
Abstract
α-Dystroglycan (DG) is a negatively charged membrane-associated glycoprotein that links the cytoskeleton to the extracellular matrix. Previously, we described that α-DG covers the whole podocyte cell membrane in the rat. However, our finding was challenged by the description of a strictly basolateral localization in human kidney biopsies, using a different antibody against α-DG. Therefore, we studied the exact localization of glomerular α-DG by using these two antibodies in both species. The studies were performed by using monoclonal antibodies (MoAbs) IIH6 and VIA4.1 in immunofluorescence, confocal microscopy, and immunoelectron microscopy on both rat and human kidney sections, as well as on cultured mouse podocytes. The apical localization of α-DG on podocytes was more dominant than the basolateral localization. The basolateral staining with MoAb VIA4.1 was more pronounced than that of MoAb IIH6. With both MoAbs, the staining in rat kidneys was more prominent, in comparison to human kidneys. We conclude that α-DG is expressed at both the basolateral and apical sides of the podocyte. This localization suggests that α-DG plays a dual role in the maintenance of the unique architecture of podocytes by its binding to the glomerular basement membrane, and in the maintenance of the integrity of the filtration slit, respectively.
D
The localization of α-DG on podocytes has been controversial. We showed, with monoclonal antibody (MoAb) IIH6 as probe, that α-DG covers both the apical and the basolateral cell membranes of rat podocytes. In contrast, Regele et al. (2000) demonstrated a strict basolateral localization on human podocytes by using a different technique and by probing with a different MoAb to α-DG, MoAb VIA4.1. Both mouse MoAbs are directed at carbohydrate epitopes on α-DG; the immunoglobulin subclass of IIH6 is IgM and that of VIA4.1 is IgG1 (Ohlendieck et al. 1991; Michele et al. 2002). Because α-DG was shown to link the actin cytoskeleton of muscle and nerve to the extracellular matrix, it has been postulated that α-DG is a major adhesion molecule for podocytes. To date, the function of α-DG localized apically has not been discussed.
DG is encoded by a single gene, DAG1, and cleaved into α- and β-DG by posttranslational processing (Ibraghimov-Beskrovnaya et al. 1992). α-DG is heavily glycosylated with many sialic acid-rich oligosaccharides, which accounts for its negative charge. The predicted molecular mass of the core protein of α-DG is ≃74 kDa, whereas the apparent molecular mass, as detected by SDS-PAGE, is ≃156 kDa in skeletal muscle, and ranges from 120 kDa in brain to ≃200 kDa in bovine kidney (Ervasti and Campbell 1991; Gee et al. 1993; Bowe et al. 1994; Yamada et al. 1994; Smalheiser and Kim 1995). Because no differences in primary protein structure have been described, these observed differences in molecular mass seem to be due to differences in glycosilation (Ibraghimov-Beskrovnaya et al. 1993). Using biophysical methods, a dumbbelllike structure was visualized, in which the C-terminal region of α-DG interacts with the N-terminal region of the transmembrane β-DG (Brancaccio et al. 1995; Sciandra et al. 2001). Within podocytes, the intracellular C-terminal tail of β-DG interacts with utrophin, an autosomal homolog of dystrophin, which finally interacts with the actin cytoskeleton (Raats et al. 2000; Ilsley et al. 2002). Extracellularly, α-DG binds to laminin G modules that are present in agrin, laminin, and perlecan, which are localized in the extracellular matrix (Gee et al. 1994; Sugiyama et al. 1994; Yamada et al. 1996; Gesemann et al. 1998; Hohenester et al. 1999; Timpl et al. 2000). α-DG has been shown to play an important role in basement membrane assembly. Mice with a targeted disruption of the DG gene (DAG1−/-) do not develop Reichert's membrane, at which stage further development of the embryo stops (Williamson et al. 1997). Blocking of the binding site of α-DG to laminin with antibody IIH6 in kidney organ culture pertubates development of epithelium (Durbeej et al. 1995). The expression of glomerular α-DG has been studied in some human and experimental glomerular diseases: It was decreased in minimal-change nephropathy and in experimental adriamycine nephropathy, but remained stable in focal segmental glomerulosclerosis and in experimental passive Heymann nephritis (Raats et al. 2000; Regele et al. 2000).
Podocytes have a unique architecture, in which pedicles spread from the cell body, covering the capillary loop by the formation of foot processes. These foot processes interdigitate and form filtration slits. This shape can be maintained by the negative charge of sialic acids, which has been shown to cover the apical membrane (Faraggiana et al. 1982; Charest and Roth 1985; Holthofer et al. 1988; Wagner and Roth 1988). Podocalyxin has been identified as a major sialic acidrich glycoprotein covering this apical membrane (Kerjaschki et al. 1984). Removal of sialic acid by sialidase induces a loss of filtration slits and foot process effacement (Andrews 1979; Gelberg et al. 1996). Neutralization of the polyanionic surface by perfusion of kidneys with polycations such as protamin sulfate or hexadimethrine has similar effects, which can be prevented by scavenging these polycations with heparin (Kerjaschki 1978, 1994; Bridges et al. 1991).
