
Abstract
Vimentin, an intermediate filament component, has been identified in many mesenchymal cells by a variety of LM and EM immunolabeling techniques. In our study, several tissue-processing conditions and monoclonal and polyclonal antibodies against vimentin were screened for immunostaining of rat incisor odontoblasts. Using postembedding colloidal gold immunocytochemistry, we were unable to detect any convincing vimentin antigenicity in these cells, but one of the monoclonal antibodies (V9-S) unexpectedly resulted in intense labeling over intra- and extracellular compartments that normally are strongly immunoreactive with anti-amelogenin antibodies. Blocking experiments showed that V9-S binding was competed by anti-amelogenin antibody. Immunoblots indicated that enamel proteins reacted with this anti-vimentin antibody after fixation with glutaraldehyde. These data suggest that the observed immunoreaction is directed against an epitope apparently created by crosslinking of enamel proteins during fixation. Although the labeling cannot be considered specific, it is nevertheless selective because it is very precisely localized over compartments containing enamel proteins and shows no binding to other calcified dental tissues, including dentin and bone. The V9-S antibody can therefore be used as a reliable probe to identify the presence and distribution of amelogenins in fixed tissues.
T
The initial objective of this study was to characterize the frequency with which odontoblast processes invaginate into differentiating ameloblasts. Because these small cytoplasmic extensions are difficult to identify and distinguish morphologically in cross-sections, we originally planned to use vimentin antibodies to “tag” and make them more visible. This intermediate filament protein is typically associated with mesenchymal cells (Lazarides 1982; Steinert and Roop 1988; Van de Klundert et al. 1993) and has been found in odontoblasts by immunofluorescence (Lesot et al. 1982; Sigal et al. 1985; Fausser et al. 1990; Webb et al. 1995) and immunoperoxidase labeling (Lombardi et al. 1992b; Byers and Sugaya 1995). Although vimentin has been revealed in many mesenchymal cell types by immunogold labeling (Shapiro et al. 1995; Kameda 1996a,b), we were unable to obtain evidence of vimentin antigenicity in odontoblasts under the tissue processing conditions used in this study. Instead, very strong immunolabeling was found with one of the antibodies over compartments that normally contain enamel proteins. The present study was therefore redirected to further examine this unexpected crossreactivity and to determine whether it is selective for enamel proteins.
Materials and Methods
Tissue Preparation for Immunocytochemistry
Male Wistar rats (Charles Rivers Canada; St-Constant, Quebec, Canada) weighing approximately 100 g were anesthetized with chloral hydrate (Sigma Chemical; St Louis, MO) and perfused through the ascending aorta with lactated Ringer's solution (Abbott; Montreal, Quebec, Canada) for 30 sec and then with either 4% paraformaldehyde containing 0.1% glutaraldehyde or 1% or 2.5% glutaraldehyde for 20 min. All fixatives were buffered with 0.08 M sodium cacodylate containing 0.05% CaCl2 at pH 7.3. After perfusion, the hemimandibles were removed and immersed in their respective fixative overnight at 4C. Some hemimandibles were decalcified in 4.13% disodium EDTA at 4C (Warshawsky and Moore 1967) for 3 weeks. Segments (5 mm) from the apical ends of both calcified and decalcified incisors were prepared and washed repeatedly in 0.1 M sodium cacodylate buffer, pH 7.3. They were then postfixed with potassium ferrocyanide-reduced OsO4 (Neiss 1984) for 2 hr at 4C and conventionally processed for embedding in various resins as described below. All animal procedures described above were in accordance with guidelines of the Comité de Déontologie de l'Expérimentation sur les Animaux of Université de Montréal.
Thin sections of the tissues were cut with a diamond knife on a Reichert Ultracut E ultramicrotome, mounted on 200-mesh nickel grids having a carbon-coated Formvar film, and processed for postembedding colloidal gold immunolabeling (reviewed in Bendayan 1995b). Sections of Epon- and LR White-embedded tissues were stained with 4% aqueous uranyl acetate for 5 min and with Reynolds lead citrate for 3 min, whereas those of Lowicryl K4M-embedded samples were stained for 10 min with uranyl acetate and 1 min with lead citrate. All sections were examined in a JEOL JEM 2000FX-II transmission electron microscope operated at 80 kV.
Tissue Processing for Embedding
Epoxy Resin. Some incisor samples fixed in 1% or 2.5% glutaraldehyde and postfixed in reduced osmium were dehydrated in a graded series of acetone (30-100%). They were then infiltrated for 8-12 hr with a 1:1 and 2:1 mixture of Epon substitute resin (Meca Laboratories; Montreal, QC, Canada) and pure acetone, followed by pure resin for 5 hr under vacuum. Tissues were embedded in a fresh mixture of pure resin and were polymerized for 2 days at 60C.
LR White. Other tooth samples fixed in 1% glutaraldehyde and postfixed in reduced osmium were dehydrated in graded concentrations of ethanol (30-100%) and infiltrated with a 1:1 mixture of LR White resin (hard grade; Bio-Rad Laboratories, Watford-Hertfordshire, UK) and absolute ethanol for 1 hr, followed by three 1-hr changes in pure LR White resin, all at room temperature (RT). The tissues were then embedded in pure LR White in gelatin capsules and the resin was polymerized at 48C for 2 days.
Lowicryl K4M. Nonosmicated tissue segments from animals fixed in paraformaldehyde/glutaraldehyde mixture were processed for embedding in a cryosubstitution apparatus (CS auto; Reichert-Jung, Wien, Austria). They were dehydrated in 30% methanol at 0C, in 50% methanol at — 20C, and in 75% methanol at −35C for 30 min each, and in 90% methanol for 60 min at — 35C. The tissues were then infiltrated at — 35C for periods of 60 min with 1:1 and 2:1 mixtures of Lowicryl K4M (Chemische Werke Lowi; Waldkraiburg, Germany) and 90% methanol, followed by pure Lowicryl K4M overnight. They were then embedded in pure resin in plastic molds and polymerized for 2 days at −35C using the UV illuminator arrangement of the CS auto apparatus.
