
Abstract
S-adenosylhomocysteine (SAH) hydrolase is a cytosolic enzyme present in the kidney. Enzyme activities of SAH hydrolase were measured in the kidney in isolated glomeruli and tubules. SAH hydrolase activity was 0.62 ± 0.02 mU/mg in the kidney, 0.32 ± 0.03 mU/mg in the glomeruli, and 0.50 ± 0.02 mU/mg in isolated tubules. Using immunohistochemical methods, we describe the localization of the enzyme SAH hydrolase in rat kidney with a highly specific antibody raised in rabbits against purified SAH hydrolase from bovine kidney. This antibody crossreacts to almost the same extent with the SAH hydrolase from different species such as rat, pig, and human. Using light microscopy, SAH hydrolase was visualized by the biotin-streptavidin-alkaline phosphatase immunohistochemical procedure. SAH hydrolase immunostaining was observed in glomeruli and in the epithelium of the proximal and distal tubules. The collecting ducts of the cortex and medulla were homogeneously stained. By using double immunofluorescence staining and two-channel immunofluorescence confocal laser scanning microscopy, we differentiated the glomerular cells (endothelium, mesangium, podocytes) and found intensive staining of podocytes. Our results show that the enzyme SAH hydrolase is found ubiquitously in the rat kidney. The prominent staining of SAH hydrolase in the podocytes may reflect high rates of transmethylation.
Keywords
T
The distribution of SAH hydrolase in various tissues of the rat was investigated by Eloranta (1977). The highest activity was found in the liver and pancreas. The kidney also contains high levels of SAH hydrolase, i.e., the activity is 40% of the values for the liver and pancreas. Intermediate values of enzyme activity are reported for the brain and low activities in the heart, testes, muscle, prostate, spleen, and lung (Finkelstein and Harris 1973; Eloranta 1977). Small variations in enzyme activity and localization are observed among different regions of the rat brain (Broch and Ueland 1980; Patel and Tudball 1986). These data suggest that SAH hydrolase is ubiquitously distributed in animal tissues. As previously reported, we have characterized the enzyme activity and adenosine binding feature of renal SAH hydrolase (Kloor et al 1996). The kidney is highly heterogeneous in structure and function. Therefore, it is of interest to analyze the distribution of SAH hydrolase in the different cell types of this organ. Moreover, no information is available on whether enzyme activity is restricted to a particular cell type in the kidney. For this reason, we examined the SAH hydrolase activity in glomeruli and tubular fragments of rat kidney, using immunohistochemical methods.
Materials and Methods
Materials
The following materials were purchased from the different sources indicated: Protran B 85 nitrocellulose membrane (Schleicher & Schuell, Dusseldorf, Germany); biotinylated antibody against rabbit IgG, streptavidin-alkaline phosphatase-biotin (ABC) complex, new fuchsin chromogen, 0.1% avidin, 0.01% biotin (Dako; Glostrup, Denmark); monoclonal mouse anti-rat THY-1 antibody (Seikagaku; Tokyo, Japan); mouse anti-human von Willebrand factor (vWF), adenosine deaminase, nucleoside phosphorylase, xanthin oxidase, and clostridiopeptidase A (Boehringer Mannheim; Mannheim, Germany); anti-mouse and anti-rabbit antibodies labeled with Texas Red and Cy5 (Dianova; Hamburg, Germany; SAH, bovine serum albumin (BSA), and 4-chloro-1-naphthol (Sigma; St Louis, MO); O-toluidine (Aldrich; Milwaukee, WI); MTT (3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazoliumbromide) (Serva; Heidelberg, Germany). All other reagents were of analytical grade and were obtained from Merck (Darmstadt, Germany).
Human kidney tissue was obtained according to the Helsinki guidelines from patients who had undergone nephrectomy because of renal cell carcinoma in the Department of Surgery at Robert-Bosch Hospital, Stuttgart.
Enzyme Purification
SAH hydrolase was purified from bovine kidney using chromatographical techniques as described previously (Kloor et al. 1996). The purified enzyme was frozen at −20C until use.
Protein Assay
Protein concentration was determined according to the method of Bradford (1976), using bovine serum albumin as a standard.
