
Abstract
Genetic immunization can be combined with hybridoma technology to generate high-affinity monoclonal antibodies (MAbs). A new anti-BCL-6 MAb (GI191E/A8) was produced by cloning full-length BCL-6 cDNA into a eukaryotic vector and delivering this into mouse epidermis using a helium gene gun. A comparative study was made of the specificity and the effects of formalin fixation on immunohistochemistry quality of GI191E/A8 and two other anti-BCL-6 MAbs. To evaluate its possible application to differential diagnosis of lymphomas, two tissue microarrays (89 diffuse large B-cell lymphomas and 24 B-cell chronic lymphocytic leukemia cases) were stained with GI191E/A8 and another anti-BCL-6 MAb produced by conventional means. Using GI191E/A8, the detection of BCL-6 protein was significantly increased, and its specificity was independent of formalin-fixation time. Using automatic quantified analysis, the correlation between the two anti-BCL-6 MAbs tested was identical in cases with overexpression or absence of BCL-6. In cases with intermediate BCL-6 protein expression, detection with GI191E/A8 was more sensitive. A significant association of higher BCL-6 expression and longer median overall survival times in diffuse large B-cell lymphomas was found. Using conventionally produced MAbs in the same patient group, the association was not significant.
D
One of the main advantages of GI is the fact that proteins are synthesized in vivo, resulting in production of proteins in their natural conformation. Therefore, GI-produced MAbs are mainly high affinity and recognize the native form of the protein (Barry et al. 1994). This method is also extremely useful in those cases where the protein is unavailable or difficult to produce, such as membrane-bound or secreted proteins (Nagata et al. 2003).
However, the successful application of this technique for generating MAbs has been hindered by technical problems such as low efficiency, with the consequence that very few hybridomas have been produced. This problem has been solved by giving mice that have previously been genetically immunized a final boost, using purified protein (Costagliola et al. 1998; Krasemann et al. 1999) or cells lines expressing the protein of interest (Nagata et al. 2003).
BCL-6 is a zinc-finger transcriptional repressor required for germinal-center (GC) formation, Ab-affinity maturation, and T-helper-2-mediated responses (Ye et al. 1997). The BCL-6 gene is frequently translocated and hypermutated in diffuse large B-cell lymphoma (DLBCL). Its expression is also important as an independent predictor of overall survival (OS) in DLBCL, where expression of BCL-6 is associated with improved OS, suggesting a GC origin for these tumors (Lossos et al. 2001). BCL-6 is expressed in all GC-derived B-cell tumors such as Burkitt's lymphoma, DLBCL, and follicular lymphoma. For this reason, the use of anti-BCL-6 MAbs has been an indispensable tool for the classification and diagnosis of B-cell neoplasms (Falini and Mason 2002).
The aim of our study was to demonstrate that GI is a viable and effective alternative method for production of MAbs for immunohistochemistry (IHC) on paraffin sections. For this purpose, we generated two anti-BCL-6 MAbs; one produced using GI and the second using glutathione S-transferase (GST) recombinant protein produced in bacteria. The specificity of the new GI-produced MAb was tested on paraffin sections using the pressure-cooking method of antigen retrieval and a variety of different buffers. Furthermore, the effect of formalin fixation on GI191E/A8 IHC quality was compared with two other anti-BCL-6 MAbs.
To confirm its possible application as a diagnostic marker, we examined BCL-6 protein expression using tissue microarrays (TMAs) for DLBCL and B-cell chronic lymphocytic leukemia (B-CLL), which are common examples of BCL-6-positive and -negative lymphoproliferative neoplasms, respectively. The results obtained using GI191E/A8 and ST42B/H7 MAbs were compared.
Materials and Methods
Mice
BALB/c mice were maintained under specific pathogen-free conditions and handled in laminar-flow isolation hoods in a barrier facility in the Animal Facility Unit of the Centro Nacional de Investigaciones Oncológicas (CNIO). All animal experiments were performed under the experimental protocol approved by the Institutional Committee for Care and Use of Animals, CEUCA no. 001/02.
Plasmid DNAs
The human full-length coding sequence of BCL-6 was inserted into the pc-DNA3 vector (Invitrogen; Carlsbad, CA), which contains a cytomegalovirus (CMV) promoter for gene expression and is commonly used for DNA immunization. The sequence of the BCL-6 encoding region was confirmed by DNA sequencing. The plasmid was amplified in Escherichia coli overnight in large-scale cultures and purified using the JETstar 2.0 endotoxin-free plasmid-purification kit (Genycell Biotech; Granada, Spain) according to the manufacturer's instructions. DNA was precipitated in ethanol, re-suspended in pyrogen-free water, quantified by spectrophotometry, and stored at 220C.
The ability of the constructed pCMV-BCL6 plasmid to mediate in vitro expression of BCL-6 was confirmed by calcium phosphate-mediated transient transfection of HEK-293T cells (American Type Culture Collection; Rockville, MD). Before immunization, DNA was diluted in pyrogen-free water to appropriate concentrations (2 μg/μl) for the preparation of Gene Gun (GG) cartridges.
Immunization and Production of MAbs by GI
Intradermal GI was performed using a Helios GG delivery system (Bio-Rad Laboratories; Hercules, CA). Two μg of purified plasmid DNA was precipitated onto 0.20 mg of 1-μm-diameter gold beads by CaCl2 in the presence of spermidine, according to the manufacturer's instructions (Bio-Rad Laboratories). Bullets (5 μg DNA/shot) were stored in a dry atmosphere at 4C in the dark, in the presence of desiccant pellets.
