
Abstract
In vivo-applied sodium selenide or sodium selenite causes the appearance of zinc-selenium nanocrystals in places where free or loosely bound zinc ions are present. These nanocrystals can in turn be silver enhanced by autometallographic (AMG) development. The selenium method was introduced in 1982 as a tool for zinc-ion tracing, e.g., in vesicular compartments such as synaptic vesicles of zinc-enriched (ZEN) terminals in the central nervous system, and for visualization of zinc ions in ZEN secretory vesicles of, e.g., somatotrophic cells in the pituitary, zymogene granules in pancreatic acinar cells, beta-cells of the islets of Langerhans, Paneth cells of the crypts of Lieberkühn, secretory cells of the tubuloacinar glands of prostate, epithelium of parts of ductus epididymidis, and osteoblasts. If sodium selenide/selenite is injected into brain, spinal cord, spinal nerves containing sympathetic axons, or intraperitoneally, retrograde axonal transport of zinc-selenium nanocrystals takes place in ZEN neurons, resulting in accumulation of zinc-selenium nanocrystals in lysosomes of the neuronal somata. The technique is, therefore, also a highly specific tool for tracing ZEN pathways. The present review includes an update of the 1982 paper and presents evidence that only zinc ions are traced with the AMG selenium techniques if the protocols are followed to the letter.
T
In addition to zinc ions found in ZEN neurons, zinc ions are known to exist in secretory vesicles of a number of exo- and endocrine glands, including testis, epididymis, prostate, pancreas, male mouse salivary glands, pituitary, and intestine, and extracellularly in uncalcified bone and semen (Danscher 1982; Danscher et al. 1985a,1999; Thorlacius-Ussing et al. 1985; Frederickson et al. 1987a; Stoltenberg et al. 1996,1997; Sørensen et al. 1997,1998; Kristiansen et al. 2001).
The NeoTimm method was introduced in 1981. This zinc ion-specific version of Timm's original sulfide silver method (Timm 1958) evolved from two Timm modifications (Haug 1974; Danscher and Zimmer 1978). Timm's technique involves the creation of zinc-sulfur nanocrystals, and the NeoTimm method includes transcardial perfusion or intravenous (IV) injections in deeply anesthetized animals with a sodium sulfide solution or immersion of tissue blocks, slides, or sections in a solution of glutaraldehyde and sodium sulfide (Danscher 1981; Danscher et al. 2004). To ensure that the NeoTimm method demonstrates only zinc ions, the amount of sodium sulfide molecules has to match the amount of zinc ions in the tissues. The right amount has been defined as the highest level of sulfide ions that does not cause staining of sections from an animal exposed to 1000 mg/kg body weight of the zinc chelator diethyldithiocarbamate (DEDTC) 1 hr before perfusion (Danscher et al. 1973; Danscher 1981). In vivo chelation with the red chelator dithizone likewise results in a lack of Timm staining (Danscher et al. 1985b). Tissue sections from animals that have been perfused with sodium sulfide according to the NeoTimm method or treated in vivo with sodium selenide/selenite cannot be stained with the fluorescence techniques TSQ and Zinquin, which proves that it is the same pool of zinc that is stained with the different zinc-specific methods (Danscher 1981,1982; Danscher et al. 1985b; Frederickson et al. 1987b; Zalewski et al. 1993).
The selenium technique involves, as mentioned above, in vivo injections of sodium selenite/selenide and is presently the only available way of capturing zinc ions in vivo for subsequent tracing of zinc-selenium nano-crystals in tissue sections by autometallographic (AMG) silver enhancement. Another valuable quality of the technique is that zinc-selenium nanocrystals are more resistant to weak acids than are the zinc-sulfur nano-crystals, the silver-enhanceable entities of the NeoTimm methods (Danscher 1981,1996; Stoltenberg and Danscher 2000; Danscher et al. 2004). This relative pH stability of the zinc-selenium nanocrystals is also the quality that makes it possible to trace the neuronal somata of ZEN neurons. Nanocrystals of zinc-selenium created in the ZEN terminals and/or in the ZEN axons after, e.g., a local injection with sodium selenide, accumulate in lysosomes of the ZEN neuronal somata after having been translocated by retrograde axonal transport (Danscher 1982; Howell and Frederickson 1990; Christensen et al. 1992).
We did not find AMG grains in the lysosomes of animals injected intracerebrally (IC) with sodium sulfide, but only in a pool of synaptic vesicles in the ZEN terminals and their synaptic clefts. The AMG silver grains resulting from such IC injections are released into the synaptic clefts over the following 24-48 hr (Danscher 1984b; Pérez-Clausell and Danscher 1986). We have hypothesized, therefore, that due to the low pH in the lysosomes, retrogradely transported zinc-sulfur nanocrystals are dissolved while zinc-selenium nanocrystals are left intact for a period of time (Danscher 1994). Thus, the zinc-selenium autometallography (ZnSeAMG) technique for tracing ZEN neuronal pathways involves systemic or intraperitoneal (IP) injections of sodium selenite/selenide into brain and spinal cord and survival times that allow the zinc nanocrystals to be transported along the ZEN axons and deposited in lysosomes in the ZEN neuronal somata (Danscher 1982,1984b,1994; Howell and Frederickson 1990; Slomianka et al. 1990; Christensen et al. 1992; Wang et al. 2001a; Brown and Dyck 2003).