Isolation of Glomeruli
Glomeruli were isolated using a modified sieving method according to Misra (1972). The capsules of the kidneys were removed and each kidney was bisected longitudinally. The medulla was removed by macroscopic dissection and the cortical tissue was minced using the same scissors. All further isolation steps were performed at 4C. The tissue was then gently pushed through a 315-μm sieve. The material on the sieve was washed with physiological saline solution and the suspension was collected in a siliconized glass tube. The suspension was then sieved stepwise through 280-μm, 200-μm, and 125-μm sieves. The glomeruli were recovered on a 63-μm sieve.
Isolation of Tubules
Tubules were isolated by collagenase digestion. The kidney was diced with scissors and incubated for 45 min at room temperature (RT) in a 0.4% collagenase solution (115 mM NaCl, 5 mM KCl, 10 mM Na-acetate, 1.2 mM NaH2PO4, 1.2 mM MgSO4, 1 mM CaCl2, and 25 mM NaHCO3). After the incubation, the mixture was sieved through a 320-μm sieve and washed with physiological saline solution to remove adherent collagenase. The tubules were collected after centrifugation at 1000 times; g for 20 min.
Enzyme Assay
SAH hydrolase activity was assayed in the crude homogenate in the hydrolytic direction. The reaction mixture (1 ml) contained 50 mM potassium phosphate, pH 7.0, 50 μl homogenate, 1 mM MTT, 0.14% gelatin, 1.0 U adenosine deaminase, 0.8 U nucleoside phosphorylase, and 0.8 U xanthine oxidase. This reaction mixture was incubated for 5 min at RT to remove all endogenous substrates that might react with the added enzymes. After this incubation, the reaction for specific determination of SAH hydrolase activity was started by the addition of 80 μM SAH. The formation of formazan was measured at 578 nm.
Preparation of Antibodies
Antibody against bovine SAH hydrolase was raised in rabbits by multisite SC injections of an emulsion of the enzyme (100-150 μg protein) prepared by mixing equal volumes of protein and Freund's complete adjuvant (Sigma). After 4 and 8 weeks, the animal received an SC booster injection of 0.5 mg enzyme prepared as above with incomplete Freund's adjuvant (Sigma). The animals were bled 7 days after the last injection. Serum was centrifuged at 3500 times; g for 20 min and stored at −20C. The specificity of the antibody was examined by testing its ability to precipitate enzyme activity from SAH hydrolase and by Western blotting.
Western Blot Analysis
Rat, bovine, and human kidney tissues were homogenized in 2 volumes of 8% sucrose containing 50 mM potassium phosphate, 1 mM DTT, 1 mM EDTA, and 100 μM PMSF and were centrifuged at 50,000 times; g for 60 min at 4C. The supernatants were separated by SDS-PAGE according to Laemmli (1970) using 7-19% polyacrylamide gradient gels. After electrophoresis, the fractionated proteins were electrophoretically blotted onto nitrocellulose filters for 2 hr at 4C using constant current (400 mA) in 25 mM Tris, 190 mM glycine, 10% (v/v) methanol, pH 8.2. The filters were blocked overnight with 5% skim milk powder in PBS (150 mM NaCl, 50 mM KH2PO4, 20 mM Na2HPO4, pH 7.2). The filters were incubated for 1 hr at RT with the first antibody in PBS in a 1:250 dilution containing 1% BSA. The blots were washed with TPBS (PBS with 0.05% Tween-20) three times for 10 min to remove unbound antibodies and were incubated for 1 hr at RT with a second antibody (goat anti-rabbit peroxidase-conjugated IgG) in PBS at 1:1000 dilution containing 1% BSA. After washing with TPBS (three times for 10 min), the immunoreactive bands were visualized by O-toluidine-dependent POD (peroxidase) reaction (100 μl 4-chloro-1-naphthol 2% w/v in diethyleneglycol, 500 μl O-toluidine saturated in 7% acetic acid, 500 μl 3% H2O2 in 50 ml PBS).
Immunoprecipitation of Purified SAH Hydrolase
SAH hydrolase from bovine and rat kidney (50 μg/ml) was incubated with 10, 25, 50, 75, 100, 125, and 150 μl anti-SAH hydrolase sera. The SAH hydrolase activity was assayed in the hydrolytic direction as mentioned above and was expressed as a percentage of the residual enzyme activity.
