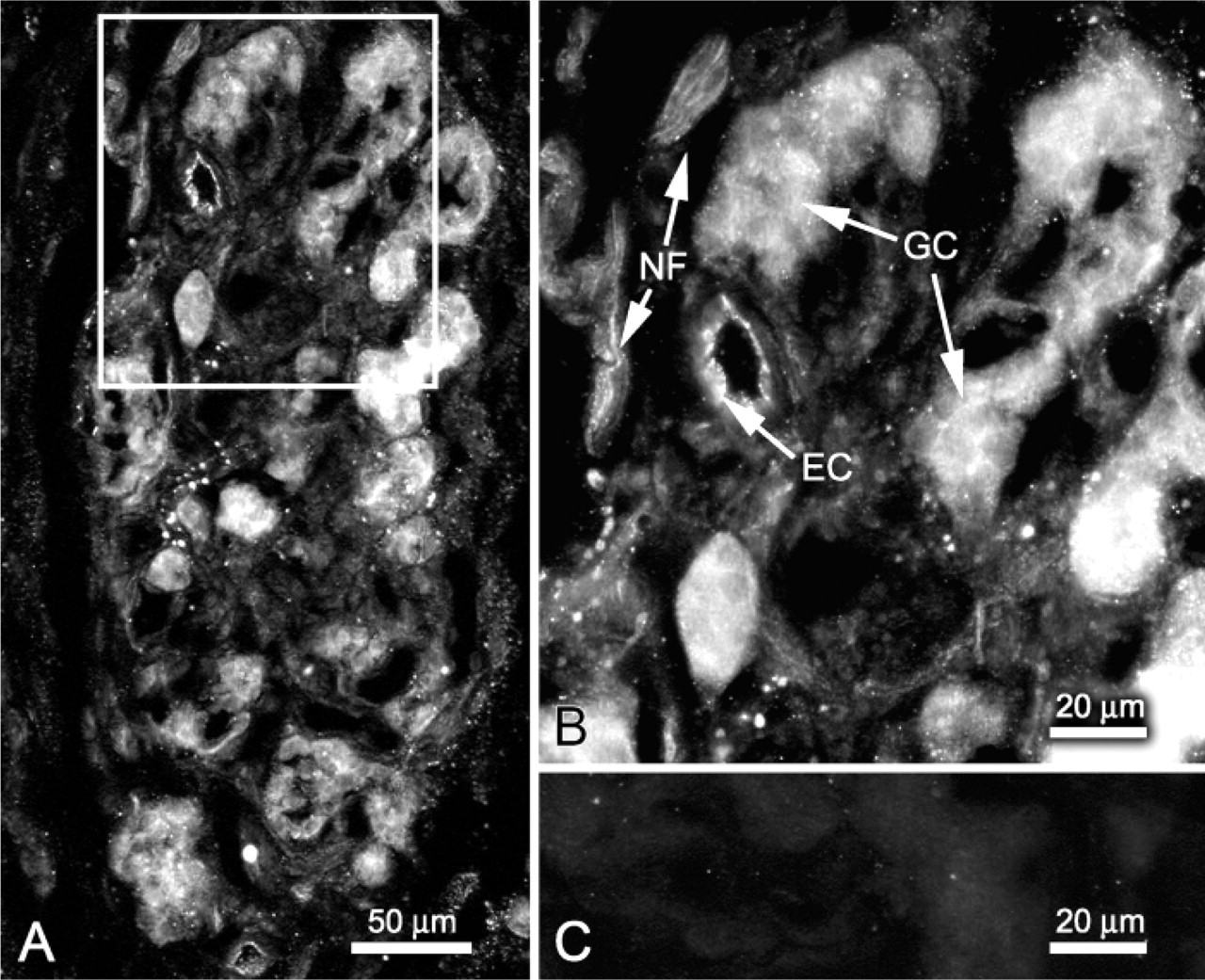
Immunoreactive sites for synaptophysin, the general marker for glomus cells, were also immunoreactive for TREK-1 (Figures 4A-4C). Immunoreactive sites for GFAP, marker for sustentacular cells, and for vimentin, marker for sustentacular cells and fibroblasts, did not show TREK-1 immunoreactivity (Figures 4D-4I). GFAP-immunoreactive Schwann cells were observed in the nerve bundles in the carotid body but were not immunoreactive for TREK-1. Vascular smooth muscle cells immunoreacted for ASMA were not immunoreactive for TREK-1 (Figures 4J-4L).