Selenium is the third member of the sixth group of the periodic system, next to sulfur. The chemistry of these two elements is in many respects quite similar, but they also differ considerably due to differences in ionic volume and electron negativity.
Sodium selenite was the first selenium compound used to create zinc-selenium nanocrystals in living organisms (Danscher 1982). It is a rather toxic substance, and experimental animals can show signs of lung edema after IP injections and demonstrate considerable lesions after IC injections. Therefore, sodium selenite exposure should always be performed on anesthetized animals and be followed by pain-killing drugs like buprenorphin when longer survival times are needed. The best possible selenide ion donator for IC injections has been found to be sodium selenide (Christensen et al. 1992). The synthesis of this drug is laborious in a standard laboratory but, fortunately, sodium selenide is now commercially available (Alfa 36187; Zeppelinstrasse 7, Karlsruhe, Germany).
The aim of the present study was to present updated, optimal, and easy-to-apply protocols for light-and electron-microscopy ZnSeAMG. The possibility of using the technique in mapping the biological and toxicological aspects of selenium compounds is mentioned in the Discussion.
Materials, Methods, and Results
It is recommended that animals undergoing ZnSeAMG be anesthetized with ketaminol and narcoxyl and that Temgesic (buprenorphin) be used as an analgeticum when the selenium-exposed animals must survive the initial anesthesia.
Tracing of ZEN Terminals by IP Injections of Sodium Selenite
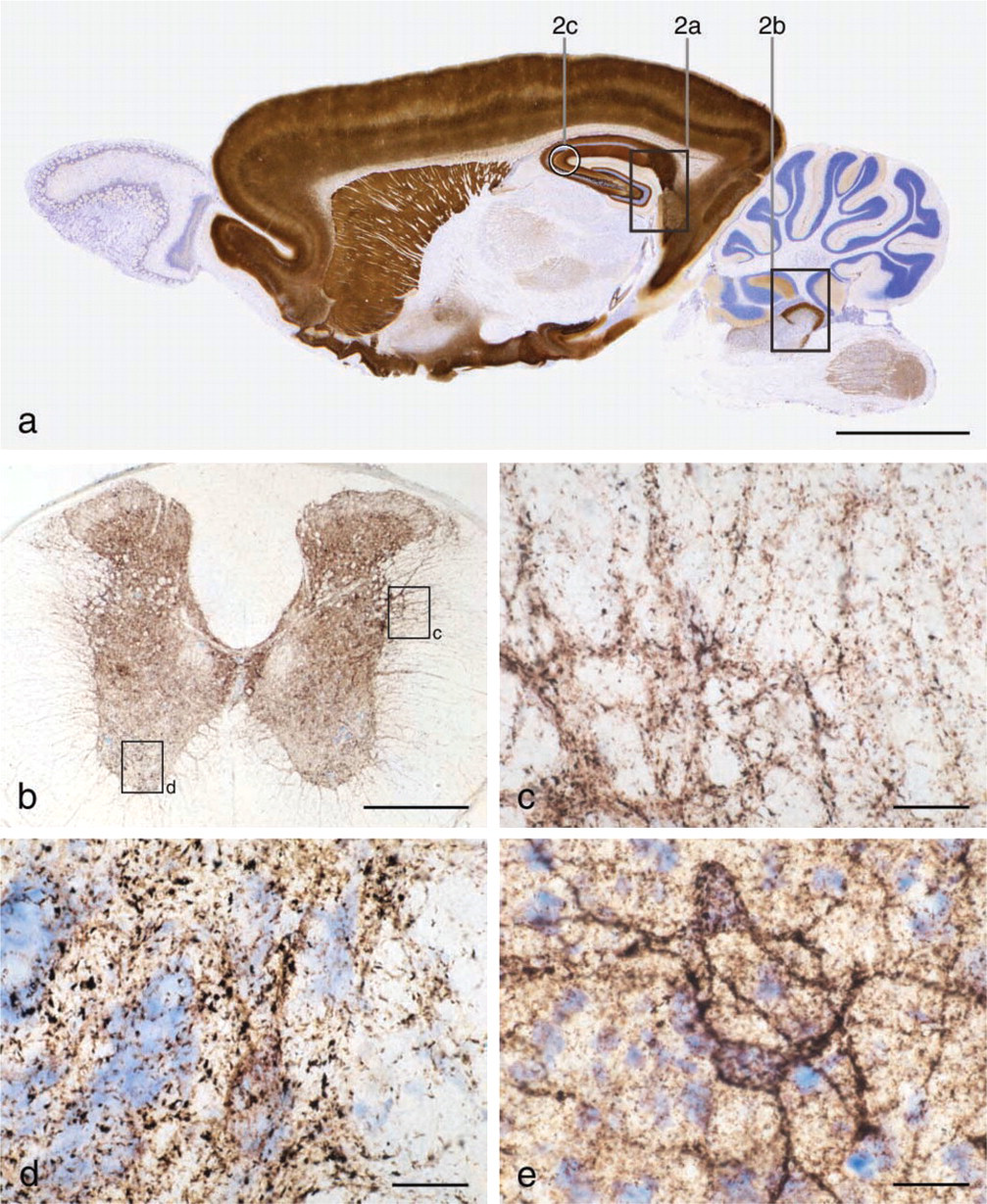
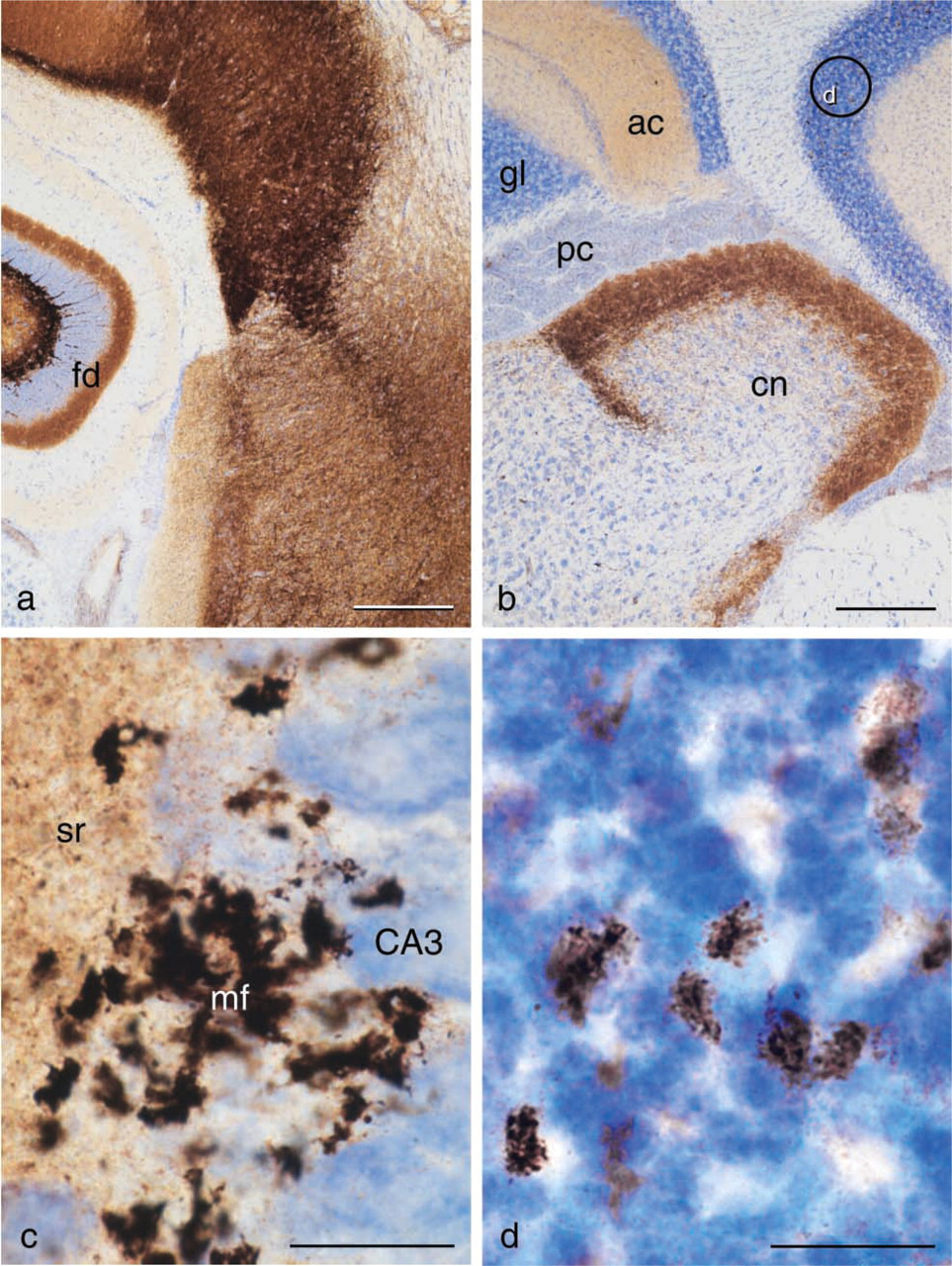