As outlined above, there is still discussion about the exact localization of α-DG on podocytes. In this study, we addressed this question by applying different techniques (immunofluorescence and confocal and electron microscopy) and by using the available MoAbs against α-DG, i.e., IIH6 and VIA4.1. We found that in situ α-DG is localized at both the apical and basolateral sides of rat and human podocytes, and we confirmed this finding in cultured mouse podocytes. These observations indicate that glomerular α-DG plays a dual role in the maintenance of the unique architecture of podocytes by its binding to the GBM, and in the maintenance of the integrity of the filtration slit, resulting from its negative charge by sialic acid residues.
Materials and Methods
Immunofluorescence and Confocal Microscopy
Kidneys were harvested from adolescent female Wistar rats under isoflurane anesthesia, and immediately snap frozen in liquid nitrogen and stored until use. Human kidney sections were derived from donor kidneys unsuitable for organ transplantation and were stored at -80C, or from specimens of the nontumorous part of the kidney obtained by (partial) nephrectomy from patients with renal cell carcinomas, snap frozen in liquid nitrogen. Two-micron-thick kidney sections were prepared and stored at -80C until use. These sections were first air-dried, and subsequently fixed in ice-cold acetone (Merck; Darmstadt, Germany) for 10 min, unless stated otherwise. Subsequently, the sections were air-dried and incubated for 60 min at room temperature (RT) with mouse MoAb VIA4.1 (mouse IgG1, 20 μg/ml), MoAb IIH6 (mouse IgM, 6 μg/ml) in immunofluorescence (IF) buffer [1% BSA (Sigma; St Louis, MO), 0.1% sodium azide (Merck) in phosphate-buffered saline (PBS)]. Both antibodies were raised against rabbit muscle membranes and directed against carbohydrate epitopes of α-DG (Ohlendieck et al. 1991; Michele et al. 2002). Agrin was stained with hamster MoAb MI-90 (diluted 1:400) (Raats et al. 1997, 1998). Primary antibody isotype controls were used in the exact same concentrations. Subsequently, sections were washed three times for 10 min in PBS and incubated for 45 min at RT with the appropriate 488 or 568 Alexa fluorochrome conjugates diluted 1:200 in IF buffer: goat anti-mouse IgG (Molecular Probes; Leiden, The Netherlands), or FITC-conjugated goat anti-Syrian hamster IgG (Jackson Immunoresearch Laboratories, Inc.; Westgrove, PA). Goat anti-mouse IgG1 Alexa 568 and goat anti-mouse IgM Alexa 488 were used as conjugates in double stainings for α-DG with VIA4.1 and IIH6, respectively. Normal rat serum (diluted 1:25) was added when probing rat kidney sections. After rinsing, sections were postfixed in 1% paraformaldehyde (Merck) in IF buffer and embedded in Vectashield (Vector Laboratories Inc.; Burlingame, Canada). The sections were analyzed by a Leica TCS NT confocal system (Leica Lasertechnik GmbH; Heidelberg, Germany).
Immunoelectron Microscopy
Immunoelectron microscopy (IEM) was performed according to the two methods described previously for α-DG, respectively a pre- and a post-embedding technique (Raats et al. 2000; Regele et al. 2000).
For the pre-embedding technique, kidneys were immersion-fixed in a mixture of 10 mM periodate (Merck), 75 mM lysine (Merck), and 2% paraformaldehyde (PLP), pH 6.2, for 3 hr. After rinsing several times in PBS, the kidneys were cryoprotected by immersion in 2.3 M sucrose for 1 hr, and subsequently frozen in liquid nitrogen. Twenty-micronthick sections were incubated with the MoAbs IIH6, VIA4.1, or isotype controls, in IF buffer for 18 hr at 4C and then washed three times for 30 min with PBS. After rinsing, the sections were incubated for 90 min with the appropriate peroxidase-conjugated secondary antibody rabbit anti-mouse IgM/IgG (Dako; Glostrup, Denmark), then incubated for 90 min in IF buffer. After rinsing in PBS for three 30-min periods, the sections were preincubated for 10 min in PBS containing 0.05% diaminobenzidine (DAB; Fluka/Sigma-Aldrich, Germany) in 50 mM Tris[hydroxymethyl]aminomethane (Sigma). Subsequently, the sections were stained for 10 min with the same medium containing 0.003% hydrogen peroxide (Merck). After washing three times in distilled water, the sections were postfixed for 30 min at room temperature in 1% OsO4 [Electronic Microscopy Sciences (EMS); Hatfield, PA], pH 7.4, dehydrated, and embedded in Epon 812 (Merck). Ultrathin sections were prepared on an ultratome (Leica; Reichert Ultracuts, Wien, Austria).
Additionally, a post-embedding technique was performed. Kidneys were immersion-fixed in PLP and embedded in Lowicryl HM20 resin (EMS). Ultrathin sections were incubated via droplet method with the MoAbs IIH6 or VIA4.1, or isotype controls for 1 hr in IF buffer at RT, and washed several times in PBS. Subsequently, sections were incubated with 10 nm gold-labeled goat anti-mouse IgM or IgG (Amersham Biosciences; Little Chalfont, UK) for 1 hr in IF buffer, washed several times in PBS, then washed three times in distilled water and contrasted with 4% aqueous uranyl acetate (EMS) for 15 min. All sections were examined by a JEOL 1200 EX2 electron microscope (JEOL; Tokyo, Japan).
