
Abstract
One of the unsolved key questions in neuroimmunomodulation is how peripheral immune signals are transmitted to the brain. It has been reported that the vagus might play a role in this regard. The underlying mechanism for this immune system-to-brain communication route is related to the binding of cytokines, such as interleukin (IL)-1β originating from activated immune cells, to their receptors in glomus cells of the vagal paraganglia. The existence of IL-1 receptor type I (IL-1RI) in vagal paraganglia has been proved. On the basis of these studies, a hypothesis is raised that the carotid body, as the largest paraganglion, might play a similar role to that of its abdominal partner. In this study we examined the distribution of IL-1RI in the carotid body by immunohistochemistry (IHC) and Western blotting techniques. The IHC results showed that almost all glomus cells in the carotid body displayed strong IL-1RI immunoreactivity. The IL-1RI-immunoreactive products were localized in the cytoplasm, nucleus, and cell membrane of the glomus cells. The Western blotting results also confirmed the existence of IL-1RI in both membranous and cytoplasmic elements of the carotid body. These results imply that the carotid body not only serves as a chemoreceptor for modulation of cardiorespiratory performance, as traditionally recognized, but also acts as a cytokine chemorereceptor for sensing immune signals.
A
It is generally believed that the informational molecules for immune-to-brain communication are proinflammatory cytokines originating from activated immune cells. Interleukin-1β (IL-1β) is one of the most likely mediators for immune-to-brain communication (Goehler et al. 1997). Two major classes of IL-1 receptor, type I and II, have been recognized (Bird and Saklatvala 1986; Dinarello 1991). Type II IL-1 receptor (IL-1RII) is a 68-kD glycoprotein and is localized mainly on B-cells, neutrophils, and bone marrow cells. Because of the short length of its cytoplasmic region, IL-1RII is unable to transmit signals and therefore acts as decoy target for IL-1 (Colotta et al. 1993). In contrast, type I IL-1 receptor (IL-1RI), expressed mainly on, e.g., T-cells, fibroblasts, endothelial cells, and hepatyocytes, is an 80–90-kD glycoprotein and mediates IL-1 effects. More attention has been paid to IL-1RI in the neuroimmunomodulation studies.
The mechanism(s) that enable the immune cytokines, such as IL-1β, to stimulate the peripheral endings of sensory fibers in the vagus is still unclear. Recently, Goehler et al. (1997) reported that paraganglia, which are located along the branches of the vagus in the peritoneum (Kummer and Neuhuber 1989), might play a possible role in this mechanism. Paraganglia contain two kinds of cells: type I cells (or glomus cells) and type II cells (or sustentacular cells) (Tischler 1987). Type I cells from synapses with the afferent fibers of the vagus (Berthoud et al. 1995: Goehler et al. 1997) and are capable of synthesizing catecholamine (Fidone and Gonzalez 1987; Berthoud et al. 1995). Goehler et al. (1997) found that a biotinylated IL-1R antagonist could bind to the glomus cells of vagal paraganglia. We also have demonstrated the existence of IL-1RI in vagal paraganglia by using immunohistochemistry (IHC) (Wang et al. 2000).
According to anatomic criteria, the carotid body, which is well known for its chemoreceptive function, is the largest paraganglion in the body. The carotid body in mammals has a similar configuration to that of the abdominal paraganglion (Fidone and Gonzalez 1987) and is also composed of two types of cells, type I cells (glomus cells, also called principal cells) and type II cells (sustentacular cells, also called dendritic cells). Like its peritoneal partner, the carotid body is also innervated by the sensory fibers of a cranial nerve (the carotid sinus nerve, a branch of the glossopharyngeal nerve). Given these structural similarities, a hypothesis is raised that the carotid body also plays a role in the sensation and transmission of immune stimulation. As a first step, we examined the presence of IL-1RI in the carotid body by using IHC and Western blotting techniques.
Materials and Methods
Adult male Sprague-Dawley rats weighing 200–300 g were used (n=29). The animals were housed under a 12:12 light:dark cycle and had access to laboratory chow and water ad libitum until they were deeply anesthetized with sodium pentobarbital (60 mg/kg IP).