Mice were anesthetized with isoflourane gas for shaving off abdominal fur and for all immunization times. The helium-powered GG was used at 400 psi (2758 kPa) to deliver one shot to the inguino-abdominal skin of six mice. Immunizations were repeated five times at 14-day intervals, and serum samples were collected by puncture of the retro-orbital plexus after 40 and 70 days. Lesion discomfort or pain was not observed in the experimental mice after GG-immunization. Mouse serum was tested on frozen tonsil sections by IHC, showing that all mice developed strong and specific Ab responses. The last boost was performed using HEK-293T cells transfected with pCMV-BCL6 plasmid. Cells washed and resuspended in PBS (pH 7) were injected on day 80 (4 × 107 cells in 400 μl) into the mouse's intraperitoneal cavity (Nagata et al. 2003). Spleen cells were harvested 84 hr after the last cell boost for cell fusion with the NS-1 myeloma cell line (P3/NS1/1-Ag4-1) using the conventional technique (Mason et al. 1983). Initial screening was performed using an immunoperoxidase technique on cryostat sections of normal tonsil.
Western Blotting and Cell Lines
The GI191E/A8 MAb was characterized by Western blotting (WB) in a full range of cell lines and tissues using conventional technique (Sanchez-Aguilera et al. 2004). The non-Hodgkin's lymphoma-derived cell lines Toledo (DLBCL) and Ramos (Burkitt's lymphoma) were obtained from the American Type Culture Collection. The cell lines KARPAS-422 (DLBCL), DB (DLBCL), and REH (acute lymphoblastic leukemia) were obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ; Braunschweigh, Germany). The Burkitt's lymphoma-derived cell lines Mutu-1 (Gregory et al. 1990) and AKATA (Takada et al. 1991) were kindly provided by Dr. Miguel Campanero (Centro de Investigaciones Biológicas; Madrid, Spain).
Tissue Samples
Biopsy samples (five normal tonsils, 89 DLBCL, and 24 B-CLL lymphomas cases fixed in formalin and embedded in paraffin by conventional techniques) were obtained from the tissue archives of the CNIO Tumour Bank Network. All tissue samples were collected following the protocols established by the Tumour Bank Network and represented randomly chosen consecutive biopsies. Informed consent was obtained from all patients enrolled in the study under the supervision of the local ethical committees. All samples were centrally reviewed by a panel of pathologists and diagnosed using uniform criteria based on clinical, histological, immunophenotypical, and molecular characteristics (Harris et al. 2000). All B-CLL samples corresponded to tumoral lymph nodes involved by B-CLL/small lymphocytic lymphoma. Tissue sections (2–4 μm) were cut onto Dako slides (DakoCytomation; Glostrup, Denmark), incubated at 60C overnight, and deparaffinized twice using xylene (10 min, room temperature). Slides were hydrated in a series of ethanol solutions (100, 95, and 75% for 5 min each) and then washed with PBS.
To study the effect of formalin fixation on the quality of BCL-6 IHC, two samples of normal tonsil were cut into five pieces. Four pieces of each were fixed in 10% neutral buffered formalin (Panreac; Barcelona, Spain) for different times (12, 24, and 48 hr and 1 week, respectively), whereas the fifth piece was fixed for 1 hr in B5 fixative (Panreac). All samples were then processed and embedded in paraffin.
Antigen Retrieval
The tested retrieval solutions were 1 mM EDTA (pH 8) and 0.01 M citrate buffer (pH 6.5). Two liters of each retrieval solution were placed in a 6-liter stainless steel pressure cooker with an operating pressure of 0.09 bars at 117C (WMF Perfect Plus; Madrid, Spain) and brought to a boil on an electric hotplate without sealing the lid. The slides were placed in a metal rack and lowered into the boiling buffer. The pressure cooker was then sealed and brought to full pressure. Thereafter, it was depressurized using running tap water and the lid removed. An additional step of 10-min incubation with proteinase K (40×) diluted in proteinase K diluent (DakoCytomation) was applied to some of the slides previously treated with 0.01 M citrate buffer (pH 6.5).
Source of Antibodies
IHC studies were performed using three MAbs that are reactive on paraffin sections. The first was anti-BCL-6 (clone PG-B6p; DakoCytomation); the other two were MAbs against BCL-6 produced by the Monoclonal Antibody Unit of the CNIO: clone ST42B/H7 produced using recombinant GST-BCL-6 recombinant protein and clone GI191E/A8 produced by GI.
WB study was performed using the GI191E/A8 MAbs (diluted 1:2 in PBS with 10% fetal calf serum) and anti-α-tubulin (diluted 1:10.000) (Sigma Chemical Company; St. Louis, MO).
Antibody Purification
Tissue-culture supernatants of GI191E/A8 and ST42B/H7 clones were solubilized in 20 mM sodium phosphate buffer (pH 7). These solutions were filtered through 0.45-μm filters and applied to 5 ml HiTrap affinity columns (Protein G HP; Amersham Pharmacia Biotech, Uppsala, Sweden) previously equilibrated in 20 mM of buffered phosphate (pH 7), integrated with the AKTA Prime system (Amersham Pharmacia Biotech). The samples were loaded at 5 ml/min, and the column was washed with several column volumes of equilibration buffer. The samples were eluted in a single-step elution with 0.1 glycine-HCL (pH 2.7). IgG-containing fractions, as determined by a chromatogram, were pooled and recovered in tubes with 100 μl 1 M Tris-HCL, pH 9.0, as a safety measure to preserve the activity of labile IgGs. In this way, the final pH of the samples was approximately neutral. After sample recovery, the column was washed in several volumes of equilibration buffer. The fractions containing the purified IgG were dialyzed against PBS overnight at 4C. The final product concentration and the number of IgG molecules were determined from the ABS280nm using a spectrophotometer.