Immunocytochemical Procedures
Antibody Incubation. All immunocytochemical procedures were performed at RT. Grid-mounted sections of osmicated tissues embedded in Epon or LR White were pretreated with saturated sodium metaperiodate for 1 hr (Bendayan and Zollinger 1983) and rinsed with distilled water before incubation with the respective antibodies, as follows. Sections were floated for 5 min on a drop of 0.01 M PBS containing 1% ovalbumin (Sigma) and then transferred and incubated for periods ranging from 1 to 9 hr on a drop of diluted primary antibody (see Table 1 for a list of the various antibodies used and their abbreviated names). After incubation with some of the primary antibodies, sections were rinsed with PBS and again blocked with PBS-1% ovalbumin for 5 min before further incubation for 1 hr with a secondary antibody (see Table 1). The incubation with secondary antibodies was followed by rinsing with PBS and floating on PBS-1% ovalbumin for 5 min. In all cases, the sites of antibody-antigen binding were revealed by floating grids on a drop of protein A-gold complex (Bendayan 1995b) prepared in house using colloidal particles of approximately 14 nm (Frens 1973). The grids were then rinsed with PBS, washed thoroughly with distilled water, and air-dried before heavy metal staining (see above).
Primary and secondary antibodies used for incubation, their working dilutions, time of incubation, and commercial sources/catalog and lot numbers
Sigma: Sigma Chemical, St Louis, MO; BM: Boehringer Mannheim, Mannheim, Germany; OTC: Organon Teknika (Cappel), Durham, NC; #, catalog number; L, lot number.
The antibody from Sigma was provided as ascites fluid and from BM as a lyophilized powder.
For controls, grid-mounted sections were incubated with secondary antibody only (see Table 1) followed by protein A-gold, or with protein A-gold alone.
Evaluation of the Specificity of the Anti-vimentin Immunolabeling by Competitive Binding
Lectin-Gold Cytochemistry. Tissue sections of Lowicrylembedded samples were floated on a drop of wheat germ agglutinin (WGA) at a concentration of 25 μg/ml for 1 hr, rinsed with PBS, and incubated for 30 min with ovomucoid-gold (8-nm) prepared as described in Nanci et al. (1989) and diluted 1:20. Both WGA and ovomucoid were purchased from Sigma.