Immunohistochemistry
Immunohistochemistry was performed using the biotin-streptavidin-alkaline phosphatase complex technique. Cryostat sections from kidneys of Sprague-Dawley rats (10 μm) were fixed with acetone for 10 min at RT and air-dried. The sections were washed for 30 min in TBS (150 mM NaCl, 50 mM Tris-HCl, pH 7.6). Nonspecific background staining was reduced by preincubating sections for 1 hr with 5% normal goat serum at RT. The sections were incubated with the polyclonal antibody against SAH hydrolase (1:100 dilution) overnight at RT. After rinsing, the sections were incubated with the biotinylated antibody (1:300 dilution) against rabbit IgG for 1 hr at RT. The sections were washed in TBS and incubated for 10 min with 0.1% avidin in 50 mM Tris-HCl, pH 7.6, to reduce the endogenous biotin. The avidin solution was then rinsed off and the slices were incubated for 10 min with 0.01% biotin in 50 mM Tris-HCl, pH 7.6 (Woods and Warnke 1981). Then the sections were incubated with biotin-streptavidin-alkaline phosphatase complex for 30 min at RT. Color was developed with 0.05% new fuchsin in TBS and 0.03% naphthol-phosphate dissolved in N,N-dimethylformamide. To reduce endogenous alkaline phosphatase activity, the color developing mixture contained 1 mM levamisole. The slices were counterstained with hematoxylin, mounted with a water-soluble coverslip, and photographed with a light microscope (Zeiss; Oberkochen, Germany). Control slices were prepared by using antiserum preabsorbed with SAH hydrolase.