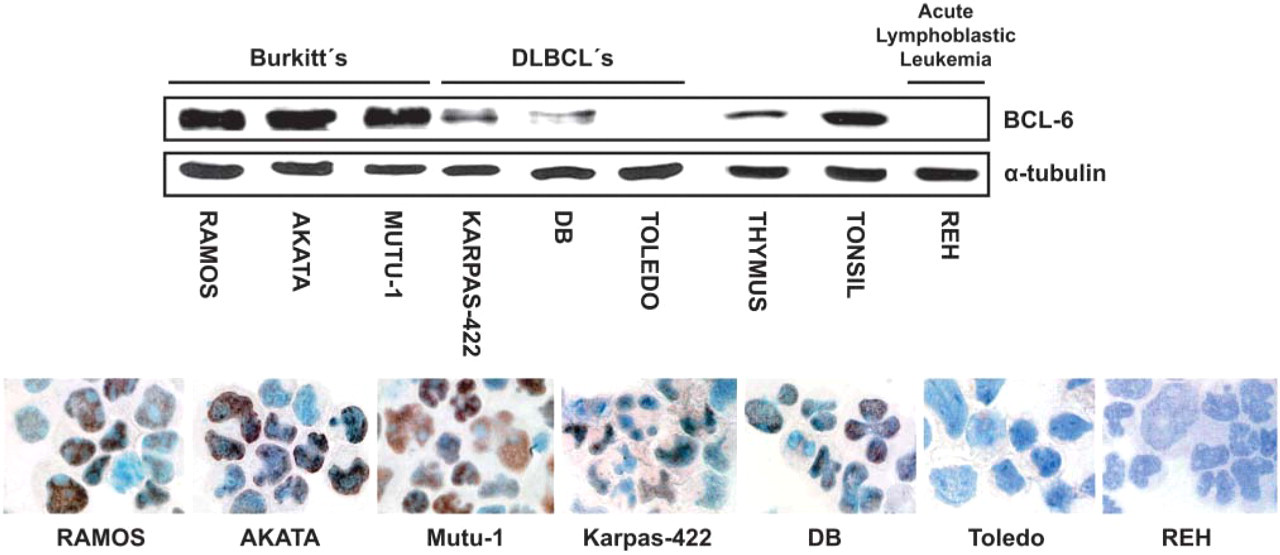
Correlation between Western blot (WB) and immunohistochemical staining for BCL-6 protein using the GI191E/A8 MAb. Top: WB analysis of lymphoid tissue and cell line samples for BCL-6 (upper lane) and α-tubulin (lower lane). Bottom: BCL-6 protein expression in cytocentrifuge preparations of lymphoid cell lines.
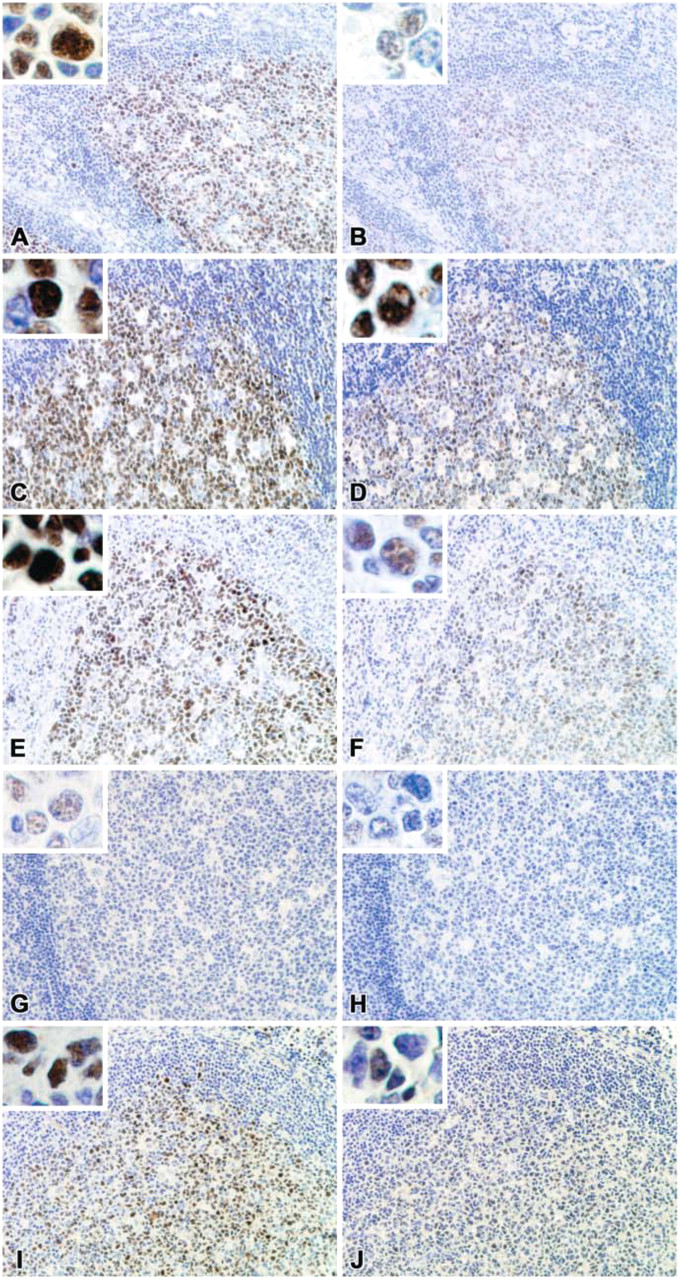
BCL-6 immunostaining of normal tonsil using different formalin-fixation times and B5 fixative with GI191E/A8 (
Comparative assessment of antibodies and retrieval solutions
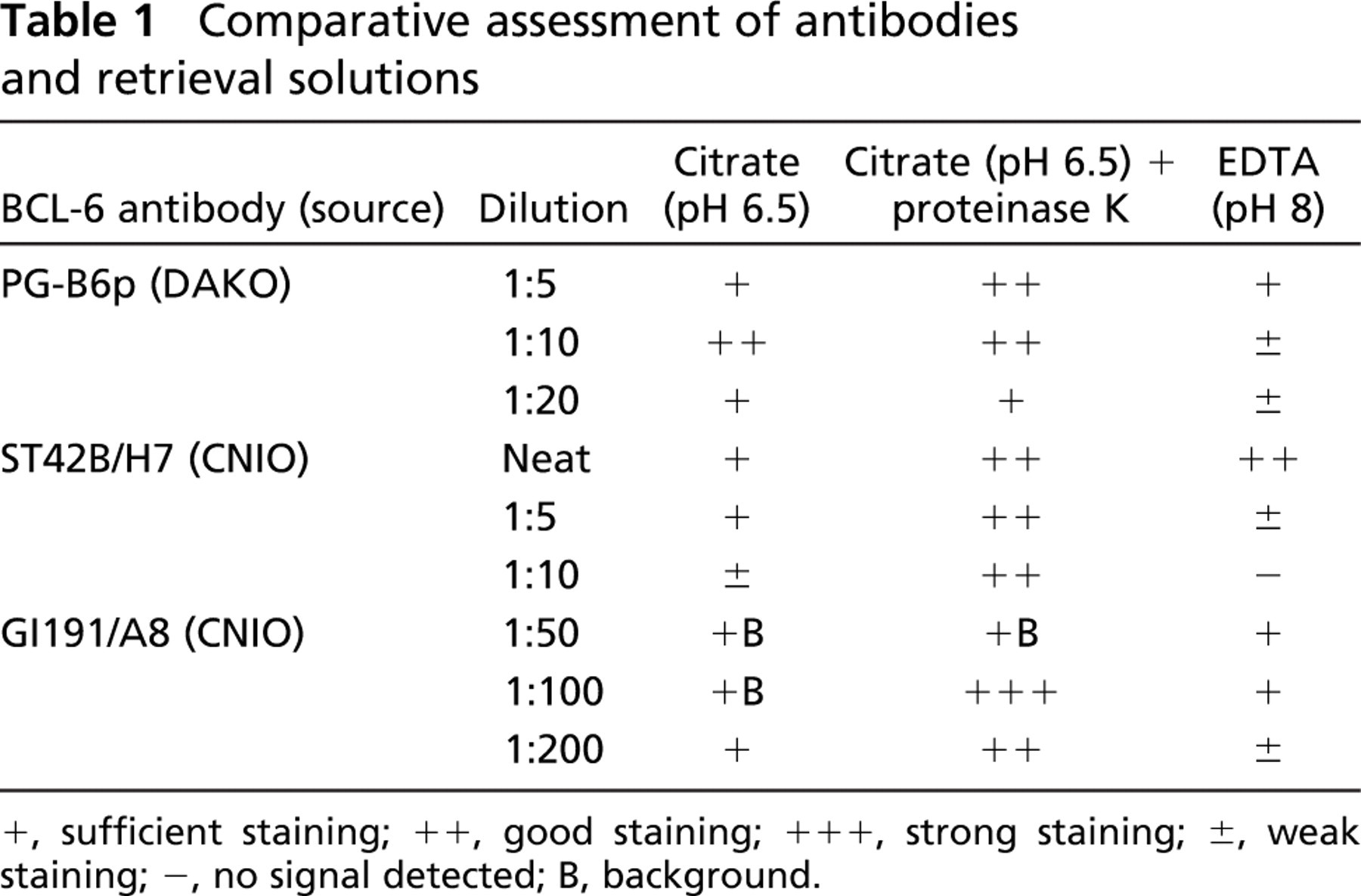
+, sufficient staining; ++, good staining; +++, strong staining; ±, weak staining; -, no signal detected; B, background.
Antibody-Affinity Assay
ELISA was performed to determine Ab affinities. Ninety-six-well microtiter plates were coated for 1 hr at 37C with the BCL-6 protein at 5 μg/ml. The plates were saturated with 2% dry milk powder, followed by incubation with serially diluted Ab (400, 200, 100, 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78, 0.39, and 0.2 nM). GI191E/A8 and ST42B/H7 clones were incubated in duplicate in the BCL-6-coated wells. After washing with PBS-Tween, horseradish peroxidase-conjugated goat anti-mouse Ig (DakoCytomation) was added. The plates were washed again and developed with horseradish peroxidase substrate kit solution (Bio-Rad) for 15 min. The reaction was stopped by adding 0.5% sodium dodecyl sulfate. Optical density (OD) was measured at 405 nm.