Immunohistochemistry for Light Microscopy
Rats (n=14) were transcardially perfused with 100 ml of 0.9% saline followed by 500 ml of 4% paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4, at 4C. After perfusion the carotid bodies, including the bifurcation of the carotid artery, were removed and put in 4% paraformaldehyde for 1 hr at 4C before being moved into 20% sucrose in 0.1 M PB overnight at 4C for cryoprotection. Fifteen-μm-thick frozen sections were cut with a cryostat and mounted on gelatinized slides. After being dried at room temperature (RT), the sections were blocked with 5% normal goat serum, 1% bovine serum albumin (BSA), and 0.3% Triton in 0.01 M PBS at RT for 50 min.
For double immunofluorescent staining, the sections were incubated with a mixture of anti-IL-1RI polyclonal antiserum raised from rabbit (1:300; Santa Cruz Biotechnology, Santa Cruz, CA; or 1:100, Research Diagnostic, Flanders, NJ) and anti-tyrosine hydroxylase (TH, a marker for glomus cells) (Fidone and Gonzalez 1987) monoclonal antibody (MAb) raised from mouse (1:500; Sigma, St Louis, MO) in 0.01 M PBS (pH 7.4) containing 0.3% Triton and 1% normal goat serum overnight at RT. After three rinsings, the sections were incubated with a mixture of goat anti-rabbit IgG-Alexa 488 (1:400; Molecular Probes, Eugene, OR) and goat anti-mouse IgG-Texas Red (1:400; Molecular Probes). After 2-hr incubation at RT, the slides were washed and coverslipped with 50% glycerol in 0.1 M PB. The fluorescent sections were observed and photographed with a fluorescent microscope (BX 60; Olympus, Tokyo, Japan) or a confocal laser scanning microscope (CLSM) (FV-300; Olympus).
Several controls were conducted to confirm the specificity of IL-1RI immunoreactivity. First, as noted above, the anti-IL-1RI polyclonal antiserum from different sources (Santa Cruz Biotechnology and Research Diagnostic) was used. Second, omission controls were performed. The primary anti IL-1RI antibody was replaced by normal rabbit serum or 0.01 M PBS (pH 7.4) containing 0.3% Triton and 1% normal goat serum, and then the sections were incubated with goat anti-rabbit IgG-Alexa 488. The anti-TH MAb was replaced by normal mouse serum or 0.01 M PBS (pH 7.4) containing 0.3% Triton and 1% normal goat serum, and the sections were incubated with goat anti-mouse IgG-Texas Red. Finally, to ensure that there was no crossreactivity between the primary antibodies and the non-related second antibodies in double fluorescent staining, some sections were incubated in goat anti-rabbit IgG-Alexa 488 after the incubation in anti-TH MAb raised from mouse or in goat anti-mouse IgG-Texas Red after the incubation in anti-IL-1RI polyclonal antiserum raised from rabbit.
For IHC staining with the avidin-biotin complex (ABC) method, the rabbit-raised primary antibodies against IL-1RI from both companies (Santa Cruz Biotechnology and Research Diagnostic) were used. The biotinylated goat antirabbit antiserum and Elite ABC kit were from Vector Labs (Burlingame, CA). The standard staining protocol for the ABC method was used and has been reported elsewhere (Wang et al. 2002). The omission control was the same as above.