Cell Culture
Conditionally immortalized mouse podocytes (MPC5, generously provided by Dr. Peter Mundel, Division of Nephrology, Albert Einstein College of Medicine, Bronx, NY), which harbor the gene encoding the temperature-sensitive SV40 large T antigen under control of a γ-interferon-inducible H2Kb promoter, were used. These cells have been shown to polarize during differentiation (Mundel et al. 1997). Podocytes were maintained in RPMI 1640 (Invitrogen; Breda, The Netherlands) supplemented with 10% FCS (Greiner; Alphen aan de Rijn, The Netherlands) and 100 U/ml penicillin-streptomycin (Invitrogen) at 97% humidity, in 5% CO2. Cells were propagated in polystyrene flasks (Greiner) at 33C, with 20 U/ml mouse γ-interferon (Sigma) added to the medium. Podocytes were allowed to differentiate by culturing at 37C, without additional mouse γ-interferon, on collagen A (Biochrom; Berlin, Germany), laminin EHS (Campro Scientific; Veenendaal, The Netherlands), or Matrigel (BD; Alphen aan de Rijn, The Netherlands) -coated glass coverslips for 3 weeks. Passages 10-17 were used in these experiments. For immunofluorescence and confocal microscopy, podocytes were fixed in PLP for 10 min at RT, and subsequently washed three times in PBS and blocked for 20 min in blocking solution [2% FCS, 2% BSA, 0.2% fish gelatin (Amersham) in PBS]. Indirect immunofluorescence and confocal microscopy were performed essentially as described above in blocking solution, without postfixation. Normal goat serum (diluted 1:10) was added when probing with the conjugates, and the nucleus was probed with topro (diluted 1:20; Molecular Probes).
Results
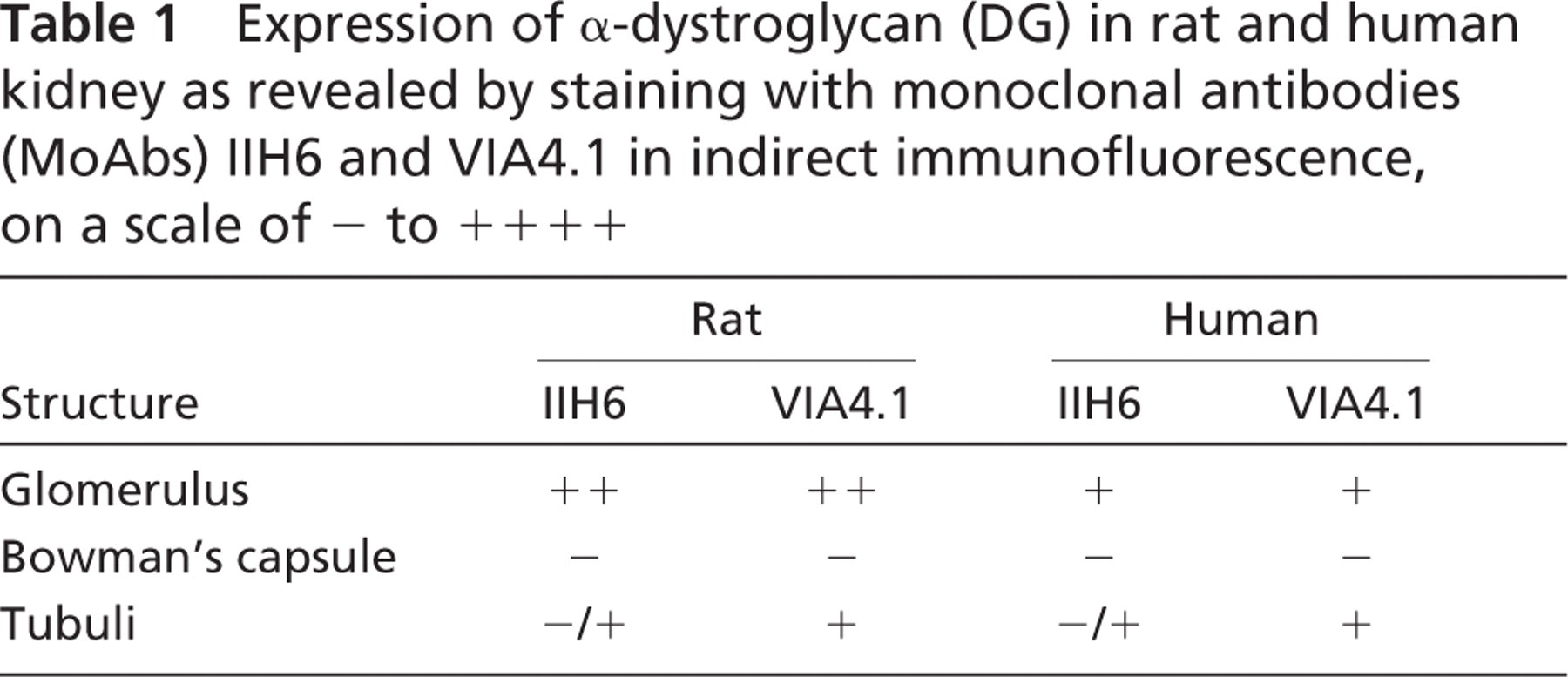
The indirect immunofluorescence of rat glomeruli with both MoAbs revealed a granular staining in a podocyte-like manner. In human glomeruli, however, this podocyte-like staining was less pronounced and a more fine granular aspect near the GBM was observed (Figure 1, left columns). The staining patterns and intensity of α-DG with MoAbs VIA4.1 and IIH6 on rat and human kidney sections were different (Table 1; Figure 1 and Figure 2, left columns). In general, the glomerular staining of α-DG with both antibodies was more pronounced in rat kidney than in human kidney. Bowman's capsule was never stained. For the tubuli, the staining patterns were similar for both antibodies in both species, but the staining intensity by both antibodies was different, showing a minor staining by MoAb IIH6 near the tubular basement membrane (TBM), in contrast to a marked staining by VIA4.1.