Immunostaining Techniques
Frozen tonsil tissue sections and cytocentrifuge preparations (Erber et al. 1984) were incubated for 30 min with MAb (supernatant or mouse serum), washed in PBS, and incubated with horseradish peroxidase-conjugated goat anti-mouse Ig (diluted 1:50 in PBS) (DakoCytomation). The peroxidase reaction was developed using diaminobenzidine (DakoCytomation) for 5 min and washed with distilled water. All sections were counterstained with hematoxylin before mounting.
Comparison of antibodies and fixation times
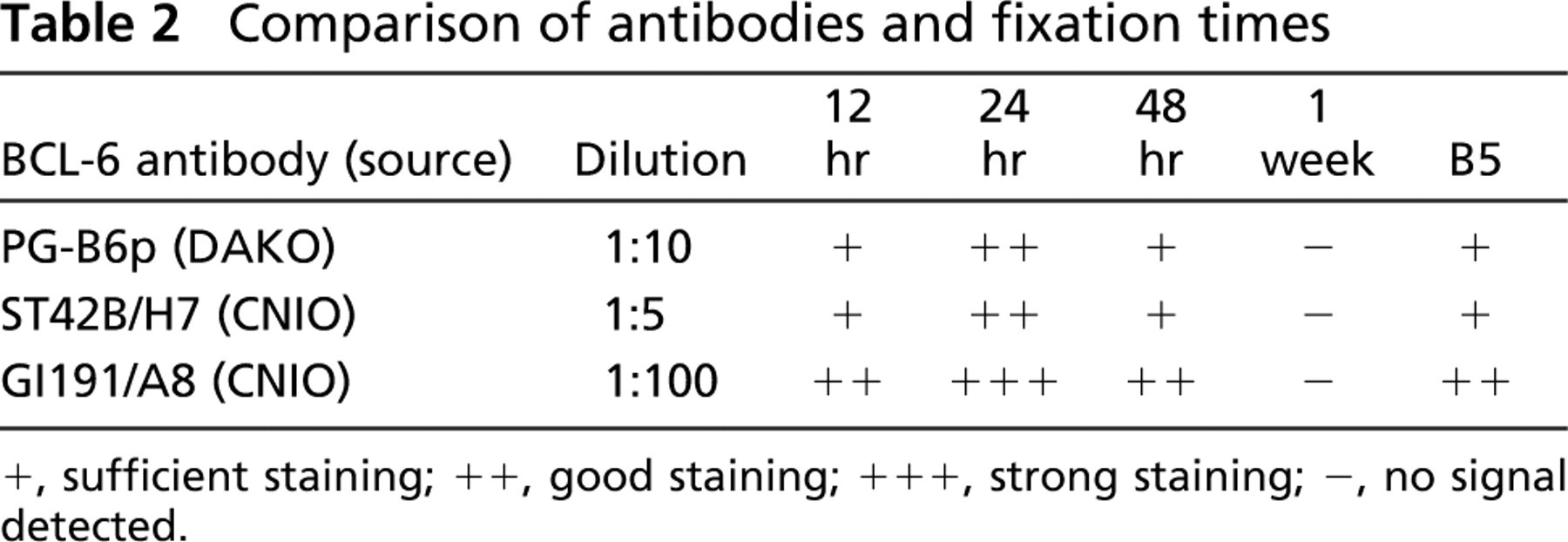
+, sufficient staining; ++, good staining; +++, strong staining; -, no signal detected.
Before staining paraffin-embedded sections, endogenous peroxidase was blocked, the slides were incubated for 40 min with the primary MAb, washed with PBS, and the immunodetection was performed with biotinylated anti-mouse secondary Abs (25 min), followed by peroxidase-labeled streptavidin (25 min) and diaminobenzidine chromogen as substrate. The immunostaining was performed using the Techmate 500 (Dako-Cytomation; Tucson, AZ) automatic immunostaining device. Reactives were supplied by DakoCytomation. Incubations either omitting the specific Ab or containing unrelated Abs were used as a control of the technique.
Analysis of BCL-6 Protein by TMAs and Quantification of BCL-6 Protein Expression
Both series of lymphomas (DLBCL and B-CLL) were processed using a Tissue Arrayer device (Beecher Instrument; Sun Prairie, WI). Briefly, all cases were histologically reviewed and the richest areas of tumoral cells were marked in the paraffin blocks. Two selected 1-mm-diameter cylinders were included for each case as previously described (Kallioniemi et al. 2001; Garcia et al. 2003). IHC was performed on these sections using the different Abs and procedures described above.
After immunostaining, TMAs were scanned and analyzed using the BLISS system (Bacus Laboratories, Inc.; Lombard, IL), which employs a three-charge-coupled device (CCD) red-green-blue (RGB) sensor optically coupled to a microscope, following the manufacturer's recommendations. Multiple × 40 images were scanned for each core, and the stored images were quantified using TMAscore v.2.0 image analysis software (Bacus Laboratories, Inc). For scoring the level of expression of BCL-6 (percentage of immunostained area) in the DLBCL and B-CLL TMAs, the threshold values for all the quantitative measures and for the different TMAs and Abs evaluated were adjusted accordingly. All measurements were made using equal threshold values.