Immunolabeling with Antibody to Amelogenin. Sections of nonosmicated tissues were first incubated with AMEL antibody (see Table 1) for 1 hr, rinsed with PBS, and floated on PBS-1% ovalbumin (5 min), followed by V9-S antibody (diluted 1:10) for 1 hr. To exclude the possibility that steric hindrance of the AMEL IgG might affect the labeling pattern of V9-S antibody, nonosmicated sections were floated on WGA (25 μg/ml) for 1 hr before incubation with clone V9-S (dilution 1:10) for 1 hr. All sections were then rinsed and incubated with protein A-gold as described above.
Immunoblotting. Purified vimentin (Sigma), human fibroblast lysate, and proteins extracted from freeze-dried secretory stage enamel samples (Smith et al. 1989) were separated by SDS-PAGE and transferred onto 0.45 μm nitrocellulose membranes for immunoblotting as previously described (Chen et al. 1995). The blots were probed with V9-S antibody and AMEL antibody and revealed with appropriate alkaline phosphatase-conjugated secondary antibodies (Chen et al. 1995). Some blots were fixed with 1% glutaraldehyde before incubation with the antibodies.
Results
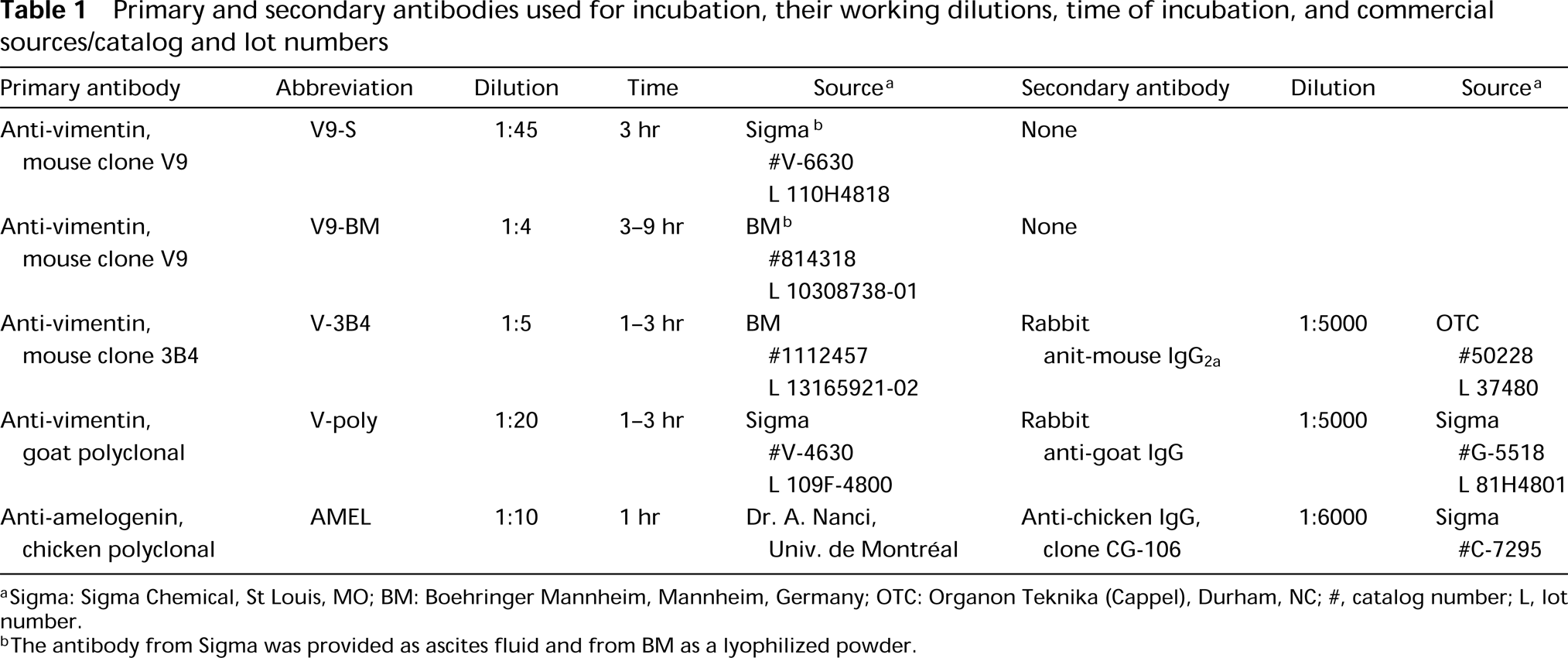
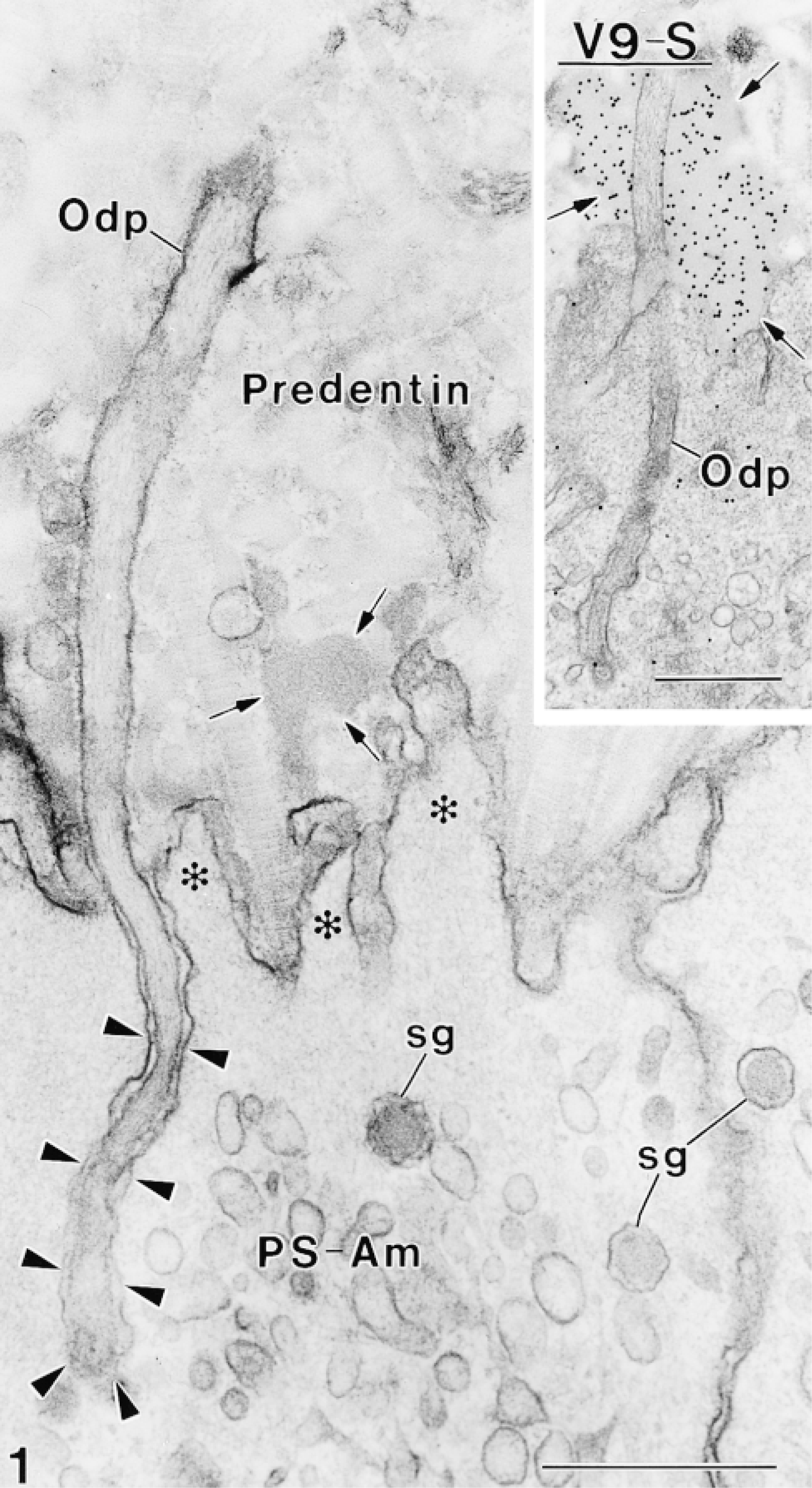
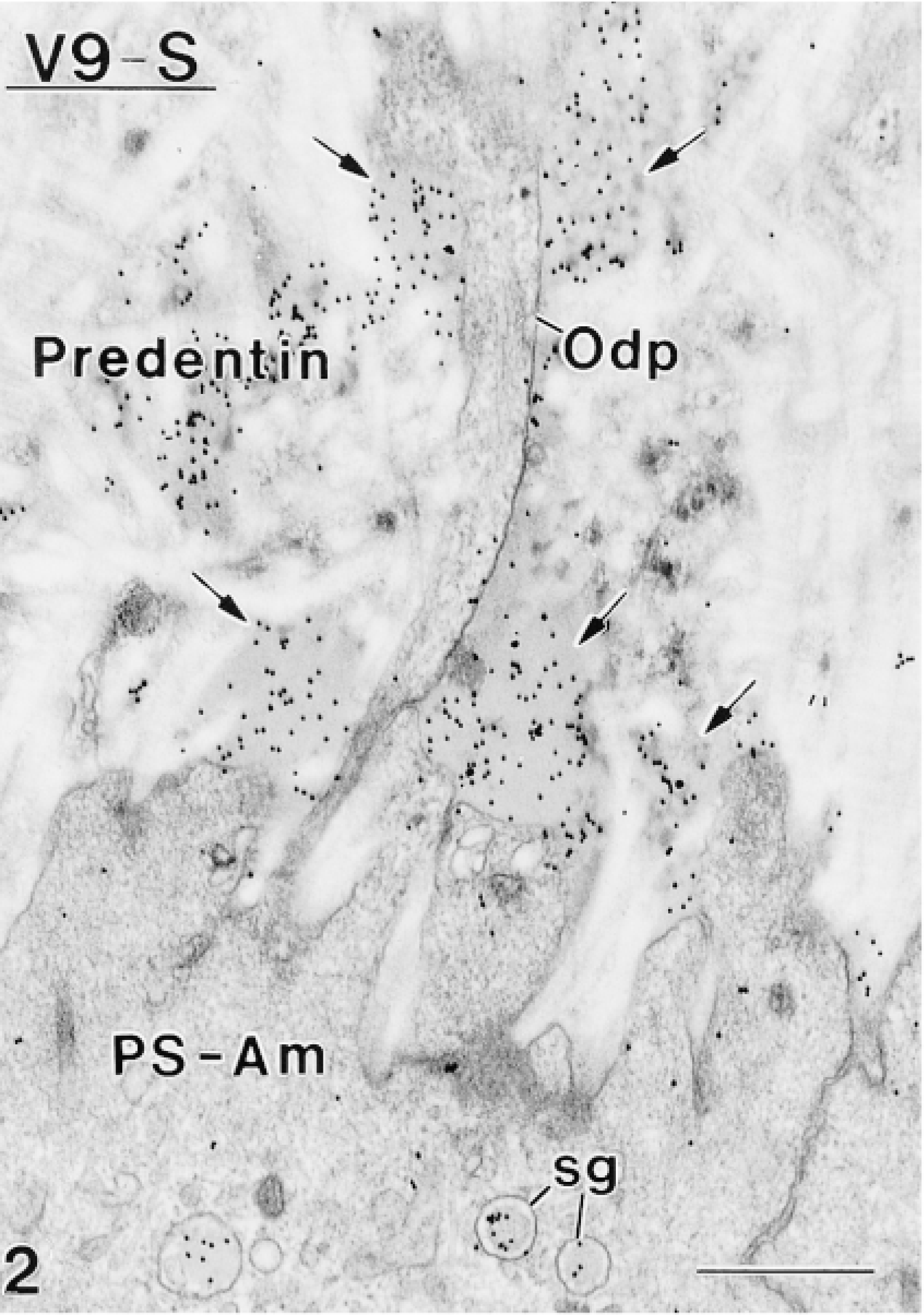
Four antibodies against vimentin were tested on plastic sections of rat incisors for their ability to immunoreact with odontoblasts. None showed any significant immunolabeling in these cells. However, some of these antibodies were found to localize over intra- and extracellular compartments of ameloblasts previously shown to contain amelogenins (Nanci et al. 1987). Tissue fixation with 2.5% glutaraldehyde yielded little or no immunoreactivity over these compartments, whereas labeling was observed with both 1% glutaraldehyde and 4% paraformaldehyde + 0.1% glutaraldehyde. There were no apparent differences in the distribution of binding sites in calcified and decalcified specimens. Qualitative assessments of the density of labeling for various anti-vimentin antibodies are shown in Table 2. Antibody V9-S consistently resulted in the highest