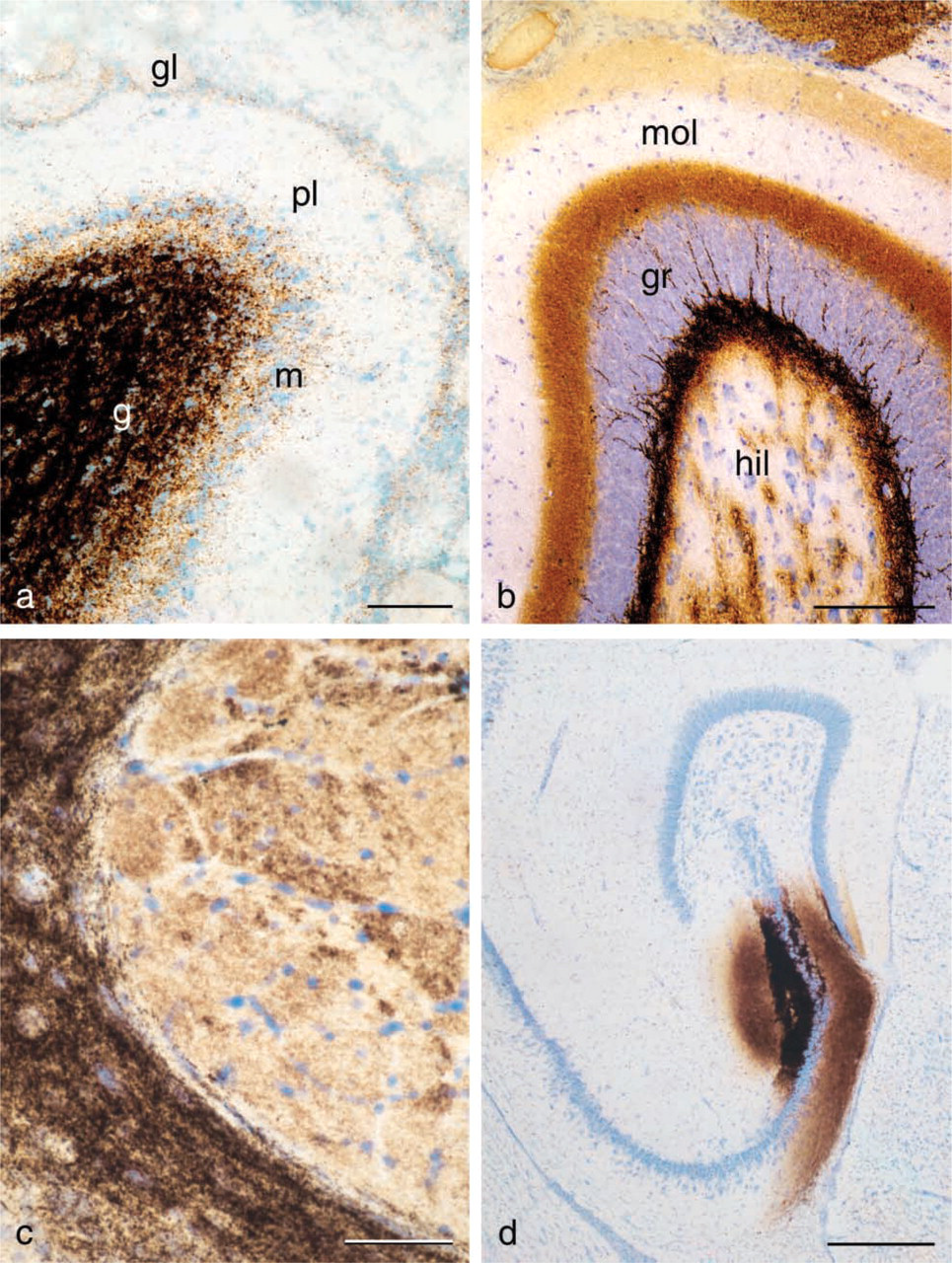
Tissue sections containing ZEN cells from animals given selenium under anesthesia and sacrificed after 1-2 hr demonstrate an AMG staining of zinc-selenium nano-crystals located in different types of ZEN vesicles (Figures 1 and 2, Figures 3a and 3c, Figure 4c).
Sodium selenite is a good choice for systemic creation of zinc-selenium nanocrystals. However, in some cases an IP or IV injection of sodium selenide might prove a better solution.
(
Tracing of ZEN Pathways by IC Injections of Sodium Selenide
Tracing of ZEN neuronal somata takes place by IP or IC injections of sodium selenide. The IC injections can be performed either by electrophoreses or by pressure injection of selenide ions. If the IP-injected selenide is used for tracing ZEN neuronal somata in the brain, it is important that the dose of sodium selenide used is high enough to ensure that all ZEN somata are loaded (Slomianka et al. 1990). The IP technique has, e.g., proven to be unsuccessful for tracing ZEN somata in the spinal cord, most likely because a sufficient level of selenium-loaded ZEN terminals cannot be achieved.
After 24 hr, some ZEN terminal staining will continue to be visible in the brain, in particular if the IC technique is used. This staining results from synaptic vesicles that have not yet released their content of zinc-selenium nanocrystals into the synaptic cleft or have been transported to the ZEN somata by retrograde axonal transport (Figure 4d) and can be seen up to 48 hr after injection. Nevertheless, the retrogradely stained ZEN somata are distinct (Figures 4a and 4b). Terminal staining can be reduced by pre-incubation with H2O2. Although this technique, of course, reduces the overall staining, it might be useful for preparing clear low-magnification photographs of the ZEN somata (Brown and Dyck 2003).