Survival Analyses
The impact on the prognosis of BCL-6 expression measured with both BCL-6 MAbs was estimated by OS analysis. Survival curves were plotted using the Kaplan-Meier method, and statistical significance of associations between individual variables and OS was determined using the log-rank test.
Results
The anti-BCL-6 GI191E/A8 MAb was initially validated by comparing WB data and IHC on cytocentrifuge preparations of human cell lines (Figure 1). Tissue lysates of tonsil and thymus, Burkitt's lymphoma cell lines (Ramos, AKATA, and Mutu-1), and DLBCL cell lines (KARPAS-422 and DB) reveal a clear 95-kDa band of BCL-6 protein. BCL-6 protein was not detected by WB or IHC studies in the Toledo (DLBCL) and REH (acute lymphoblastic leukemia) cell lines.
BCL-6 protein expression was then studied by IHC on paraffin-embedded tissues with the GI191E/A8 MAb and compared with other anti-BCL-6 MAbs produced by conventional means (ST42B/H7 and PG-6Bp). Positive staining of nuclei was seen in normal GC B-cells with all three MAbs. Although the specificity was similar for the three MAbs, the intensity of staining was significantly increased using the new GI191E/A8 MAb, as shown in Figure 2C.
The results of the comparison between the pressure-cooking methods of antigen retrieval using five different tonsil samples, buffers, and/or enzyme digestion with proteinase K for the anti-BCL-6 MAbs dilutions are shown in Table 1. Optimal results were obtained using citrate buffer (pH 6.5) followed by 10-min proteinase K enzyme digestion for all anti-BCL-6 MAbs tested. We found that the use of proteinase K as an additional step in the antigen retrieval method helped to decrease background staining.
The effects of formalin and B5 fixation on BCL-6 immunostaining are shown in Table 2 and Figure 2. Because the results achieved comparing the BCL-6 clones PG-B6p and ST42B/H7 were extremely similar, only the results of clone ST42B/H7 are shown in Figure 2, Figure 3, and Figure 5.
For all three MAbs tested, the best results were achieved after 24 hr of formalin fixation (Table 2 and Figures 2C-2D), the standard method used in most laboratories (Werner et al. 2000).
In the case of ST42B/H7, under- and overfixation (12 and 48 hr, respectively) drastically decreased the BCL-6 signal, culminating in almost complete loss of signal after 1 week of fixation (Figures 2B, 2F, 2H). The B5-fixed sample showed weak BCL-6 staining (Figure 2J). In the case of GI191E/A8, under- and overfixation or B5 fixation only slightly decreased the staining, indicating that its specificity is less dependent on formalin-fixation times and type of fixative used (Figures 2A, 2E, 2G, 2I).