To investigate differences in the accessibility of carbohydrate epitopes on α-DG in rat and human kidney, we examined whether MoAbs VIA4.1 and IIH6 could inhibit the binding to their respective epitopes (Figure 1). MoAb VIA4.1 is a mouse IgG1, and MoAb IIH6 is a mouse IgM, which can inhibit the binding of α-DG to laminin G modules (Durbeej et al. 1995; Matsumura et al. 1997). The glomerular staining by IIH6 (green) was only slightly inhibited by preincubation with MoAb VIA4.1 (red), because the merge appeared predominantly yellow (colocalization; Figure 1, right columns). However, the accessibility of the glomerular epitope of MoAb VIA4.1 (red) could be blocked substantially by preincubation with MoAb IIH6 (green), because the merge appeared predominantly green. In the tubuli, the more pronounced staining by VIA4.1 near the TBM could not be blocked by preincubation with MoAb IIH6. Preincubation with isotype controls did not affect the glomerular or tubular staining by either antibody.
To investigate how the staining for glomerular α-DG was related to the GBM, we performed double stainings for α-DG and agrin on rat and human kidney sections using confocal microscopy (Figure 2). Generally, staining for α-DG was observed outside the GBM in a podocyte-like manner. Partial colocalization with agrin was observed, especially when stained by VIA4.1 (Figure 2, rat merge VIA4.1).
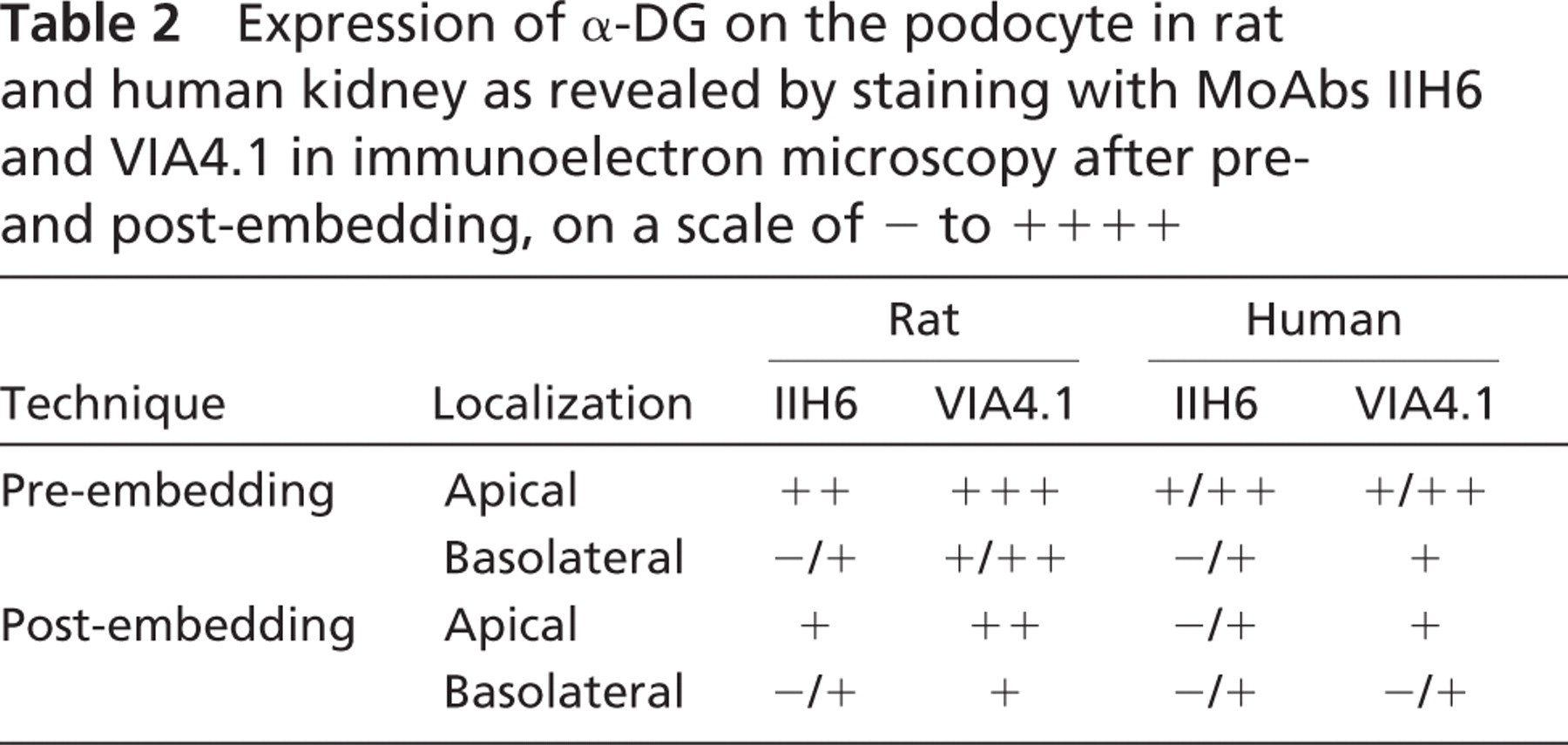
The analysis with immunofluorescence revealed a lesser basolateral staining of glomerular α-DG, suggesting that α-DG is predominantly expressed at the apical side of the podocyte. To determine the ultrastructural localization of α-DG on the podocyte, IEM was used, applying two different techniques. Preembedding