labeling density with all resins. As examples, the dense patches of enamel proteins among the collagen fibrils of the forming mantle predentin or adjacent to odontoblast processes were (Figures 1 and 2), the enamel layer (Figure 3), and patches of granular material between odontoblasts and/or ameloblasts (not shown) were intensely immunoreactive. Intracellularly, the Golgi apparatus, secretory granules, and multivesicular bodies of ameloblasts were labeled (Figures 2–4 and 6B). Occasionally, the cisternae of the rough endoplasmic reticulum were dilated and contained accumulations of immunoreactive material (Figure 5).
Immunogold labeling a over secretory stage enamel in sections incubated with different antibodies to vimentin

intensities are expressed relatively as: + + + +, intense; + + +, strong; + +, moderate; +, weak; ±, very weak; -, none.
V9-S, clone V9 from Sigma; V9-BM, clone V9 from Boehringer Mannheim; V-3B4, clone 3B4 from Boehringer Mannheim; V-poly, polyclonal anti-vimentin from Sigma.
Tissues fixed in 1% glutaraldehyde and osmicated.
Tissues fixed in 4% paraformaldehyde +0.1% glutaraldehyde.
Antibody V9-BM gave almost no immunolabeling over ameloblasts, regardless of resin type. The density of gold particles over enamel was also highly variable in relation to embedding media and fixatives used (Table 2). Antibody V-poly gave only weak immunolabeling over the enamel matrix, whereas antibody V-3B4 showed no immunoreactivity over intra- and extracellular amelogenin-containing compartments (Table 2).
Sections incubated with AMEL showed labeling comparable to that of V9-S (Figures 6A and 6B). Competitive incubations with AMEL followed by V9-S resulted in significant abolition of the labeling normally obtained with V9-S (Figure 6C). Sections of Lowicryl K4M-embedded samples incubated with WGA followed by ovomucoid-gold likewise showed many gold particles over secretory granules and enamel (Figure 7A). Lectin binding sites were conspicuously concentrated at enamel growth sites (Figure 7A). However, incubation of sections with WGA before labeling with V9-S did not affect binding of the antibody to the tissue sections, as did AMEL (compare Figures 6C and 7B).