Antibody-affinity assay by ELISA

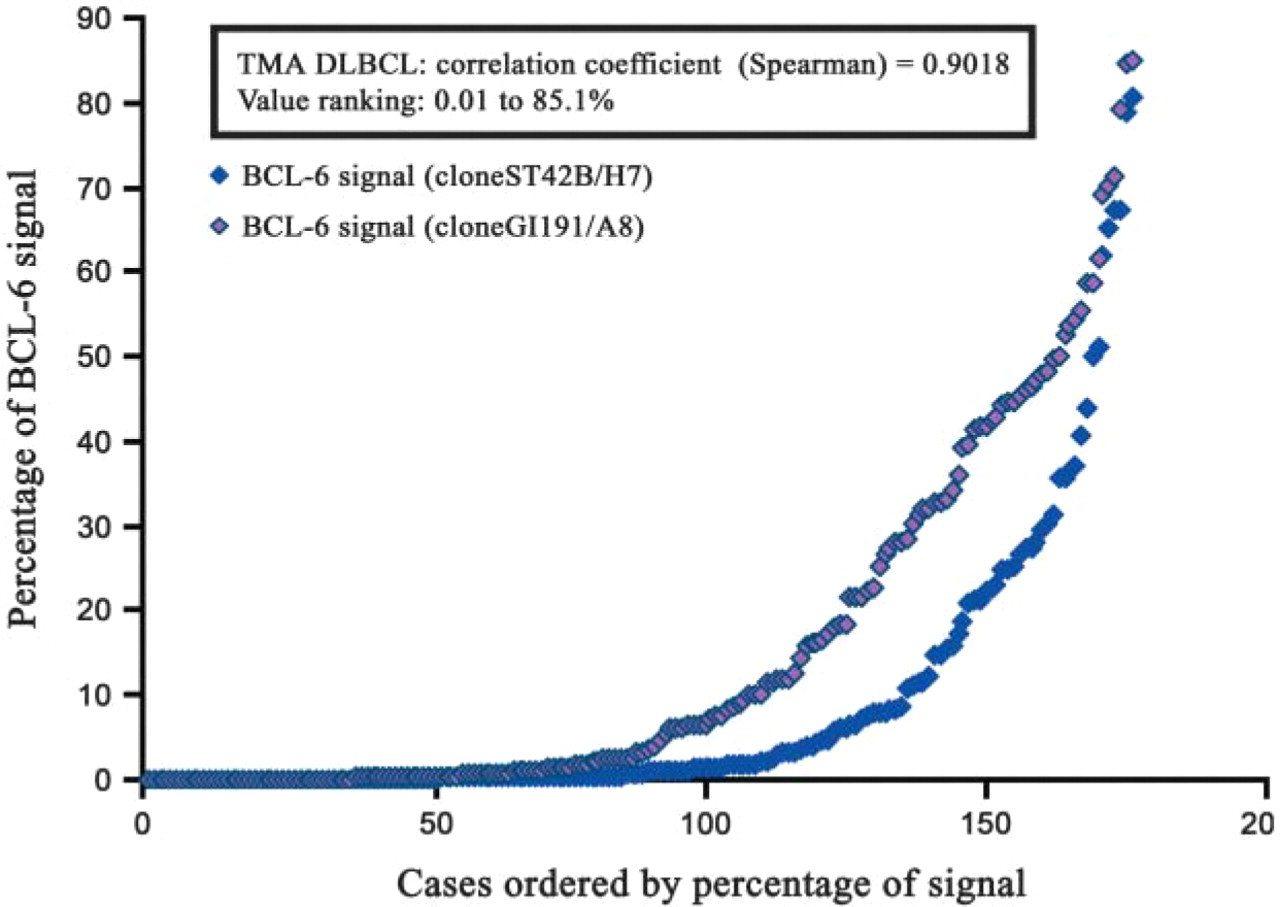
Comparative analysis using BLISS analysis and quantification of BCL-6 staining with GI191E/A8 and ST42B/H7 MAbs on a TMA of DLBCL cases, ordered by percentage of area with positive signal. BCL-6 values ranged from 0.01 to 85.1%, with a close correlation between the two MAbs (Spearman correlation coefficient = 0.9018). Staining intensity was very similar in those cases where BCL-6 expression was highly overexpressed or absent. BCL-6 detection with GI191E/A8 MAb was more sensitive in cases with intermediate BCL-6 expression.
To evaluate the affinity of our MAbs, we performed an Ab-affinity assay using an ELISA assay. The affinities of the MAbs were estimated by reacting them with BCL-6 protein captured on the solid phase followed by OD measurement at 405 nm. Clone GI191E/A8 gave an OD 405-nm reading of 1.0 at 6.25 nM, whereas ST42B/H7 yielded a value of 1.0 at 100 nM (Table 3). Thus, the affinity of clone GI191E/A8 was ∼16 times that of clone ST42B/H7.
The study of anti-BCL-6 MAbs immunostaining on tumor samples was performed using TMA analysis for DLBCL and B-CLL and analyzed using the BLISS system. As expected, all B-CLL cases staining with both MAbs were BCL-6 negative or displayed only faint nuclear expression in scattered cells (BCL-6 values ranging from 0 to 0.88%), showing that the staining was specific for BCL-6 protein and that similar epitopes were not recognized (data not shown).
The comparative study of BCL-6 expression in TMA analysis of DLBCL cases (Figure 3) revealed a close correlation between the ST42B/H7 and GI191E/A8 MAbs (Spearman correlation coefficient = 0.90). The majority of samples expressed a strong nuclear signal in most tumoral cells (values from 0.01 to 85.1%, Figure 3). Although the detection of BCL-6 protein was identical in those cases where BCL-6 was highly overexpressed or absent, in the cases where BCL-6 expression was intermediate, its detection with GI191E/A8 MAb was more sensitive (Figure 3). Moreover, the average percentage of BCL-6 signal significantly increased when GI191E/A8 MAb was used (15.11 for GI191E/A8 vs 8.5 for ST42B/H7).
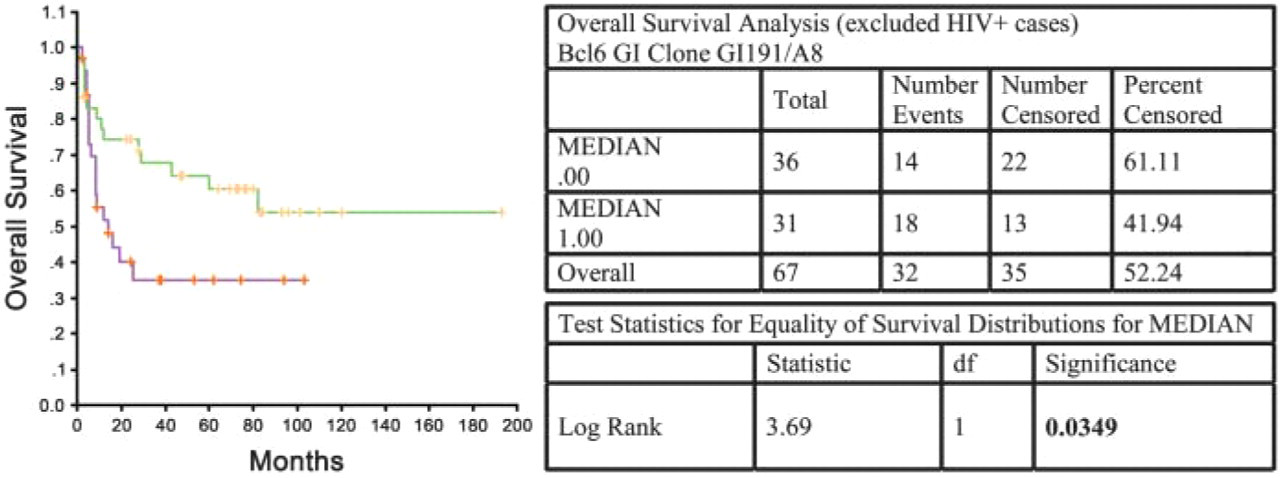
Survival analysis (overall survival) in cases of DLBCL (excluding HIV + cases) using the GI191E/A8 MAb. Log-rank test: p = 0.0349.