incubation combined with DAB revealed a homogenous staining along the apical cell membrane and a weak staining at the basal side of the podocytes facing the GBM (Figures 3A, 3B, 3G, and 3I). The glomerular staining patterns of MoAbs IIH6 and VIA4.1 were in general identical, except that VIA4.1 showed a stronger staining at the basal side of the podocyte, particularly in human kidney sections (Figures 3I and 3J). The staining was consistently stronger in rat kidneys compared with human kidneys (Table 2). In addition, after Lowicryl embedding and staining with immunogold (post-embedding), the same pattern as in the pre-embedding technique was revealed, but with a much lower intensity (Figures 3C-3F, 3H, and 3J). In fact, this latter staining method seemed to be less efficient, in comparison to the pre-embedding technique.
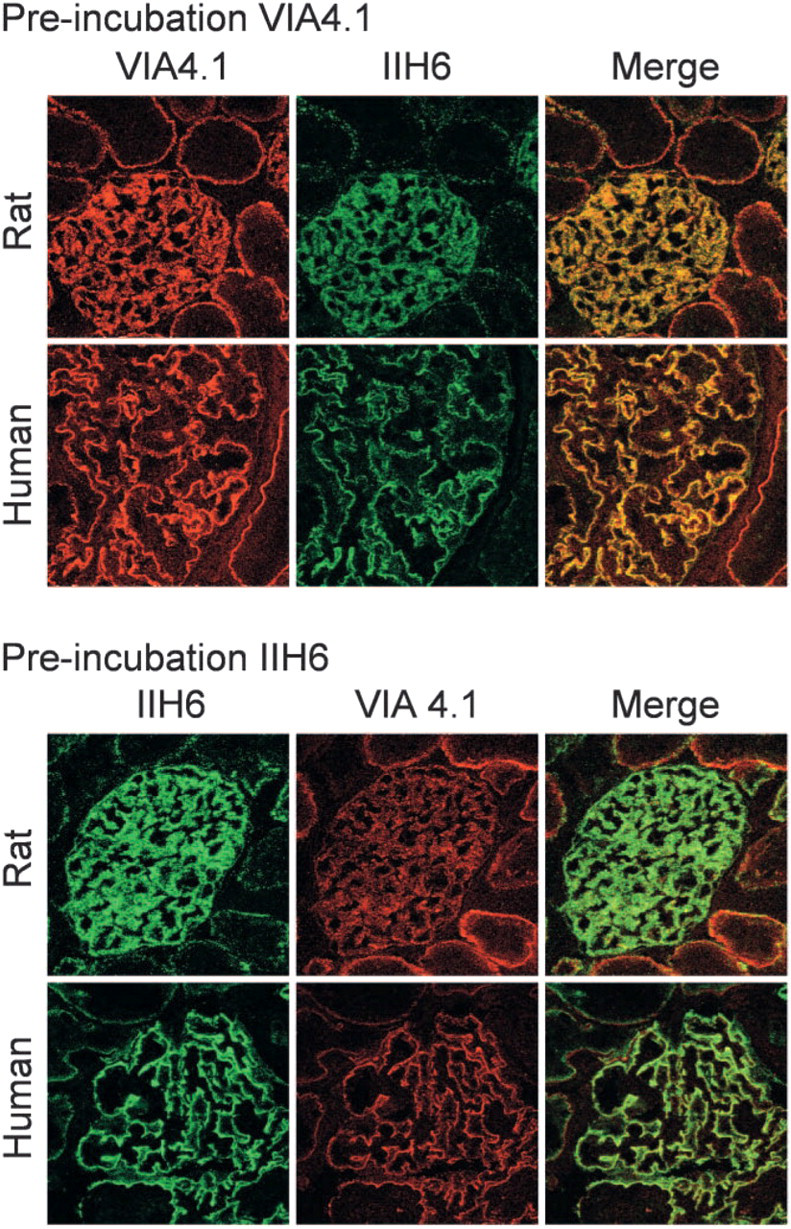
Cross-inhibition of staining of α-dystroglycan (DG) by monoclonal antibodies (MoAbs) VIA4.1 and IIH6. Confocal microscopy of rat kidney sections incubated first with MoAb VIA4.1 (red) and subsequently with MoAb IIH6 (green) or incubated first with IIH6 and subsequently with VIA4.1. The staining on rat kidney was stronger than that on human kidney. Within the glomerulus, preincubation with MoAb VIA4.1 (red) only slightly inhibited the staining of IIH6 (green), because the merge appeared predominantly yellow (colocalization). However, preincubation with MoAb IIH6 (green) substantially blocked the glomerular epitope of MoAb VIA4.1 (red), because the merge appeared predominantly green. Furthermore, the epitope of MoAb VIA4.1 was more abundantly expressed in the tubuli, compared with the epitope of MoAb IIH6 (magnification ×630). Isotype controls were negative (not shown).