Unfixed secretory stage enamel proteins incubated with V9-S were unreactive. In constrast, purified vimentin and proteins extracted from cultured human fibroblasts showed strong immunostaining (Figure 8). However, when the enamel proteins were fixed with 1% glutaraldehyde before staining with the antibody, immunoreactive bands below 31 kD were revealed. The AMEL antibody did not significantly stain vimentin or any proteins in extracts from cultured human fibroblasts in fixed blots (Figure 8). Faintly stained bands were observed near 60 kD in blots of unfixed vimentin and enamel extracts but not in fibroblast extracts (Figure 8).
Discussion
Vimentin has been detected in various mesenchymal cells types of the tooth organ, including odontoblasts (Lyn et al. 1991; Seux et al. 1991; Inoue et al. 1992; Lombardi et al. 1992a; Davidson 1994; Papagerakis et al. 1999). The fact that we were unable to detect its presence in odontoblasts by high-resolution immunocytochemistry may be due to denaturation of antigenic sites during tissue processing and/or to its presence in amounts that are below the detectability threshold of postembedding colloidal gold immunolabeling. The latter possibility is likely because we have been able to obtain labeling in some periodontal ligament fibroblasts under similar tissue-processing conditions (unpublished observations). Indeed, it should be noted that even with pre-embedding colloidal gold labeling, only a very weak vimentin immunoreaction was observed in cultured dental follicle cells (Wise et al. 1992) and none was observed in odontoblasts.
Electron micrograph of rat incisor presecretory stage ameloblasts (PS-Am) at a position where they show short cytoplasmic processes (asterisks) extending into the forming mantle predentin. Patches of electron-dense material (arrows) are found near the apical surface of the ameloblasts. An odontoblast process (Odp) extends into a deep membrane invagination (arrowheads) at the apical surface of an ameloblast. (
Gold particles are present over enamel protein-containing compartments such as electron-dense patches (arrows) in mantle predentin, secretory granules (sg) in Tomes' processes (Tomes), and over enamel.
Weak but distinct labeling (arrowheads) is seen over the Golgi apparatus. Multivesicular bodies (mvb) are also immunoreactive.
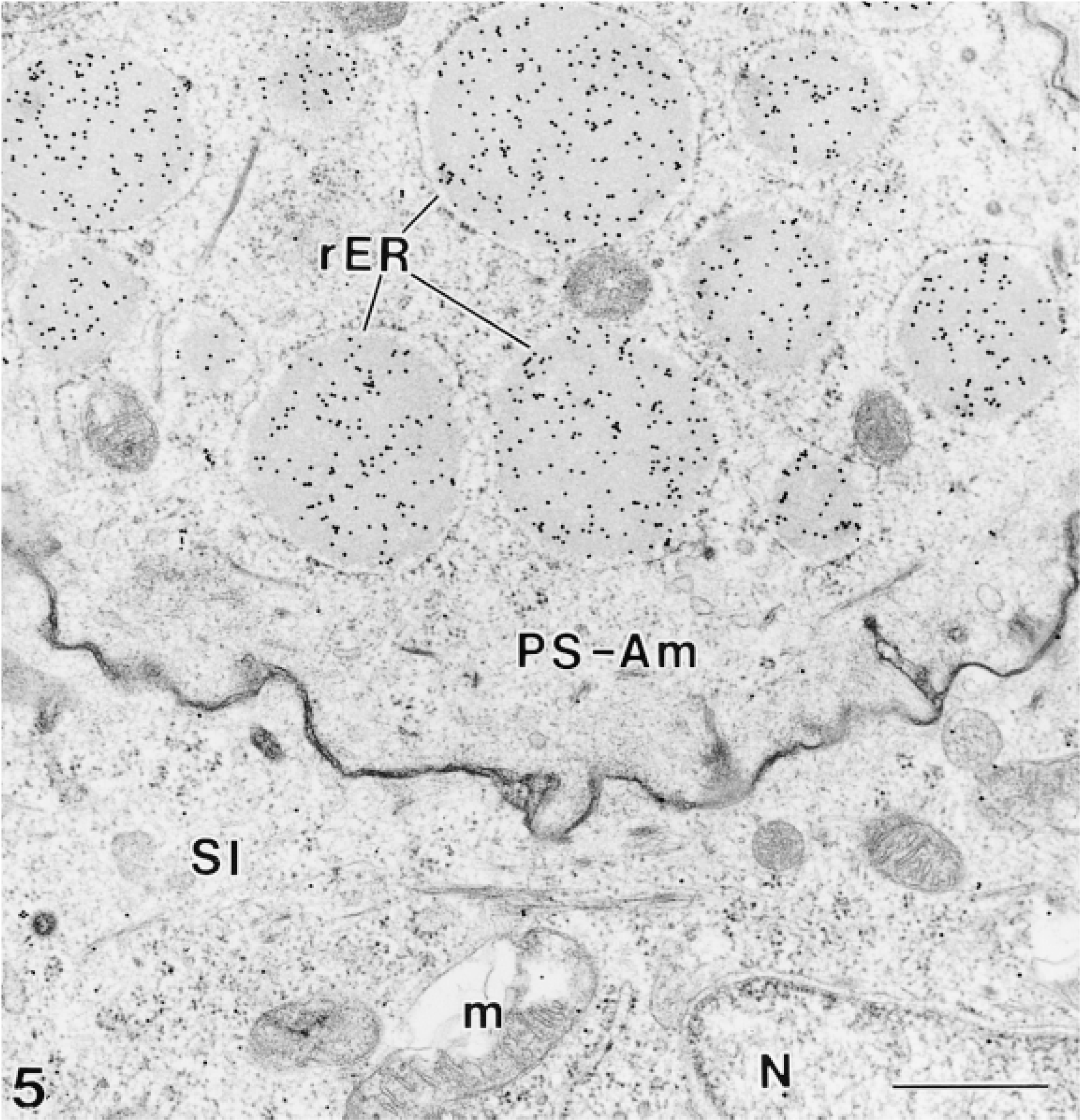
Many gold particles decorate the material found in distended rough endoplasmic reticulum (rER) profiles situated in the infranuclear compartment of some presecretory stage ameloblasts. N, nucleus; m, mitochondria; SI, stratum intermedium. Bars = 0.5 μm.