For the double-immunofluorescence staining study, we used antiserum against SAH hydrolase, a mouse monoclonal antibody (MAb) against THY-1, and a mouse MAb against vWF. Sections were incubated simultaneously with a cocktail of antibodies against SAH hydrolase (1:100 dilution) and against THY 1 (1:150 dilution), and with a cocktail of antibodies against SAH hydrolase (1:100 dilution) and against vWF (1:100 dilution). Then the slices were incubated with indodicarbocyanin (Cy5) conjugated anti-rabbit IgG and with Texas Red-conjugated anti-mouse IgG for 1 hr at RT. The sections were examined with a two-channel fluorescence confocal LSM. The crossreactivity of the anti-mouse IgG with the rabbit IgG and the anti-rabbit IgG with the mouse IgG was determined by dot-blot analysis. To evaluate the background staining, sections were stained only with the fluorescent conjugates.
Data Calculation and Statistics
Data are expressed as mean value ± SEM. For statistical analysis, Student's t-test for unpaired data was used and p values of <0.05 were considered to be significant.
Results
The isolation of distinct morphological regions of the kidney with different physiological functions allows a quantitative assessment of the distribution of SAH hydrolase activity. With the sieving method, the glomeruli were clearly separated from tubule fragments and cells. The purity of the glomerular preparation was about 96%. The sieving procedure removed the Bowman capsule from glomeruli. The average diameter of glomeruli was 100 μm. Tubules were isolated by collagenase digestion of rat kidney. The purity of the tubule fragments was about 85%.
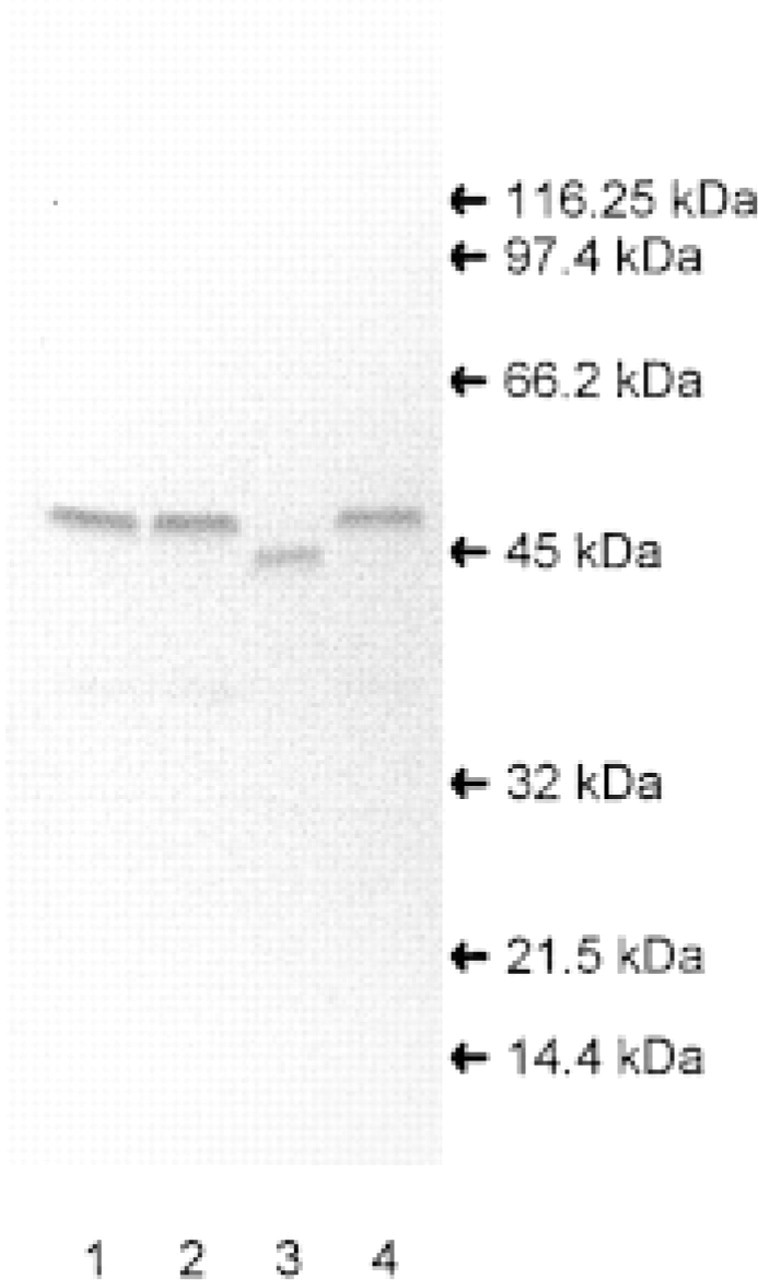
Western blot of SAH hydrolase from kidneys. Kidney homogenate (100 μg/lane) was subjected to SDS-PAGE followed by immunoblotting with anti-bovine SAH hydrolase. Lane 1, human; Lane 2, pig; Lane 3, rat; Lane 4, bovine. Molecular mass markers used are β-galactosidase (116.25 kD), phosphorylase B (97.4 kD), albumin (66.2 kD), ovalbumin (45 kD), carbonic anhydrase (32 kD), trypsin inhibitor (21.5 kD), and lysozyme (14.4 kD).