Discussion
It has been reported that TREK-1 is mainly expressed in the central nervous system at mRNA level (Medhurst et al. 2001; Talley et al. 2001) and protein level (Hervieu etal. 2001). Furthermore, expression of TREK-1 is also reported in other excitatory cells such as trigeminal and petrosal ganglion cells (Matsumoto et al. 2001), smooth muscle cells (Koh et al. 2001), and cardiomyocyte (Tan et al. 2004). TREK-1 in the glomus cells and possibly afferent nerve fibers in the carotid body, which was revealed in the present study, may indicate TREK-1 would be expressed in the peripheral chemosensory cells. The dot-like structures around the glomus cells may be the TREK-1 channels bound to excitable membrane of nerve fibers and nerve endings. Our previous study demonstrated that immunoreactivities for other K2P channels, TASK-1, TASK-2, TASK-3, and TRAAK, were found in the glomus cells and/or nerve fibers in the carotid body (Yamamoto et al. 2002). Thus, the glomus cell and nerve fibers in the rat carotid body expressed a wide variety of K2P channels to modulate K+ current of resting membrane and/or depolarizing membrane potential.
In situ hybridization for TREK-1 mRNA in the carotid body. Positive signals for TREK-1 are present in the glomus cells (GC). Rectangle in (
Immunolocalization of TREK-1 in the rat carotid body. Rectangle in (
It has been reported that acute hypoxia occluded human TREK-1 expressed in the HEK293 cells under ischemic and/or acidotic conditions (Miller et al. 2003) and that inhibited TREK-1 current by intracellular alkalization was further suppressed by hypoxia (Miller et al. 2004). If TREK-1 were an oxygen-sensitive channel, it would be closed by hypoxic stimulation to modulate the cellular membrane of glomus cells in cooperation with other oxygen-sensing potassium channels, e.g., TASK-1, TASK-3, Kv3.4, and Kv4.3. Furthermore, it has been reported that TREK-1 is inhibited by glutamate via group 1 metabotropic glutamate receptors (Chemin et al. 2003). During hypoxia, glutamate is released in the nucleus tractus solitarii that is the central terminal part of the carotid sinus nerve (Mizusawa et al. 1994). Torrealba et al. (1996) reported that glutamate is not released from the cat carotid body even in the high concentration of potassium. However, glutamate is still a candidate of excitatory transmitter between the glomus cells and afferent nerve endings, because glutamate-immunoreactive glomus cells and nerve fibers were observed in the carotid body of cat (Torrealba et al. 1996) and guinea pig (Kummer 1997). Thus, TREK-1 may enhance membrane excitation of the glomus cells via involvement of glutamatergic neurotransmission in addition to direct occlusion during hypoxia.
On the contrary, other reports demonstrated that TREK-1 was not oxygen sensitive (Buckler and Honore 2005; Caley et al. 2005). If TREK-1 is not an oxygen-sensitive molecule, another hypothesis for TREK-1 function is raised. Generally, it is suggested that TREK-1 acts as a potent neuroprotector (Duprat et al. 2000; Lauritzen et al. 2000). It opens with arachidonic acid and certain phospholipids such as phosphatidylinositol 4-monophosphate and phosphatidylinositol 4,5-bisphosphate (Maingret et al. 2000). In the carotid body, hypoxia increased the activity of phospholipase C, one of the important enzymes for generation of phospholipids (Pokorski and Strosznajder 1993). Furthermore, Ca2+ enhanced the production of arachidonic acid in homogenized samples of the carotid body (Strosznajder 1996). According to these reports, TREK-1 may act for cellular protection. A second hypothesis is that TREK-1 in the glomus cells may close with arachidonic acid generated under hypoxia to protect membrane excitation.
Double immunofluorescence for TREK-1 combined with synaptophysin (
In conclusion, TREK-1 would be important for K+ current modulation of the glomus cells and/or afferent nerve endings in the rat carotid body. However, two contradictory hypotheses on TREK-1 function are proposed at present. One hypothesis is that TREK-1 enhances membrane depolarization of glomus cells by hypoxic stimulation. A second hypothesis is that it is protected via phospholipids and/or arachidonic acid. To clarify the exact function of TREK-1 in the carotid body, further studies are needed.
Footnotes
Acknowledgments
This study was supported in part by the Grant-in-Aid for Young Scientists (