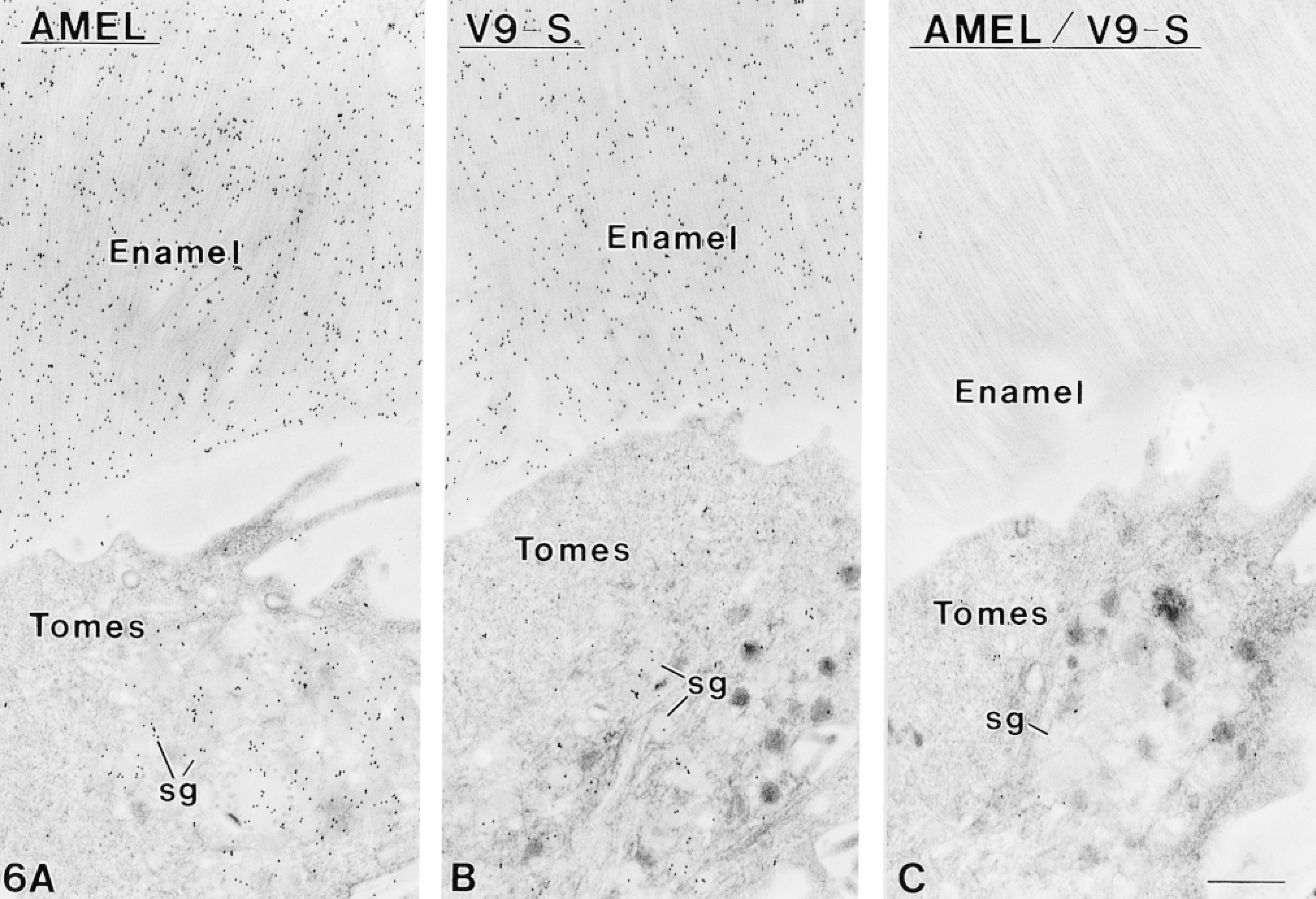
Immunocytochemical preparations of sections from nonosmicated Lowicryl-embedded secretory stage ameloblasts and enamel. (
A variety of monoclonal and polyclonal antibodies against enamel proteins have been used over the years to elucidate the process of amelogenesis (reviewed in Nanci and Smith 1992). The protein synthesis pathway for amelogenins has been well characterized, and these proteins have been immunodetected in the Golgi apparatus, secretory granules, and multivesicular bodies of ameloblasts. Extracellularly, they are found in enamel and in patches of material among collagen fibrils of the mantle predentin and along the basolateral surfaces of both ameloblasts and odontoblasts (reviewed in Nanci and Smith 1992). Comparison of the labeling obtained with V9-S, in particular, and with AMEL indicates that their labeling patterns are identical. Much to our surprise, V9-BM, a monoclonal from the same clone as V9-S, resulted in generally weaker labeling in Epon-embedded tissues, even after prolonged incubation times (see Table 2). Differences in their production and handling may in part explain the discrepancy in labeling density between the two vimentin antibodies. Irrespective of this difference, only V9-S and V9-BM labeled intracellular enamel protein-containing compartments. Antibody V-3B4 showed no labeling at all, and V-poly revealed very few binding sites only in LR White- and Lowicryl-embedded enamel. Therefore, the V9 clone antibody to vimentin appears to behave very much like an anti-amelogenin antibody. It is evident from the immunoblots that this effect is created solely as a consequence of tissue fixation. We therefore conclude that the reaction we observe in the tissue sections is not specific for enamel proteins but is still selective because it localizes only over enamel protein-containing compartments and shows no binding to other dental proteins, such as those found in dentin. Moreover, because V9-S binding can be blocked with AMEL, the epitopes recognized by these two antibodies must be very close. To evaluate the possibility that this abolishment was related to “crowding” (steric hinderance), blocking was also carried out with WGA, a lectin previously shown to bind to enamel (Nanci et al. 1989). Although the possibility of steric hinderance cannot be completely ruled out, it is unlikely that “crowding” plays a significant role in abolition of the labeling with V9-S, because the intense staining obtained with WGA does not affect binding efficiency of V9-S.