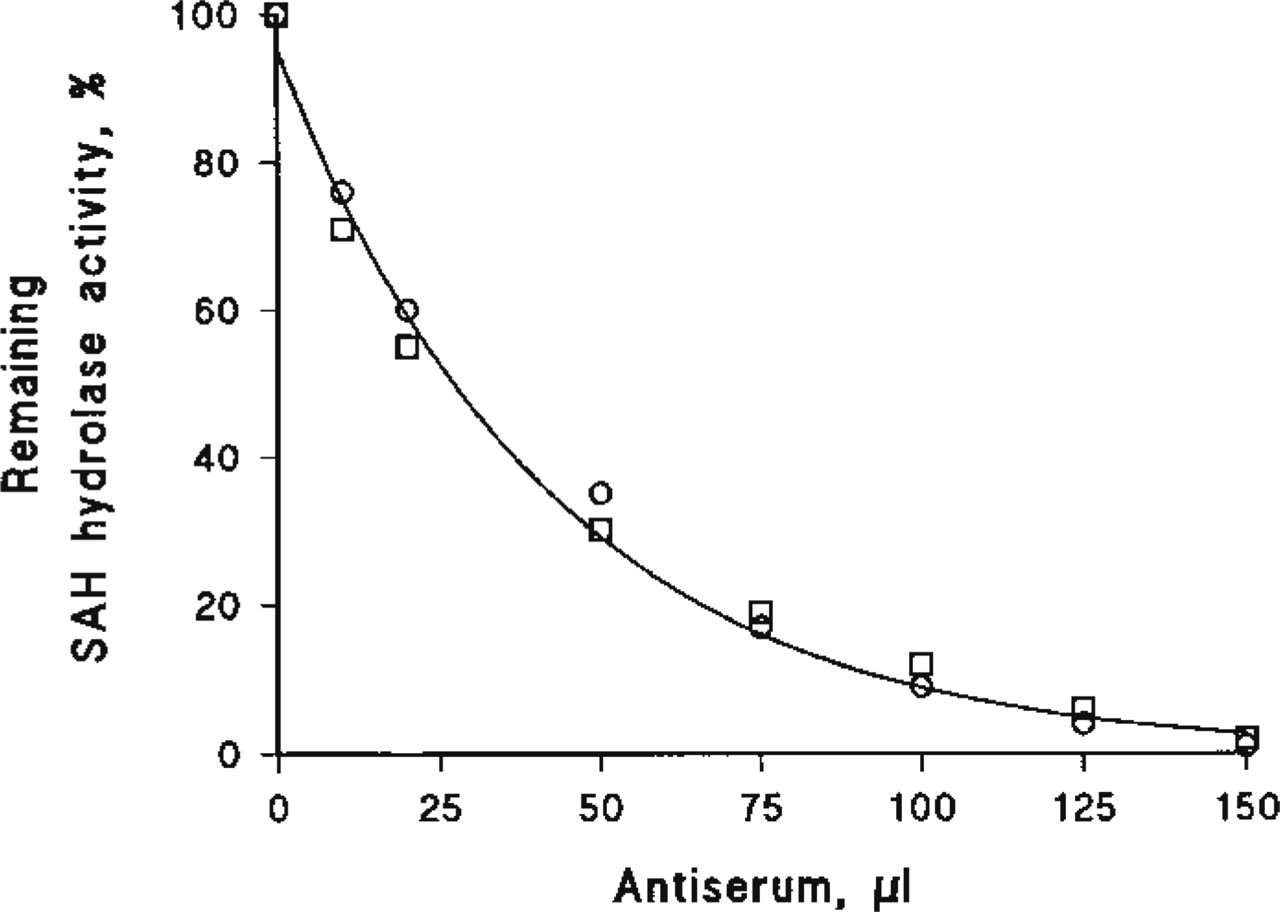
Immunological titration of SAH hydrolase activity from bovine □ and rat ○ kidney. SAH hydrolase (50 μl) was incubated with the indicated amounts of the antiserum at RT in 50 mM potassium phosphate, pH 7.0. After 20-min incubation, residual activity of SAH hydrolase was measured in the direction of hydrolysis (n = 2).
The specific enzymatic activity of SAH hydrolase determined in the entire kidney was 0.62 ± 0.02 mU/mg (n =13). The SAH hydrolase activity of the glomerular preparation was 0.32 ± 0.03 mU/mg (n = 11) and was therefore 50% lower than the activity of the entire kidney, whereas the tubule preparation showed an enzyme activity of 0.50 ± 0.02 mU/mg (n = 10).
To localize SAH hydrolase at the cellular level in the kidney, we performed immunohistochemistry on cryostat sections of rat kidney. The specificity of the anti-SAH hydrolase antibody used in this study and the ability of the antiserum prepared against the purified SAH hydrolase from bovine kidney to crossreact with the SAH hydrolase from different species were examined by Western blot analysis of kidney homogenates from bovine, pig, rat, and human (Figure 1). The anti-bovine SAH hydrolase antibody from bovine kidneys reacted only with a single band in all the kidney extracts examined. The molecular weights of the SAH hydrolase monomers for bovine, pig, and human kidney were similar, with a mass of 47.5 kD, whereas the monomer of rat SAH hydrolase showed a band at 45 kD. The immunological titration of SAH hydrolase activity from bovine and rat kidney with the antiserum (Figure 2) revealed complete inactivation of enzyme activity.