Western Blotting
Rats (n=15) were transcardially perfused with 100 ml of 0.9% saline under deep anesthesia with sodium pentobarbital (60 mg/kg IP) to wash the blood out. Bilateral carotid bodies with some connective tissues were removed. At first, about 60 mg of tissue was lysed in 150 μl of ice-cold Tris-buffered saline (TBS) containing 2% protease inhibitors (Boehringer Mannheim; Mannheim, Germany) by using a glass homogenizer and was left to stand on ice for 30 min to release the cytoplasmic proteins. Insoluble materials were separated by centrifugation at 10,000 X g for 10 min. The supernatant fluid, the total cytoplasmic lysate, was collected. One hundred μl of cold RIPA containing 2% protease inhibitors was added to the insoluble materials. These membranous elements were lysed by using a glass homogenizer and were left to stand on ice for 30 min to release membranous cell proteins before clarification by centrifugation at 10,000 X g for 10 min. The supernatant fluid, the membrane lysate, was collected. Both supernatants were mixed with one-quarter volumes of 4 X SDS-PAGE sample buffer (Sigma) and heated at 100C for 5 min. Before electrophoresis, each sample was clarified again by centrifugation at 10,000 X g for 10 min. Electrophoresis was carried out by SDS-PAGE using 10% polyacrylamide according to standard protocols. Then the proteins in PAG were transferred to nitrocellulose membranes (Schleicher & Schuell; Dassel, Germany). Before membrane blocking and incubation in primary antibody, these membranes were stained with Ponceau solution to show the protein bands of both the marker and the samples. The following steps were performed according to the protocol of the BM chemiluminescence Western blotting kit (Roche Molecular Biochemicals; Mannheim, Germany). The nitrocellulose membranes were blocked in 1% blocking solution (Roche Molecular Biochemicals) under shaking for 1 hr at RT to block nonspecific binding of antibody after being washed twice with TBS. Next, we incubated the membranes for 1 hr at RT with rabbit anti rat IL-1RI primary antibody (Research Diagnostic) diluted in 0.5% blocking solution with gentle shaking. After being washed twice in TBST (TBS containing 0.05% Tween-20) for 10 min each and in 0.5% blocking solution for 10 min, the membranes were incubated with POD-labeled reconstitute second antibody (40 mU/ml; Roche Molecular Biochemicals) diluted with 0.5% blocking solution for 1 hr. The membranes were washed with TBST three times for 10 min each. After exposing the membranes to the detection reagents for 60 sec, we placed a sheet of X-ray film onto the membranes in an X-ray film cassette and exposed the film for 60 sec, in a dark room. Then we developed the exposed film immediately in developer and fixed it in fixing solution in tanks under a red safelight. The black bands in the film and the marker stained by Ponceau solution were scanned into the computer and processed by Photoshop without changing the results.
Omission controls were done in which the primary IL-1RI antibody was replaced by 0.5% blocking solution or normal rabbit serum.
Results
Immunocytohistochemistry
In the fluorescent staining sections, the glomus cells of the carotid body were gathered in small cell groups and scattered in the connective tissue stroma (Figure 1D). There was strong IL-1R I-like immunoreactivity throughout the body (Figure 1A). Although the intensity of the immunostaining varied from cell to cell, almost all the glomus cells were intensely immunolabeled with IL-1RI (Figures 1E and 1H). These type I cells were ovate and about 10 μm in diameter. The cell membrane and cytoplasm of the glomus cell were immunostained intensely. In addition, their nuclei were immunostained, although the staining was weak. There was also strong TH-like immunoreactivity throughout the carotid body (Figure 1B). TH-positive products appeared in the cytoplasm of glomus cells but not in the nucleus (Figures 1F-1H). In double staining, all of the TH-positive cells were labeled with anti-IL-1RI antibody (Figures 1C, 1G, and 1H). Conversely, very few IL-1RI-positive glomus cells, judged from their morphology, were negative for TH (Figures 1G and 1H). Blood vessels were stained only by IL-1RI (Figures 1G and 1H). Some tissues between the glomus cells were labeled only with IL-1RI, but we could not identify them as sustentacular cells, connective tissue cells, or blood vessels under the present conditions. The staining results by the anti-IL-1RI antibodies from Santa Cruz Biotechnology (Figures 2A and 2B) were the same as those from Research Diagnostic.
No immunoreactive products were observed in the omission sections in which the primary anti IL-1RI antibody and anti-TH antibody were replaced by normal rabbit serum (Figure 1I), normal mouse serum (Figure 1K), or 0.01 M PBS containing 0.3% Triton and 1% normal goat serum (Figures 1J and 1L). No crossreactivity was seen between rabbit-raised primary IL-1RI antibody and anti-mouse second antibody, and vice versa.
By using ABC methods, the same staining results by using the anti IL-1RI antibodies from Research Diagnostic were obtained (Figures 3A and 3D). There was strong IL-1RI-like immunoreactivity throughout the body. The positive glomus cells clustered among the connective tissues. No immunoreactive products were observed in the omission control sections in which the primary IL-1RI antibody was replaced by normal rabbit serum (Figure 3B) or 0.01 M PBS containing 0.3% Triton and 1% normal goat serum (Figure 3C).