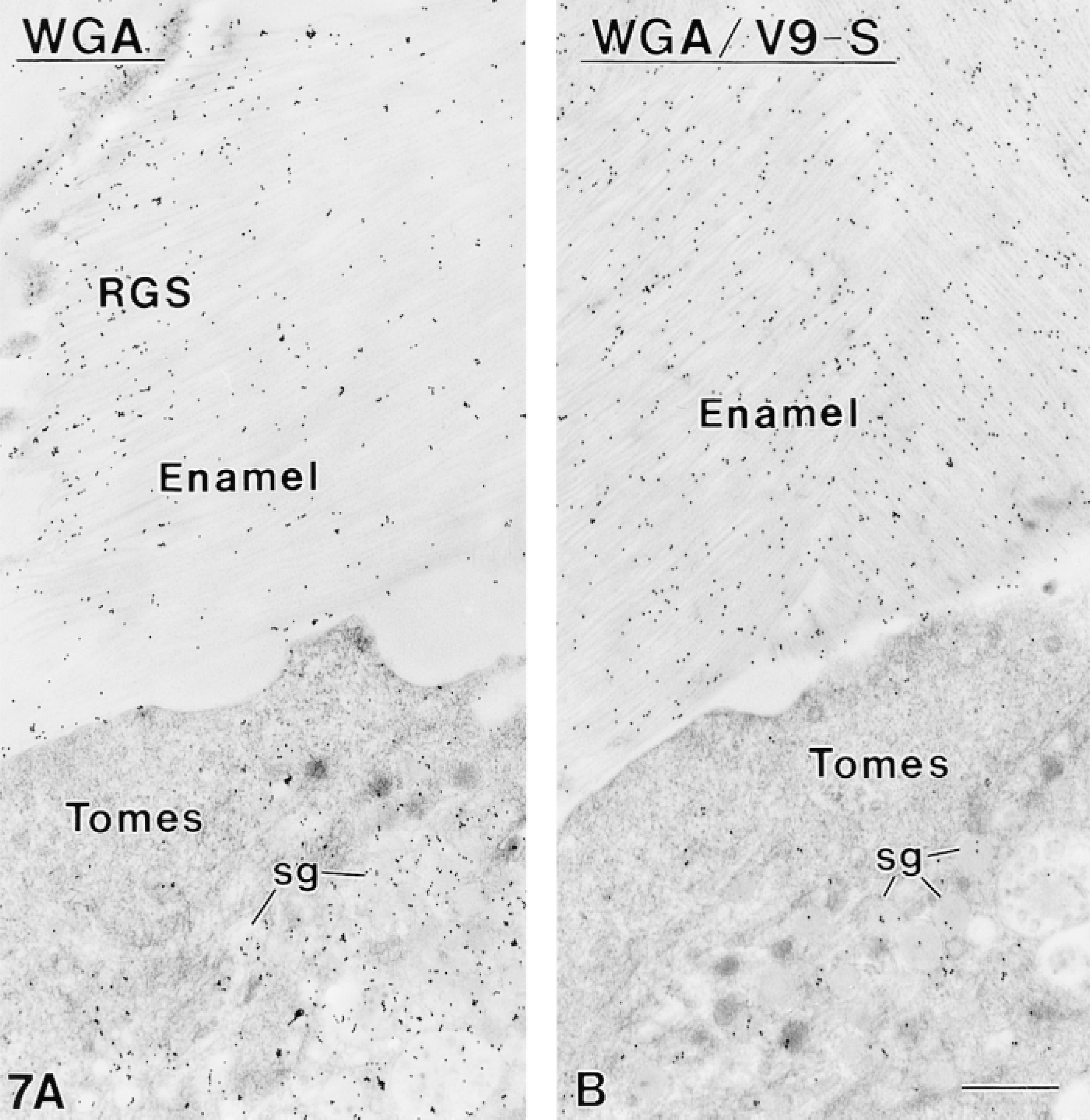
Sections from nonosmicated Lowicryl-embedded secretory stage ameloblasts and enamel. (
There are two major families of enamel proteins, amelogenins and nonamelogenins (reviewed in Robinson et al. 1995; Smith and Nanci 1996). Because the protein bands in the amelogenin stack contain both intact proteins and degradation forms of higher molecular weight proteins (see Smith and Nanci 1996), our data do not enable us to conclude unequivocally that V9-S binds exclusively to amelogenins. However, its pattern of labeling over enamel is clearly distinct from that obtained with antibodies to nonamelogenins, such as ameloblastin, and is very similar to that seen for amelogenin (reviewed in Nanci et al. 1998). Taken together, the data strongly suggest that V9-S probably binds to intact and/or degradative fragments of amelogenin. Because this antibody behaves similarly to anti-amelogenin antibodies, it may represent a useful tool for detecting and mapping of the distribution of this class of enamel proteins. Furthermore, because the labeling density increases when the concentration of enamel protein increases, e.g., in the dilated rough endoplasmic reticulum, the antibody could also be used for quantitative applications.