Tracing of Zinc Ion Pools in Exo- and Endocrine Secretory Cells
The finding of zinc-selenium nanocrystals in secretory vesicles of secretory cells of both the exocrine and the endocrine pancreas, as well as mast cells (Figure 4e), liver (Figure 4f), intestine, and many more locations, indicates that vesicular zinc ions, transported into the vesicular vacuum by specialized transmembrane transporting molecules (e.g., ZnT3), must have a more general function in the secretory and synaptic vesicles. It is also characteristic that it is always only a fraction of the vesicles in a certain secretory cell or neuronal terminal that contain zinc ions. Whether this reveals a difference in maturity of the vesicles or reflects the presence of more than one type of secretory/synaptic vesicle is not known at present. It might be reasonable to term cells with zinc ion-containing vesicles Zinc ENriched (ZEN) cells, inasmuch as they may have a basic biological mechanism in common.
Optimal and Easy-to-apply Protocols for ZnSeAMG
ZEN Terminal Tracing by In Vivo Exposure to Sodium Selenite
Animals are anesthetized with ketaminol and narcoxyl and injected with 20 mg sodium selenite per kg body-weight. Temgesic (buprenorphin) is used as an analgeticum. After 1 1/2 to 2 hr, the animals are either decapitated, the brains taken out, placed on a freeze stand, and cut in a cryostat, or the animals are transcardially perfused with buffered glutaraldehyde for 10 min and the brains placed in the fixative for 2 hr. The brains are then processed for either cryostat, paraffin, or vibratome sectioning.