Expression of α-dystroglycan (DG) in rat and human kidney as revealed by staining with monoclonal antibodies (MoAbs) IIH6 and VIA4.1 in indirect immunofluorescence, on a scale of - to + + + +
To further localize α-DG on the podocyte cell membrane, we investigated the expression on cultured mouse podocytes. Differentiated podocytes, maintained on coatings of collagen, laminin, or Matrigel, expressed α-DG on both their basolateral and their apical mem branes. Again, the staining by MoAb VIA4.1 (Figure 4, upper panel) was more pronounced at the basolateral membrane of the podocytes, compared with MoAb IIH6 (Figure 4, lower panel). The apical staining intensity of podocytes by the two antibodies was similar.
Discussion
By using two different antibodies directed against α-DG, indirect immunofluorescence, confocal microscopy, and IEM, with pre- and post-embedding, we demonstrated that α-DG is localized both at the apical and at the basolateral side of both rat and human podocytes, although staining at the apical membrane was more prominent. The staining of α-DG at the basolateral membrane of podocytes by MoAb IIH6 was less intense than was staining with VIA4.1. This could be due to the fact that the binding of this antibody at the basolateral membrane is sterically hindered by binding of α-DG to laminin G modules. Alternatively, the epitope could be part of the binding site, because it has been shown that IIH6 can block the binding of α-DG to laminin G modules (Durbeej et al. 1995; Matsumura et al. 1997). These laminin G modules are abundantly available in laminin, agrin, and perlecan, which are present in the GBM. The presence of α-DG on the basolateral side of the podocyte cell membrane can therefore be more easily visualized by MoAb VIA4.1, because this MoAb is unable to block the binding of α-DG to laminin G modules. Therefore, we think that the predominant basolateral localization in human kidney biopsies as found by Regele et al. (2000) is due to the fact that they used MoAb VIA4.1 and the post-embedding technique. Although the two anti-α-DG antibodies are most likely directed to different epitopes, IIH6 was able to inhibit the binding of VIA4.1, while VIA4.1 did not interfere with the binding of IIH6. This may be due to the fact that MoAb IIH6 is an IgM antibody, which, with its higher molecular mass, could block the epitope of MoAb VIA4.1, to some extent, by sterical hindrance. Because MoAb VIA4.1 is an IgG antibody, this strict hindrance of the epitope of IIH6 does not occur, which implies that they do not share the same epitope. In tubuli, a more pronounced staining by MoAb VIA4.1 was observed, distinct from the TBM. More proximal to the TBM, IIH6 shows a weak staining. This difference in staining implies that either glomerular α-DG has a different glycosylation compared with tubular α-DG or that the epitope of MoAb IIH6 in the tubuli is blocked by one of the ligands of α-DG.
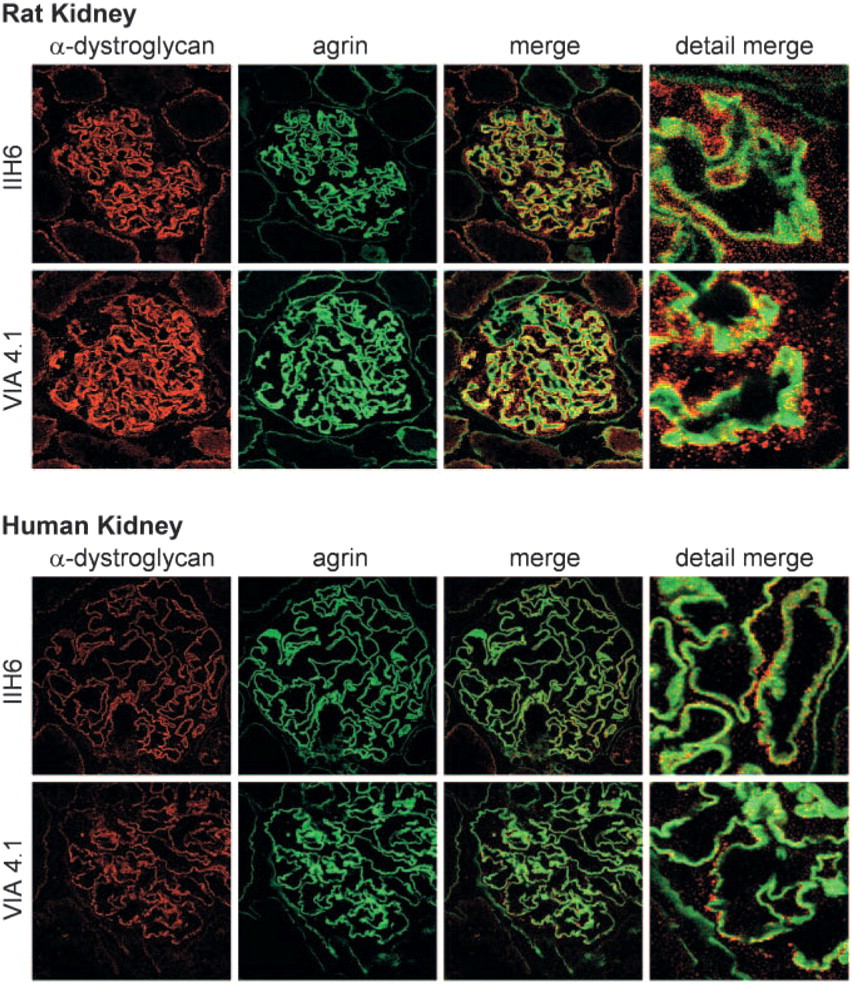
Colocalization of α-DG and agrin in rat and human kidney. Confocal microscopy of rat and human kidney sections, stained with MoAb MI90 specific for agrin (green), and MoAbs VIA4.1 and IIH6 specific for α-DG (red). Most staining of α-DG was observed outside the glomerular basement membrane (GBM), as revealed by both antibodies. This staining indicates that the predominant localization of α-DG is at the apical cell membrane of the podocyte. From the merge, especially in rat kidney sections, it is clear that there is also some staining at the GBM (yellow), which is consistent with a basolateral localization of α-DG (magnification ×630). Isotype controls were negative (not shown).