Monoclonal antibodies are directed against narrow determinants consisting of a few amino acids that can recur on several proteins and peptides. Therefore, it is not surprising that some monoclonals yield labelings on totally unrelated proteins from different tissues. In this regard, a monoclonal antibody against a monocyte-macrophage protein has similarly been found to bind to enamel proteins of less than 30 kD (Nakamura et al. 1991). A number of polyclonal and monoclonal antibodies to osteocalcin, a bone protein, have been shown to crossreact with epitopes on cultured skin fibroblasts (Bradbeer et al. 1994). Crossreactivity of an anti-human proinsulin antibody with both insulin and glucagon-secreting cells has recently been demonstrated (Bendayan 1995a). Most studies dealing with antibody crossreactivity, however, have been carried out on fixed tissues. It is therefore impossible to determine whether the reactivity observed was due to shared epitopes (crossreactivity) or to epitopes that were secondarily generated during aldehyde crosslinking. Crossreactivity can be ruled out in our present study because antibody binding to enamel proteins was obtained only after fixation, and because there is no significant homology between amelogenin and vimentin. Hence, whatever motif (amino acid sequence, posttranslational modification, and/or three-dimensional conformation) is being recognized by V9-S, it does not exist on native amelogenin and its formation is favored by tissue processing conditions (discussed in Bendayan 1995b). Although the possibility of false-positives clearly dictates that care should be taken in interpretation of immunolabeling, once the binding spectrum of an antibody has been clearly defined, as illustrated here, it is possible to take advantage of its selective characteristics to map the distribution of known molecules and even, perhaps, to help identify yet unknown ones.
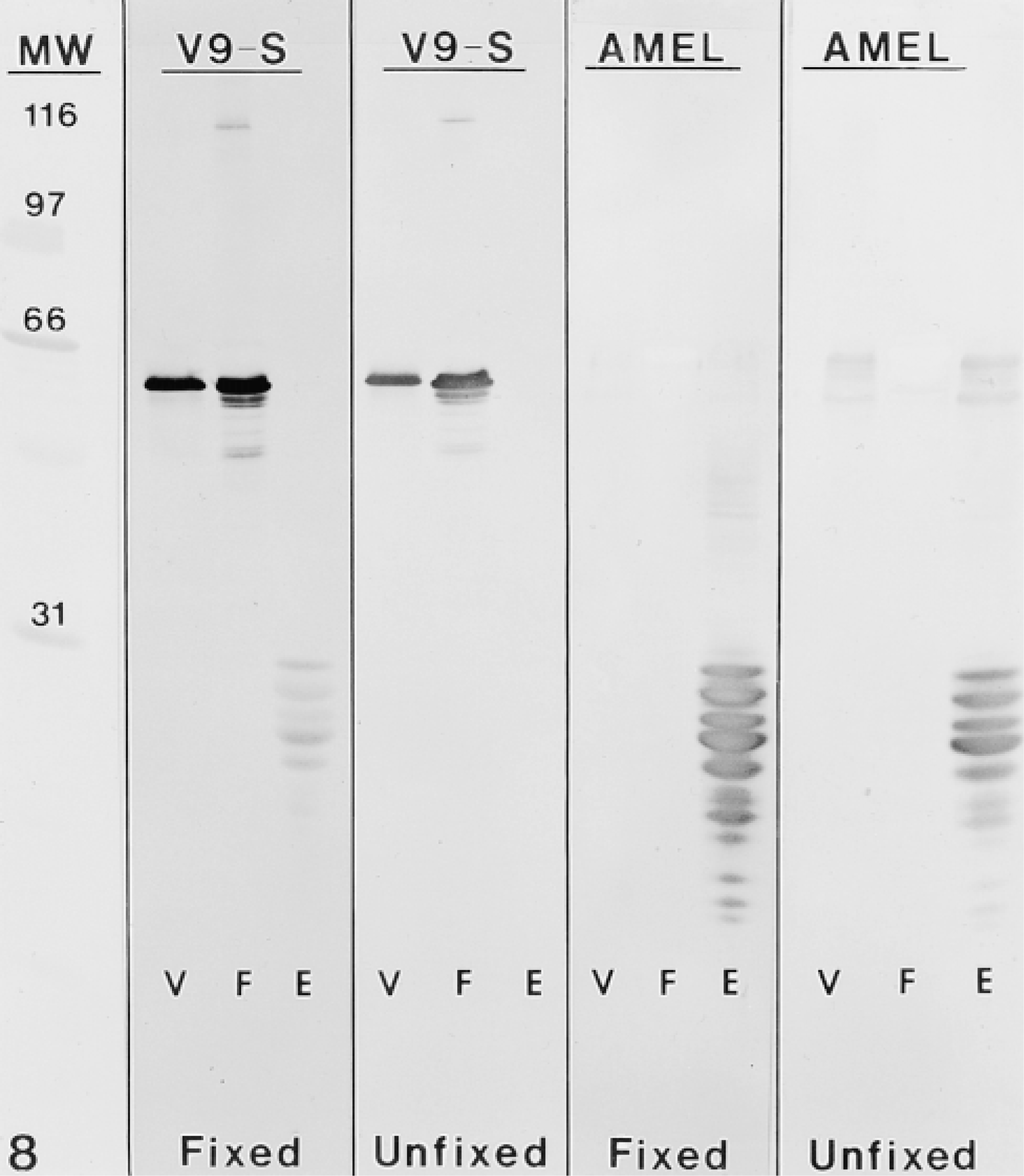
Immunoblots probed with V9-S and AMEL antibodies. The column at left shows standard broad-range molecular weight marker proteins (MW) stained with Ponceau. Each panel contains purified vimentin (V), human fibroblast lysate (F), and bulk protein extract from secretory stage enamel (E). After protein transfer, some nitrocellulose membranes were fixed with 1% glutaraldehyde before incubation with antibodies. Enamel proteins in the unfixed blots are not recognized by V9-S. However, proteins below 31 kD are revealed after fixation. Staining of vimentin and fibroblast extract samples was not significantly affected by fixation. Note also that both samples were not stained by AMEL. Whereas weak staining of some higher molecular weight bands with AMEL was abolished by fixation, staining of many enamel proteins below 31 kD was surprisingly intensified.
Footnotes
Acknowledgements
Supported by grants from Calcinfonden, the Danish Dental Association (KJ), and the Medical Research Council of Canada (AN, CES).