Western Blotting
In the cell membrane and cytoplasm lanes, specific bands were observed in the position of 80 kD, in agreement with the molecular weight of IL-1RI (Figure 4). There was no band in the corresponding position for the two omission controls. These results not only confirmed the specificity of the anti-IL-1RI antibody but also suggested that both the cytoplasm and the cell membrane contained IL-1RI.
Discussion
The carotid body mainly contains two kinds of cells: type I cells (glomus cells) and type II cells (sustentacular cells) (Fidone and Gonzalez 1987). Type I cells contain many membrane-bound cytoplasmic granules, and they synthesize and store catecholamines and a variety of neuropeptides such as dopamine, acetyl-choline, epinephrine, enkephalin, vasoactive intestinal polypeptide (VIP), neuropeptide Y (NPY), calcitonin gene-related peptide (CGRP), cholecystokinin (CCK), calcitonin, galanin, and substance P (SP). Some of these substances have been shown to co-exist in the glomus cells (Tan and He 1982; Fidone and Gonzalez 1987; Tischler 1987; Heym and Kummer 1989; Ichikawa and Helke 1993; Wang et al. 1993; Kim et al. 2001). The carotid body receives sensory innervation from the carotid sinus nerve (a branch of the glossopharyngeal nerve), efferent sympathetic innervation from the superior cervical ganglion, and parasympathetic innervation from the vagus (Fidone and Gonzalez 1987; Berger 1980). Classically, the carotid body has been confined to a chemoreceptor responding to changes of PO2, PCO2, and pH in blood, and thus plays a modulating role in the control of circulation and respiration (Gallego et al. 1979; Iturriaga et al. 1991; Kusakabe et al. 1993; Kim et al. 2001; Vidruk et al. 2001). Furthermore, it has become clear that the carotid body is also sensitive to temperature and osmotic changes (McQueen and Eyzaguirre 1974; Yamashita 1977; Gallego et al. 1979; Iturriaga et al. 1991). However, whether the carotid body is involved in neuroimmunomodulation is still unknown.
IL-1 is a prototype of the pro-inflammatory cytokines that induce expression of a variety of genes and the synthesis of several proteins which, in turn, induce acute and chronic inflammatory changes (Dinarello 1991). In recent studies it has been shown that the vagus may play an important role in transmitting peripheral immune information to the brain (Gaykema et al. 1995,1998; Laye et al. 1995; Kapcala et al. 1996; Niijima 1996; Sehic and Blatteis 1996; Maier et al. 1998; Dantzer et al. 2000; Goehler et al. 2000), and IL-1 is one of the possible mediators in immune-brain communication. The vagal paraganglia probably act as tranducers to sense IL-1 in blood or tissues so that the immune signals in the periphery can be transformed into neural pulses and transmitted to the brain via the vagus. Goehler et al. (1997) have proved the distribution of IL-1 receptor type I in vagal paraganglion through binding of the IL-1 receptor antagonist in glomus cells of the vagal paraganglion. Our previous work also demonstrated the existence of the protein of IL-RI in vagal paraganglia by using IHC (Wang et al. 2000). These data suggested the ability of the vagal paraganglia to bind IL-1β in blood or tissues.
Anatomically, the carotid body is recognized as the largest paraganglion in mammals (Fidone and Gonzalez 1987). Given the similarities between the carotid body and the vagal paraganglion, it is natural for us to speculate whether the carotid body can play a role in transmitting peripheral immune signals to the brain. Therefore, we used IHC and Western blotting to detect the distribution of IL-1 receptor type I in carotid body in the present study, and what was exciting was that the IL-1 receptor type I did exist in the glomus cells of carotid body. This implies that the carotid body may not only act as a traditional chemoreceptor but may also act as a cytokine chemoreceptor for sensing immune signals. It may be an alternative route in transmitting peripheral immune signals to the brain.