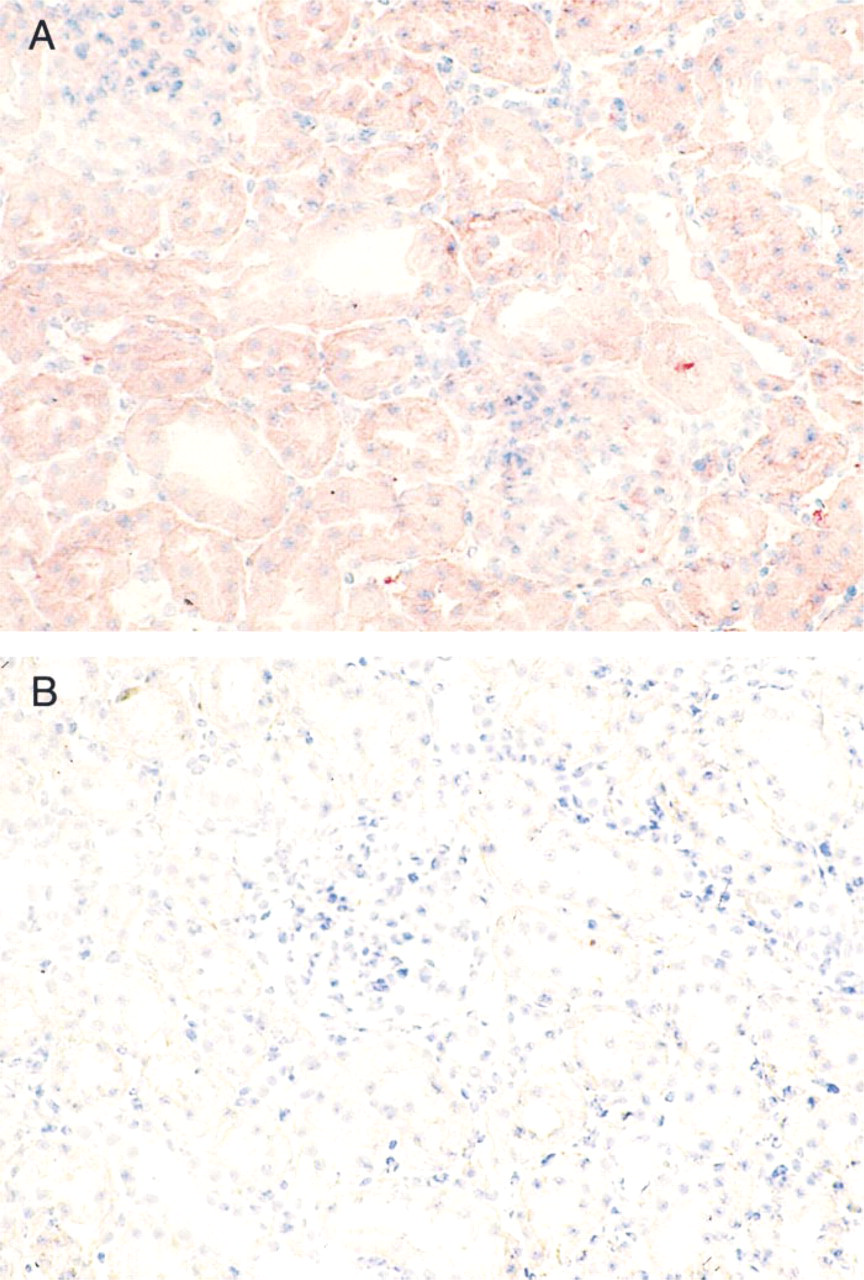
Light micrographs illustrate the immunohistochemical localization of SAH hydrolase in rat kidney. Immunostaining for SAH hydrolase is observed in the glomeruli and in the epithelium of the proximal and distal tubules (
Immunostaining of SAH hydrolase appeared to be relatively uniform (Figure 3). In the cortex, homogeneous staining was observed in the epithelium of proximal and distal tubules (Figure 3A). In addition, collecting ducts and loops of Henle from the medulla were homogeneously stained. Glomeruli showed the same staining intensity compared with the surrounding tubules. The staining was completely blocked by preincubation of the antibody with the SAH hydrolase (Figure 3B).
To differentiate the type of cells that express SAH hydrolase in the glomeruli, double labeling immunofluorescence was used with (a) the polyclonal antibody against SAH hydrolase and the MAb against THY-1 and (b) the polyclonal antibody against SAH hydrolase and the MAb against von Willebrand factor. Anti-THY-1 antibody binds to THY-1 antigen on rat glomerular mesangial cells (Hugo et al. 1997). Anti-von Willebrand factor binds to an endothelial cell antigen.
Figure 4 shows fluorescence images of rat glomeruli, depicting the distributions of SAH hydrolase (green) and THY-1 (red). Staining for SAH hydrolase (green) was detectable in cells that were characterized as mesangial cells by THY-1 staining (red), and resulted in an orange aspect (Figure 4C). The same was done for endothelial cells marked by anti-von Willebrand factor (Figure 5). In addition, both double staining procedures showed a cell type that was positive only for anti SAH hydrolase. This green staining (Figures 4 and 5) was concluded to represent the podocytes.