(
ZEN Terminal Tracing by In Vivo Exposure to Sodium Selenide
All animal procedures are the same as above for sodium selenite, except for IP injections of 15 mg sodium selenide per kg bodyweight.
Tracing of ZEN Pathways by Local or Systemic Injections of Sodium Selenide
Before injection, the animals are deeply anesthetized with ketaminol and narcoxyl. Temgesic is used as an analgeticum. The animals are then placed in a stereotaxic apparatus, the calvarium is freed, and holes are drilled with a dentist's drill. Injections of 1 μl of a 10 mg/ml freshly made sodium selenide are performed with a Hamilton syringe (injection volume can vary between, e.g., 0.02 and 10 μl) over a period of 10 min. The syringe is left in situ for 2 min after the injection.
Alternatively, the selenium is applied iontophoretically with a micropipette. An intermittent current of 4 μA (tip negative) is applied for a total of 20-30 sec over a period of 5 min. The micropipette is left in situ for an additional 5 min after the injection (Christensen et al. 1992). After 1- to 48-hr survival, depending on the length of the ZEN pathways and type of animal used, the animals are re-anesthetized and either decapitated or perfused transcardially with a fixative as described above.
Brains from the decapitated animals are frozen with CO2. Brains from the perfused animals are kept in the fixative for at least 1 hr and then either vibratomized to be processed for Epon embedding, placed in sucrose, and cut on a cryostat or embedded in paraffin, vide supra.
Tracing of Zinc Ions in Non-neuronal Tissues (e.g., Testis, Salivary Gland, Pancreas, Prostate, Epididymis, Small Intestine, Liver, Lung, Adrenal, Bone, Thyroid Gland, and Anterior Pituitary)
Sodium selenite/sodium selenide is injected IP into anesthetized animals. After 1 1/2- to 2-hr periods of survival, the animals are killed by transcardial perfusion with 3% glutaraldehyde in a 0.1 M phosphate buffer for 10 min. The organs/tissues of interest are removed and processed as described for the brain. If the organs are small or lack rigidity, it is advantageous to stabilize them by dripping agar onto the vibratome stage. A 5% agar solution is boiled for 10 min, and when the solution has cooled to ~40C, the solution is dripped onto the organ pieces, where it will solidify. Bones are sawed into 300-μm-thick sections that are dipped in a 0.5% gelatin solution and AMG developed. Small pieces of the developed bone sections are fixed in osmium and embedded in metacrylate or Epon. The vibratome sections are developed as described above.
It is possible to trace extracellular pools of zinc ions, e.g., in unmineralized bone matrix (Danscher et al. 1999), semen plasma, secretions in the prostate secretory system, ductus epididymidis, and crypts of Lieberkühn. Zinc ions in such locations are found by exposing the organisms to sodium selenite or sodium selenide in vivo as described above.
Zinc Specificity of the ZnSeAMG Technique
IP injections of 150 mg dithizone per kg body-weight 4 hr before sodium selenide exposure result in a complete block of the ZnSeAMG staining in brain sections (see also Danscher 1982,1984a-c).
IP injections of 1000 mg DEDTC 1 hr before selenium treatment result in unstained brain sections (see also Danscher et al. 1973; Danscher 1982,1984a).
Local injections of DEDTC in rat brain followed by IP injections of sodium selenite after the normal ZnSeAMG procedure result in a spherical, totally unstained “hole” in the staining pattern, revealing where the zinc ions have chemically bound to the chelating DEDTC molecules and thereby been made inaccessible to the selenide ions (Danscher 1984b).
It was not possible to stain brain sections from animals treated with sodium selenite IP 1 1/2 hr before sacrifice with either of the fluorescence stains TSQ or Zinquin.
Brains from ZnT3 mice show no ZnSeAMG staining of ZEN terminals in mouse brain (Stoltenberg et al. unpublished data).
The ZnSeAMG staining patterns of mouse brain, spinal cord, and olfactory bulb are identical to the ZnT3 immunohistochemical staining patterns of the same localities (e.g., Jo et al. 2000b).
Synaptic ZEN vesicles of both glutaminergic and GABAergic ZEN terminals contain ZnT3 proteins in their vesicular membrane.