An interesting phenomenon in the present study is that the nucleus of the glomus cells was immunostained by the anti-IL-1RI antibody. Mizel et al. (1987) reported that when they studied the dynamics of IL-1 binding and internalization in T-cells and fibroblasts they found an accumulation of relative high levels of IL-1 in the nucleus. This raised the interesting possibility that IL-1 might interact in a highly specific manner not only with cell surface receptors but also with potentially important nuclear receptors. Curtis et al. (1990) also reported that in the murine T-cell line EL-4 IL-1 bound to its receptor on the cell surface and was translocated to the nucleus as an undegraded IL-1R complex. Our result also implies the presence of a nuclear receptor for IL-1 in the carotid body.
The mechanisms of origin of nerve impulses in the carotid body chemoreceptor are still unclear. Three structures have been proposed as the primary site for chemoreception: sustentacular cells, sensory terminals, and glomus cells (Tan and He 1982; Fidone and Gonzalez 1987; Eyzaguirre et al. 1989). According to our result, showing that IL-1 binding sites were strongly present in glomus cells in the carotid body, the glomus cells are possible primary receptor sites for immune chemoreception, although we cannot deny the alternative possibility. It is possible that when type I cells receive a stimulus they release the active substance they contain, such as dopamine, which may either modulate type I cells themselves or stimulate the neural endings and then convey the signals to the brain (Tan and He 1982; Fidone and Gonzalez 1987). It is also possible that such a stimulus influences glomus cells via sustentacular cells or directly excites sensory nerve terminals and then influences the glomus cells because of the existence of reciprocal synapses between