Figure 4 and Figure 5 illustrate the application of BCL-6 protein expression (using MAb clones ST42B/H7 and GI191E/A8) as an independent predictor of OS in DLBCL. Significant association of higher BCL-6 expression (as measured using GI191E/A8, Figure 4) and longer median OS times in DLBCL (log-rank test: p = 0.0349) was found, as previously described by other groups (Lossos et al. 2001). Using ST42B/H7 MAb in the same patient group (Figure 5), there was also a direct relationship between BCL-6 expression and survival, although the association was not significant (log-rank test: p = 0.0548). These data support the greater sensitivity and specificity of the MAb produced by genetic immunization.
Discussion
MAbs and IHC techniques on formalin-fixed, paraffin-embedded sections have become indispensable diagnostic and prognostic tools in the management of lymphoproliferative neoplasms. GI has been used mainly to obtain MAbs, which recognizes the native form of the protein but, as described here, GI can also be used as an effective alternative method for the production of MAbs for IHC on formalin-fixed, paraffin-embedded sections. This technique is much faster and less expensive, compared with conventionally produced MAbs. Normally, protein production and purification take from 1 to 4 months and pose significant difficulties in some cases. GI does not require protein for immunization or for the screening procedure.
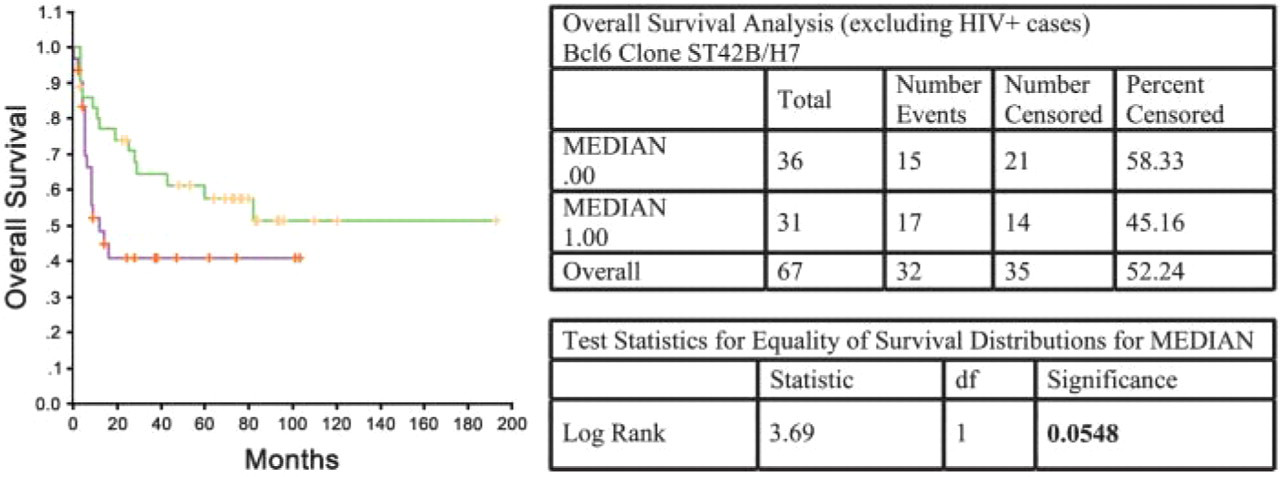
Survival analysis (overall survival) in cases of DLBCL (excluding HIV + cases) using the ST42B/H7 MAb. Log-rank test: p = 0.0548.
We demonstrate that GI191E/A8 presents superior sensitivity, and relative independence from factors such as fixation procedures allows better detection of BCL-6 protein in paraffin-embedded tissue. Low MAb sensitivity associated with low protein expression levels and incorrect tissue fixation procedures are the elements most commonly associated with pitfalls in routine IHC diagnosis. However, more studies with other antigens are necessary to demonstrate that GI is better suited for the production of MAbs for the detection of formalin-fixed, paraffin-embedded antigens.
MAbs are invaluable tools that have helped to extend our knowledge of biological systems for over two decades. The development of alternative methods for production of MAbs such as GI is likely to be important for meeting the increasing demand for MAbs for diagnosis, drug development, research, and therapy. GI is an important platform for the production of MAbs, allowing accurate and sensitive detection and screening of proteins in paraffin-embedded tissue samples and providing a link between genomic- and proteomic-derived data.
Footnotes
Acknowledgements
This work was supported by grants from the Ministerio de Ciencia y Tecnología (BIO2000-0275-C02/01-/02, SAF2001-0060), Comunidad Autónoma de Madrid (CAM 08.1/0011/2001.1), Spain. José-Francisco Garcia is the recipient of a grant from the Ministerio de Ciencia y Tecnología (FP-2001-0977), Spain.
We would like to thank Jackie Cordell, Nuffield Department of Clinical Laboratory Sciences, Oxford, for providing us with the NS-1 myeloma cell line (P3/NS1/1-Ag4-1). We also extend our appreciation to the staff of the CNIO Tumour Bank for their efficient provision of tumor simples and to Juan Fernando Martínez Leal, Carmen Blanco Aparicio, and Jesús M. Fominaya Gutiérrez of the CNIO Experimental Therapies group for advice. Further thanks are due to all members of the CNIO Animal Facility and Immunohistochemistry Unit for their technical support.