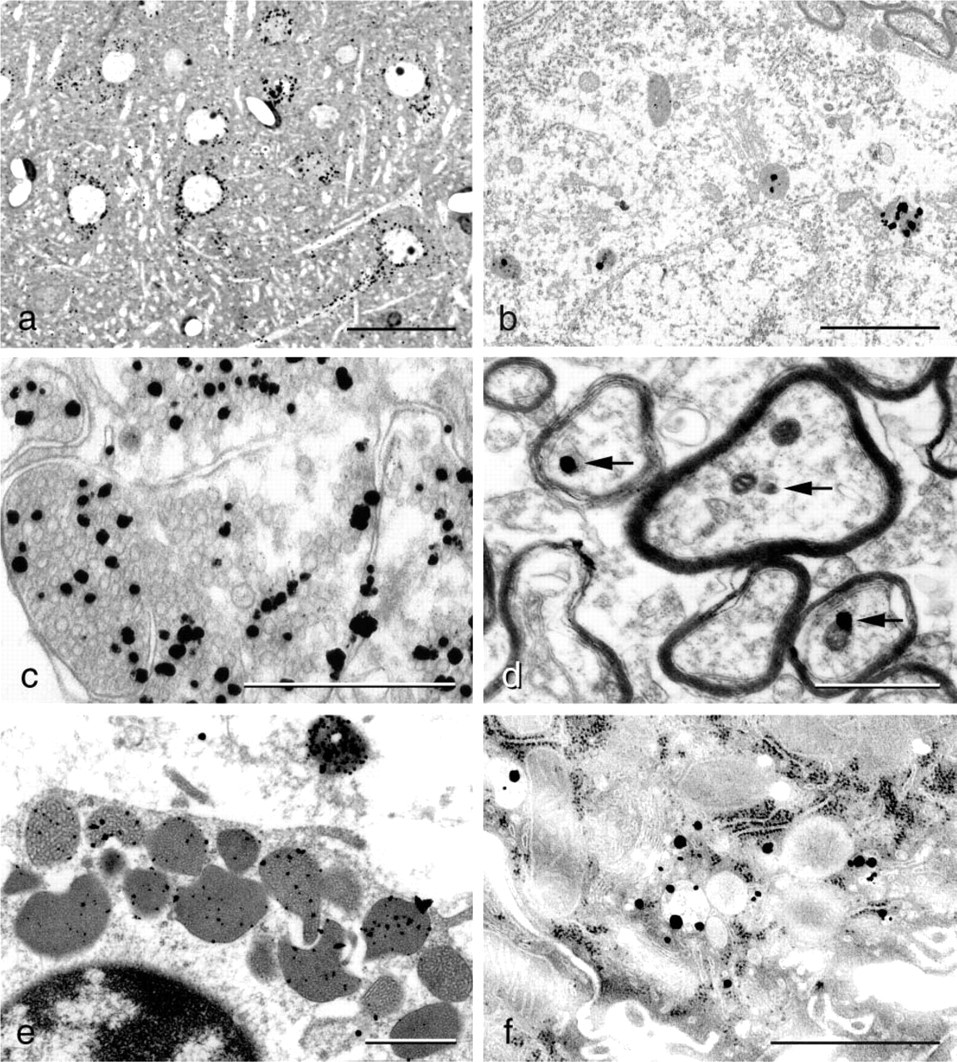
Multi-element (PIXE) analysis of isolated ZnSeAMG grains shows the presence of silver and selenium, while the same analysis on isolated ZnSAMG grains shows that these grains contain silver and sulfur. These results reveal that the zinc-selenium and zinc-sulfur nanocrystals are transformed into silver-selenium and silver-sulfur nanocrystals, respectively, in the initial phases of the autometallographic development (see also Danscher 1984a,1996).
(
(
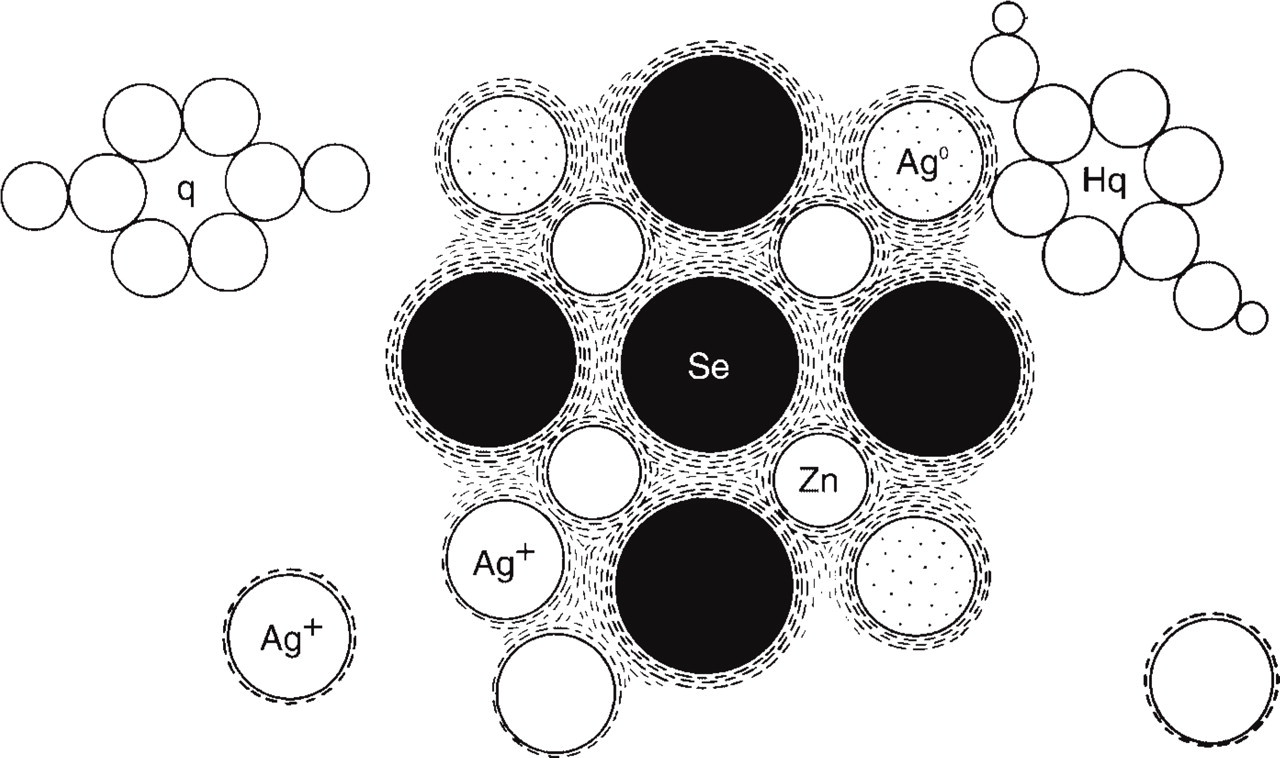
Camera lucida drawing of the autometallographic process. Electrons released from hydroquinone molecules adhering to the zinc-selenium nanocrystals build up energy in the valence cloud that forms the attraction force of the nanocrystal. The higher energy level increases the statistical probability of silver ions that have connected to the nanocrystal to catch an electron and become a silver atom. The “new” silver atom shares valence electrons with the crystal valence cloud of the original nanocrystal, i.e., has become a genuine part of the original zinc-selenium nanocrystal. As long as the AMG development proceeds, the nanocrystal will grow in size, i.e., be silver enhanced. Ag0, silver atom; Ag+, silver ions; Hq, hydroquinone; q, quinone; Se, selenium; Zn, zinc.