glomus cells and sensory nerve terminals (Tan and He 1982; Fidone and Gonzalez 1987). In addition, Kim et al. (2001) reported that low oxygen caused a release of substance P in the rabbit carotid body. Hart et al. (1993) proved that IL-1 induces SP gene expression in cultured rat superior cervical ganglion. As to the role of IL-1β in the carotid body, we speculate that IL-1β may induce depolarization of cell membrane of type I cells in the carotid body and evoke the release of neurotransmitters bind to their receptors in carotid sinus nerve endings to induce an action potential. In addition, it is also possible that IL-1β would not evoke an action potential in afferent nerve fibers but would act merely as a modulator to regulate activities of type I cells themselves. It may also function to strengthen or weaken the effects of other transmitters on the discharge of the carotid sinus nerve caused by other stimulation. Further electrophysiological and pharmacological experiments are needed.
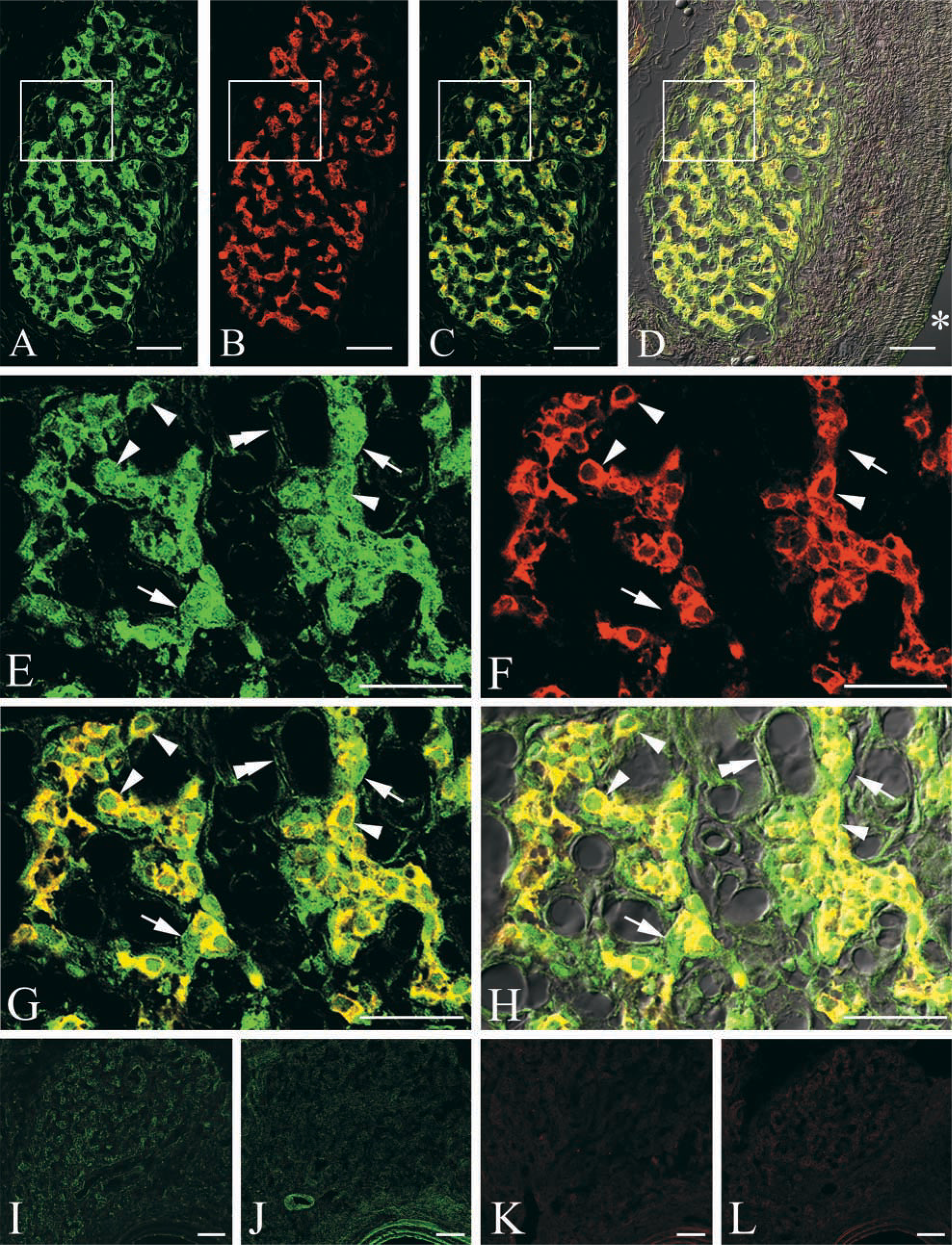
Photomicrographs taken with a confocal laser scanning microscope (CLSM) showing the IL-1RI (green) (
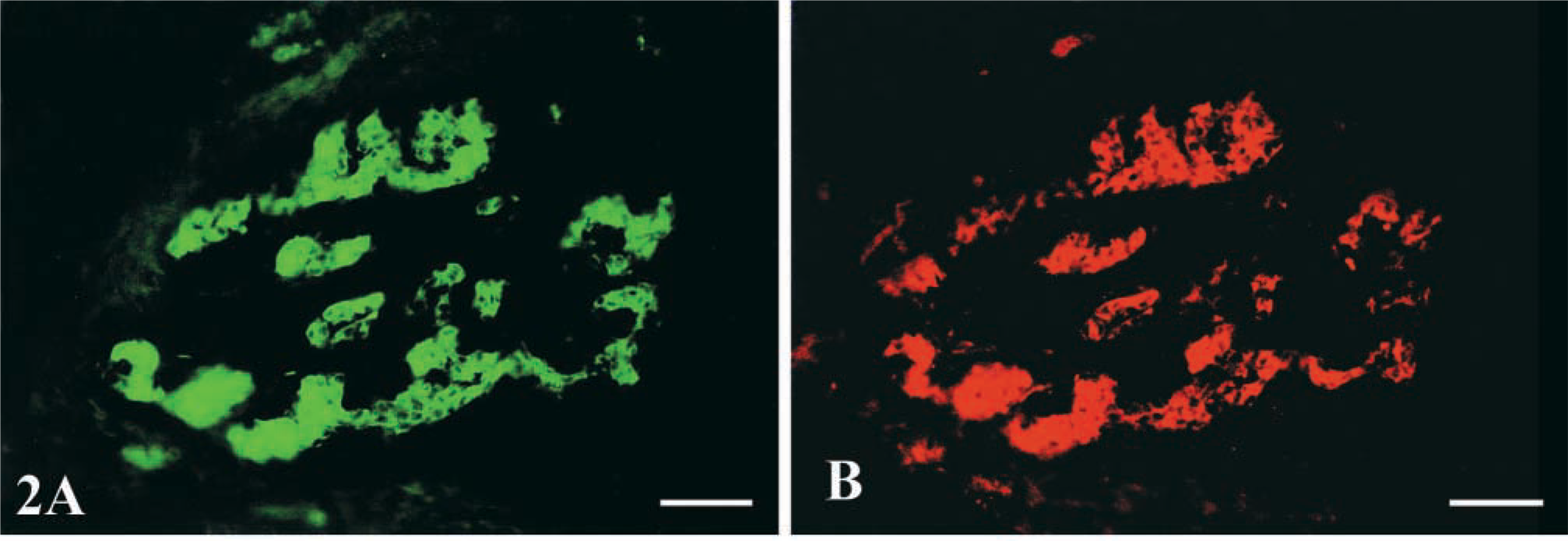
Photomicrographs taken with the BX-60 fluorescent microscope showing the IL-1RI (green) (
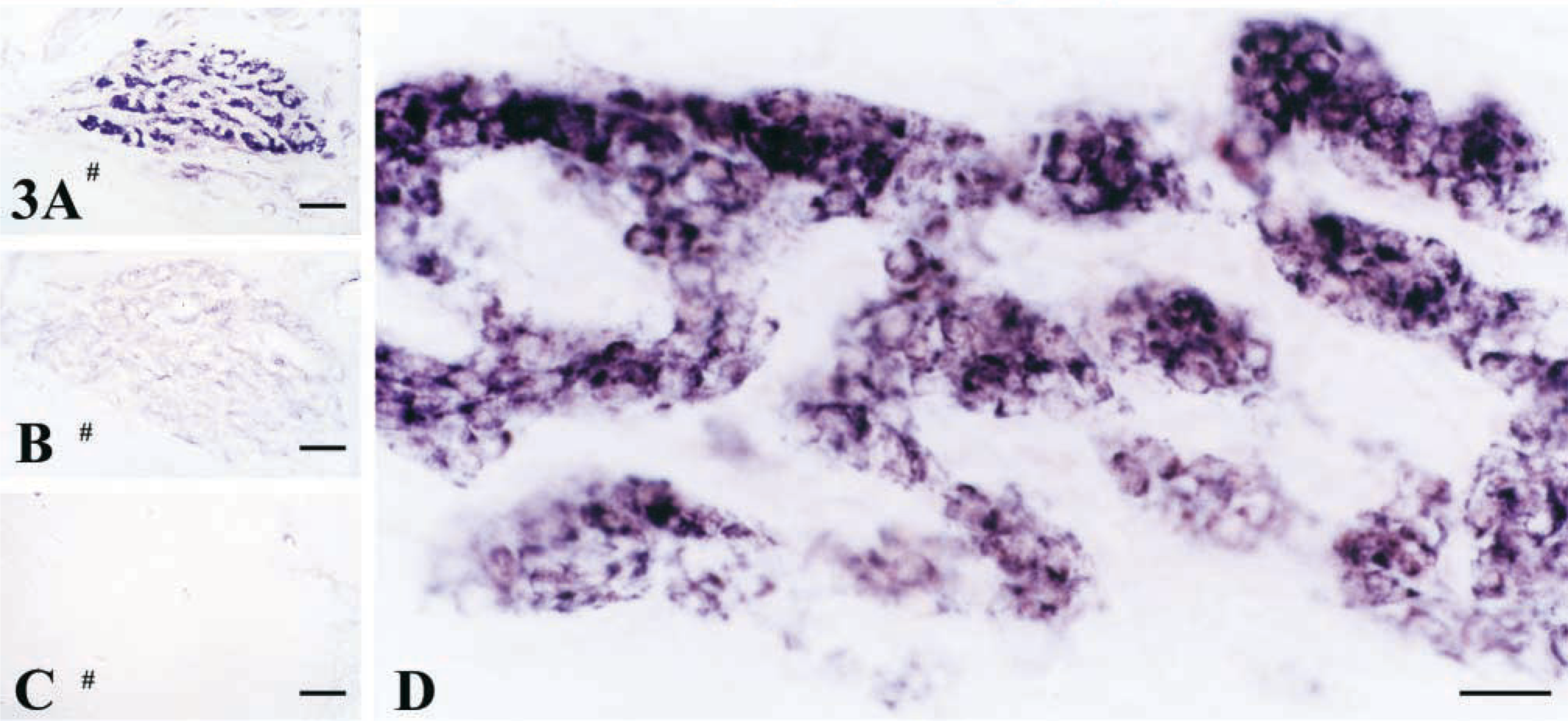
Photomicrographs showing the IL-1RI-like immunoreactivity stained with the ABC method. (
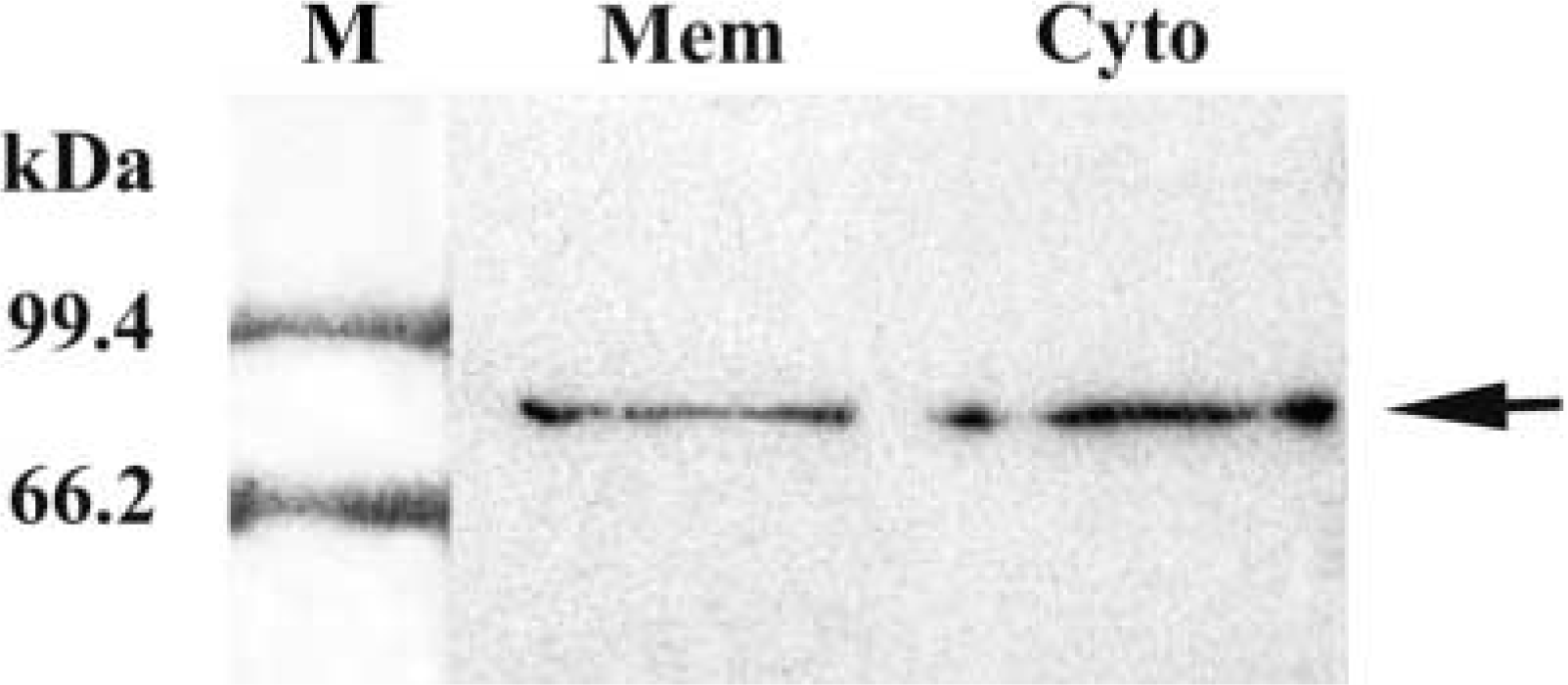
Western blotting analysis of IL-1RI expression in the cytoplasm and the cell membrane of the carotid body. Specific bands for IL-1RI appeared in the 80-kD position (arrows). M, marker; mem, cell membrane; Cyto, cytoplasm.
Footnotes
Acknowledgements
Supported by the Chinese National Nature Science Foundation (no. 39830130) and by the Key Teacher Supporting Program of the Chinese National Educational Administration.