Expression of α-DG on the podocyte in rat and human kidney as revealed by staining with MoAbs IIH6 and VIA4.1 in immunoelectron microscopy after pre-and post-embedding, on a scale of - to + + + +
Until now, the proposed function of glomerular α-DG has been that it provides a link between the actin cytoskeleton of the podocyte and the GBM (Raats et al. 2000; Regele et al. 2000; Kretzler 2002; Luimula et al. 2002), which only holds for the basolaterally localized α-DG. Because we provide here compelling evidence for the apical localization of α-DG, extra functions should be considered. α-DG has a high sialic acid content, which accounts for its negative charge (Chiba et al. 1997; Ervasti et al. 1997; Sasaki et al. 1998; Smalheiser et al. 1998). Furthermore, sialic acids have been found extensively on the apical membrane of podocytes, and podocalyxin has been regarded as the major protein carrying these sialic acid-rich residues (Faraggiana et al. 1982; Kerjaschki et al. 1984; Charest and Roth 1985; Holthofer et al. 1988). This negative charge enables podocytes to maintain the integrity of their unique filtration slits. In vivo masking of this negative charge by protamin sulfate or hexadimethrine leads to immediate foot process effacement, which can be prevented by scavenging these positively charged molecules with heparin (Kerjaschki 1978, 1994; Bridges et al. 1991). It has also been shown that enzymatic removal of sialic acid residues with neuraminidase (sialidase) leads to foot process effacement and proteinuria (Andrews 1979; Kanwar and Farquhar 1980; Gelberg et al. 1996). In addition, this coating with sialic acid residues prevents adhesion with parietal epithelial cells, thereby preventing synechia formation (Smeets et al. 2004). We hypothesize that these anti-adhesive properties are in part due to the negative charge of the sialic acid residues linked to glomerular α-DG and that therefore apical glomerular α-DG has a function similar to that of podocalyxin in this respect. Foot process effacement is a major reaction of podocytes in many proteinuric glomerular diseases, such as minimal change nephropathy and experimental adriamycin nephropathy, in which the expression of glomerular α-DG is decreased (Raats et al. 2000; Regele et al. 2000). It has also been proposed that retraction of foot processes can be induced by outside-in signaling events. Because α-DG is known for its ability to bind several ligands in the extracellular matrix, it may also have a function in signaling events. Indeed, preliminary work from our laboratory shows that α-DG on podocytes is involved in calcium signaling.
In conclusion, the glomerular anionic glycoprotein α-DG has apical as well as basolateral localization and may be involved in the maintenance of the unique podocyte architecture by acting as an adhesion molecule at the basolateral side and as an anti-adhesion molecule at the cell membranes facing the filtration slit and Bowman's space, respectively.