Future Fields of Application of the ZnSeAMG Techniques
AMG Tracing of Zinc Ions in Cell Suspensions and Cell Cultures (e.g., Sperm Cells, Macrophages, and Blood) and Organotypic Cell Cultures
On the basis of unpublished results, we suggest that the cells in question can be exposed to sodium selenide or sodium selenite either in vivo or in vitro. After having been collected, the samples are concentrated by centrifugation. The resulting pellet/concentrated probe is then fixed (e.g., with glutaraldehyde (GA) or paraformaldehyde (PFA) and embedded in Epon. Another approach involves AMG development of the pellet before embedding in Epon. The AMG development is stopped by adding the AMG stop bath (a 5% thiosulfate solution), after which the cells are rinsed by adding distilled water and centrifuged several times; finally, the cells are fixed with osmium tetroxide, rinsed again as described above, and embedded in Epon. Another possibility is to make smears on glass slides of the cell suspensions/cultures and develop them as described above. Organotypic cultures are exposed to sodium selenide/selenite by adding it to the medium. The tissue is rinsed in a 0.1 M Sørensen phosphate buffer and fixed for 10 min in buffered GA, then rinsed again in distilled water, and AMG developed for, e.g., 60 min. After a rinse, the tissue is fixed in osmium tetroxide for 30 min, re-rinsed in distilled water, and embedded in Epon.
It is important to stress that these new applications demand individual adjustments and must be checked for zinc specificity with DEDTC or other non-toxic zinc chelators.
Can Other Selenium Compounds Give Rise to ZnSeAMG Staining?
Since the introduction of the selenium method (Danscher 1982), different selenium compounds, including selenocysteine and selenomethionine (O'Toole et al. 1995), have been tested to find the ideal selenium donator for the demonstration of ZEN terminals as well as retrograde axonal transport of zinc ions (Danscher 1984b,1994; Howell and Frederickson 1990; Christensen et al. 1992). The major disadvantage of most of the compounds previously investigated has been their rather high tissue toxicity, causing large tissue necrosis and, in some cases, even death.
We have tested many commercially available selenium compounds (e.g., selenocysteine and selenomethionine, selenium tetrachloride, sodium selenate, powdered selenium, and dimethyl selenium). They all result in a less-complete binding of the AMG traceable zinc pool, but the ZnSeAMG technique might, nevertheless, be an interesting tool for research in selenium metabolism and toxicity.
Selenourea releases molecular selenium after gentle intracranial injections, with no tissue destruction along the injection canal (Figure 3d). Selenourea injected in amounts of 1 μl causes the appearance of a 2-mm spherical spot of AMG staining around the needle tip. Selenourea is an organic compound with selenium in the center of the molecule. The same properties that make selenourea suitable for intracranial injections exclude its use for IP or IV injections, inasmuch as the decomposition to molecular selenium will take place long before the selenide ions reach the brain.
Selenosulfate was synthesized in our laboratory, and it was found that 38 μmol selenosulfate resulted in a staining pattern and intensity similar to that of sodium selenite (Figure 3b). Selenosulfate-based ZnSeAMG is a stable and reproducible technique that results in uniformly stained sections of high technical quality. However, since selenosulfate is not commercially available and demands some effort to produce, we recommend sodium selenide/selenite as the best choice.
AMG Development
A vast amount of different AMG developers have been suggested since Liesegang introduced physical development into histology (Liesegang 1911). Stierhof et al. (1991) carried out a test on the quality of some of the most commonly used AMG developers at the time for silver enhancement of colloidal gold particles. Any suitable AMG developer can be used to silver enhance the zinc-selenium nanocrystals, including the silver acetate AMG developer (Hacker et al. 1988). In our laboratory, we continue to obtain the best results with the silver lactate/hydroquinone/gum arabic mixture (Danscher 1981).
It is imperative that all glassware and tools used for AMG development be rinsed in a 10% Farmer solution (see above) and that high-purity, high-quality chemicals and redistilled water be used.
In daylight, the developer is poured into jars containing the slides. These are placed in a 26C shaking water bath in a dark box—without the dark box the developer will gradually turn brown because of the creation of tiny silver particles. The developing time for 20-μm cryosections is 60 min. Development is stopped by replacing the developer with 5% thiosulfate. After 10 min, the stop bath is removed, and the slides are rinsed in distilled water before being counterstained with, e.g., toluidine blue and coverslipped. One should always include one or more control glass slides among the experimental slides, i.e., glass slides with sections that give a well-known pattern of AMG staining, to ensure that the development has been optimal.
Discussion
Selenium seems to function in the body primarily as a catalyst and has been found to function as a prosthetic group of a variety of enzymes and to interact with tocopherol to protect membranes from oxidative damage and facilitate the union of oxygen and hydrogen (Frost and Lish 1975). Selenium is also known to protect against the toxic effects of: (a) heavy metals, such as lead, cadmium, arsenic, and mercury; and (b) some organic toxicants in man and other mammals (Wilber 1980). The major selenium species in cereals and vegetables are selenomethionine and selenocysteine (Barceloux 1999), and it is now clear that selenium is essential to health and normal birth (Castillo-Duran and Cassorla 1999; Maiorino et al. 1999; Rayman and Rayman 2002).
The toxicity of most selenium compounds is rather low, but is dependant on the chemical form (Barceloux 1999). The US National Toxicology Program lists selenium sulfide as an animal carcinogen, but there are no results indicating other selenium compounds to be carcinogens (Barceloux 1999). However, from the thousands of articles on toxicity, it is obvious that the levels of sodium selenide/selenite used to create the pattern of zinc-selenium nanocrystals are very toxic to the animals. Therefore, it is imperative that the animals be anesthetized before being treated with the two selenium compounds, and that pain-killers be given if the animals must survive for periods that exceed the effect of the initial anesthesia.
The IC injection of sodium selenide results in little or no damage to brain tissue, while sodium selenite gives rise to substantial damage around the injection canal (Christensen et al. 1992; Danscher 1994; Christensen 1995; Christensen and Frederickson 1998). The selenite-caused lesions most likely are redox damage caused by the Se4+ being reduced to Se2- ions.
From the data obtained in the present study and from experience using the ZnSeAMG techniques for more than 20 years, it can be concluded that the two best selenium donors for in vivo binding of zinc ions in zinc-selenium nanocrystals are sodium selenide and sodium selenite (see also Christensen et al. 1992). The reason for this is, without any doubt, that only these two selenium compounds give rise to the necessary levels of selenide ions (Se2-) to cause the creation of zinc-selenide molecules that in turn accumulate in zinc-selenium nanocrystals.
A strange phenomenon is seen after IP injections of sodium selenite if the time of exposure is less than 1 1/2 to 2 hr. In such animals, the hilus fasciae dentatae and amygdala are not at all or only lightly stained. The reason for this is not completely understood, but we have hypothesized that it is partly the result of the fact that selenium has to be reduced from Se4+ to Se2- and that the number of zinc ions is particularly high in these areas, but also that local peculiar chemical conditions may play a role. The phenomenon vanishes after 1 1/2 to 2 hr of selenium exposure and is never seen after IC injections (see also Howell et al. 1989).
The functional roles of the ZnSeAMG-traceable zinc ions in the different cells are far from completely resolved. In pancreatic β-cells it has been found that zinc ions in the secretory granules function as a kind of glue that makes the insulin molecules collect in hexameres (Derewenda et al. 1989; Dodson and Steiner 1998; Søndergaard et al. 2003). In this way, the peptide becomes osmotically “invisible.” It could very well be that such a function of zinc ions in ZEN vesicles is the original one, i.e., the synaptic vesicles are equipped with transmembrane zinc ion transporters (ZnTs), causing the interior zinc ion pressure to increase, which then leads to an ordered packing of proteins or peptides in the vesicle. When a ZEN vesicle connects to the surface membrane and the vesicular space confluences with the extracellular space, zinc ions will pour out, leading to a release of protein/peptide from the packed form.
The next step in this process could be that the released zinc ions become involved in other processes. In pancreas, it has been suggested that the zinc ions process a paracrine function by inhibiting glucagon release from α-cells (Kim et al. 2000; Ishihara et al. 2003), and in the brain, zinc ions released in glutaminergic synapses have been suggested to influence different post-synaptic receptor sites, such as NMDA and GABA receptors (Li et al. 2001; Hosie et al. 2003).
In conclusion, for zinc-ion tracing, we recommend in vivo treatment with sodium selenite or sodium selenide, depending on the purpose. Other selenium compounds tested all result in a less-sufficient creation of zinc-selenium nanocrystals at locations known to harbor loosely bound and free zinc ions. Nevertheless, the ZnSeAMG approach might be of interest for research on the metabolism and toxicity of selenium-containing compounds that give rise to in vivo creation of zinc-selenium nanocrystals.
As an additional note, other metal-selenium nanocrystals can be silver enhanced by AMG, i.e., silver-selenium, mercury-selenium, and bismuth-selenium nanocrystals created in organisms that have been exposed to the above heavy metals (see also Stoltenberg and Danscher 2000). No doubt, more AMG-traceable heavy metal-selenium nanocrystals will be introduced in the years to come, either because it is revealed that they are metabolic end products of exogenous metals or because they can be used, e.g., for refining a histochemical technique.
Footnotes
Acknowledgements
This study was supported by The Danish Medical Research Council, Aarhus University Research Foundation, Danish Medical Association Research Fund, Aase & Ejnar Danielsens Fond, and the Lundbeck, Leo, Beckett, Gangsted, and Novo Nordic Foundations.
The authors gratefully acknowledge the skillful technical assistance of Ms D. Jensen, Mr A. Meier, Ms H. Mikkelsen, Mr T.A. Nielsen, Ms M. Sand, and Ms K. Wiedemann. We thank Søren Juhl for synthesizing the selenosulfate and commenting on the manuscript.