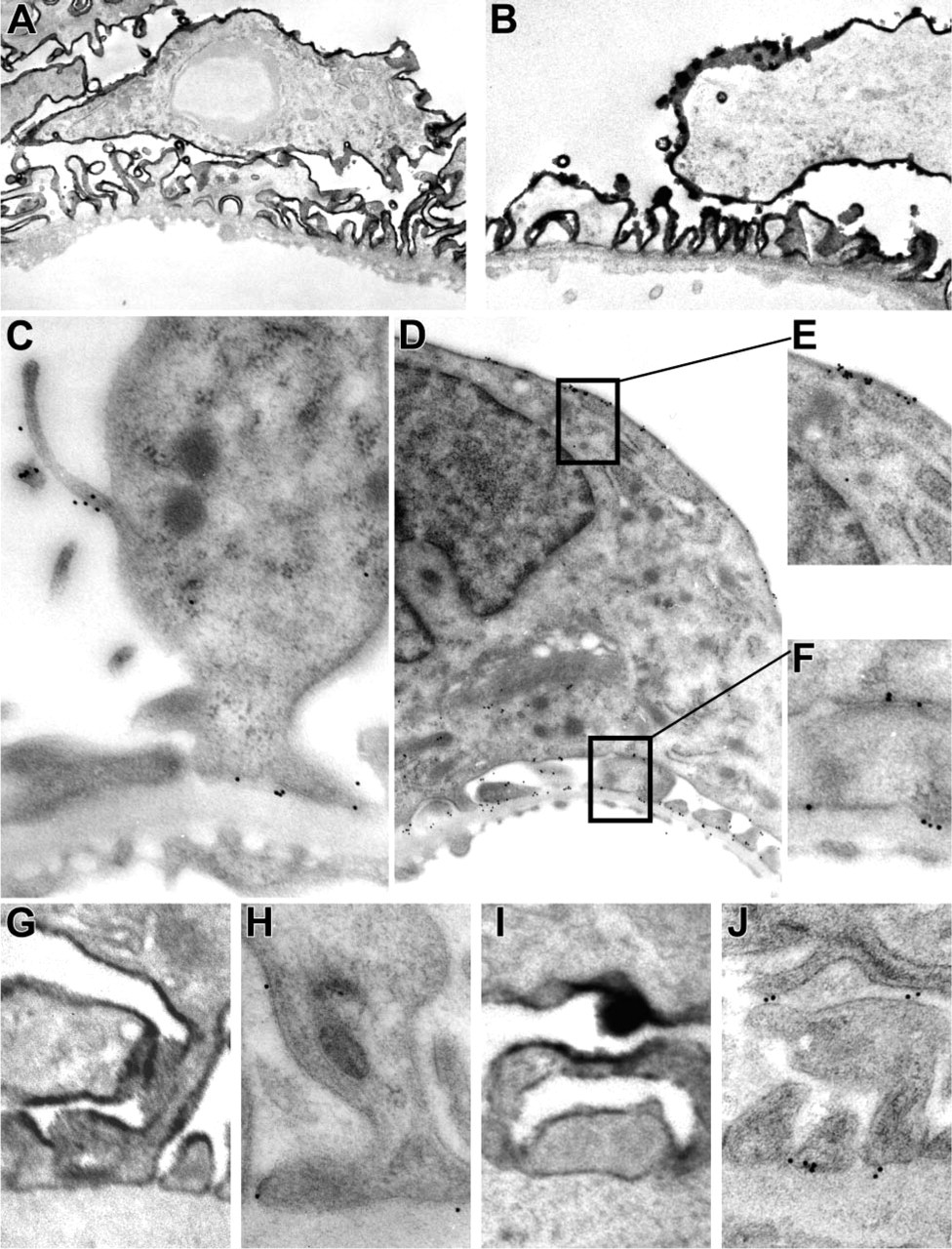
Localization of α-DG in rat and human kidney sections by immunoelectron microscopy (IEM), using pre- or post-embedding techniques. IEM was performed on rat (
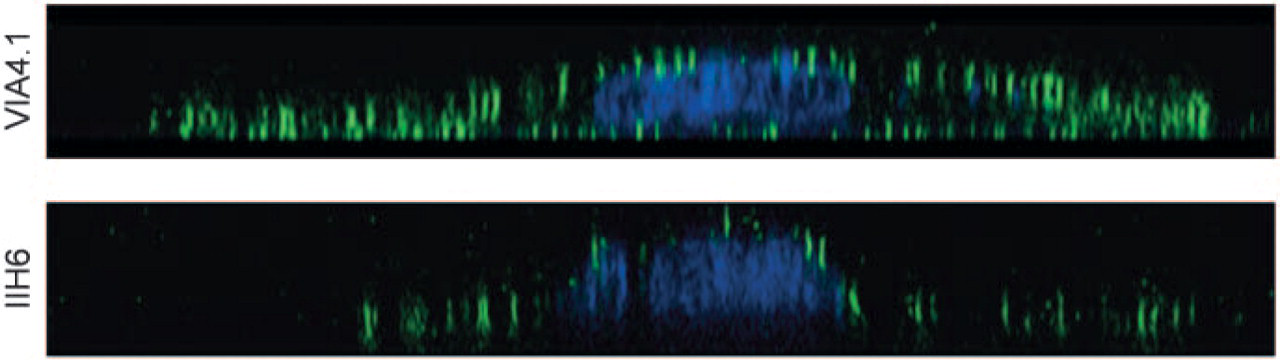
Localization of α-DG in in vitro differentiated immortalized mouse podocytes. Podocytes differentiated on Matrigel expressed α-DG (green) as revealed by staining with MoAb VIA4.1 (upper panel) and MoAb IIH6 (lower panel). The nucleus was probed with topro (blue). Shown are Z scans revealing the basolateral and apical expression of α-DG. It is clear that MoAb VIA4.1 has a better capacity to stain α-DG at the basolateral side of the podocyte (magnification ×630). Isotype controls were negative (not shown).
Footnotes
Acknowledgements
The financial support of the Dutch Kidney Foundation (grant C.99.1832) is gratefully acknowledged. The MPC-5 mouse podocyte cell line was kindly provided by Dr. Peter Mundel, Division of Nephrology, Albert Einstein College of Medicine, Bronx, NY.