Discussion
This study examined the distribution of SAH hydrolase in normal rat kidney by two different methods: assessment of enzymatic activity and immunohistochemistry.
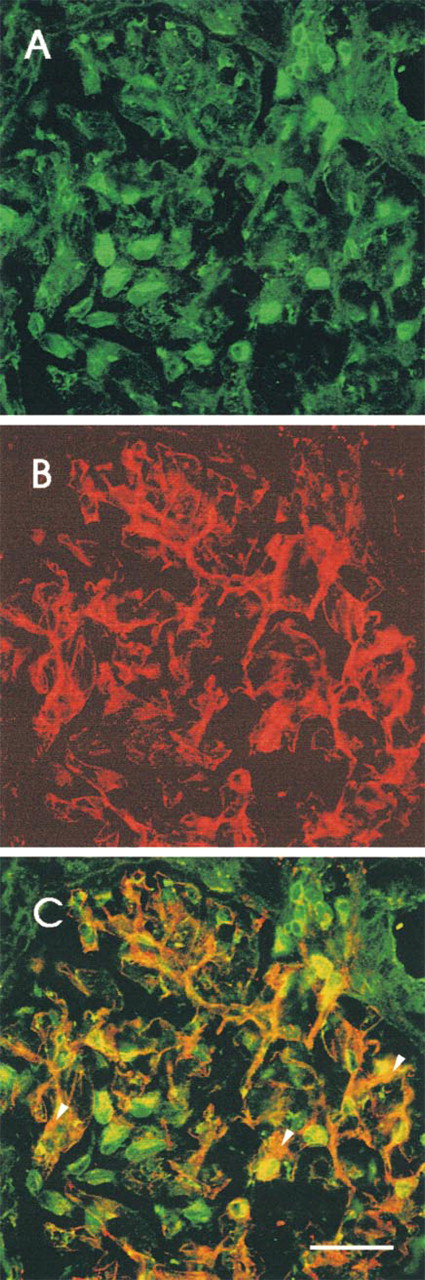
Indirect immunofluorescence micrographs showing a rat glomerulus. (
SAH hydrolase activity was determined in the whole kidney and in isolated glomeruli and tubules. SAH hydrolase activity of isolated tubules is slightly but not significantly lower than that of the whole-kidney homogenate. At least two factors may be responsible for this difference. First, a partial loss of cellular integrity may be due to the isolation procedure used, which led to leakage of the cytoplasmatic enzyme. Second, the blood vessels that showed more intense staining were removed by the isolation procedure. In addition, the similar enzyme activities in tubules and the whole-kidney homogenate imply that cells of the renal medulla are similarly equipped with SAH hyrolase activity because the preparation of tubules contained mainly cortical tissue. Because the glomeruli contribute to whole-kidney protein by only 4.9% (Seyer-Hansen et al. 1980), lower whole-kidney activity compared to the activity of tubules would not be expected.
By Western blotting, the polyclonal antibody against the purified SAH hydrolase from bovine kidney reacted with the enzyme that was used for immunization and showed a crossreaction with the SAH hydrolase from pig, rat, and human kidney. This result is in accordance with the structural similarities of the SAH hydrolase in different species. The amino acid sequences for rat liver and human placenta SAH hydrolase are reported to be 97% homologous (Coulter-Karis and Hershfield 1989). Therefore, our polyclonal antibody is suited for immunohistochemical localization of SAH hydrolase in rat kidney.
SAH hydrolase immunostaining shows a homogeneous distribution of the enzyme in the rat kidney. The activity of SAH hydrolase in glomeruli is 1.5-fold that in tubules. The ratio of the reduced to the oxidized form of the nucleotide (NADH/NAD+) on SAH hydrolase in the cells might be responsible for the difference in staining intensity and enzyme activity of SAH hydrolase. The antibody identifies the enzyme through several epitopes, whereas measurement of enzyme activity can determine only the rate of SAH hydrolysis but not the SAH hydrolase protein concentration. As has previously been shown (Kloor et al. 1998), renal SAH hydrolase activity is controlled by the ratio of NAD+/NADH that is tightly bound to the enzyme. The NADH form of SAH hydrolase is enzymatically inactive. The binding of the antibody cannot differentiate between the two forms of the enzyme (NAD+/NADH), and therefore it is not possible to compare staining intensity and enzyme activity.
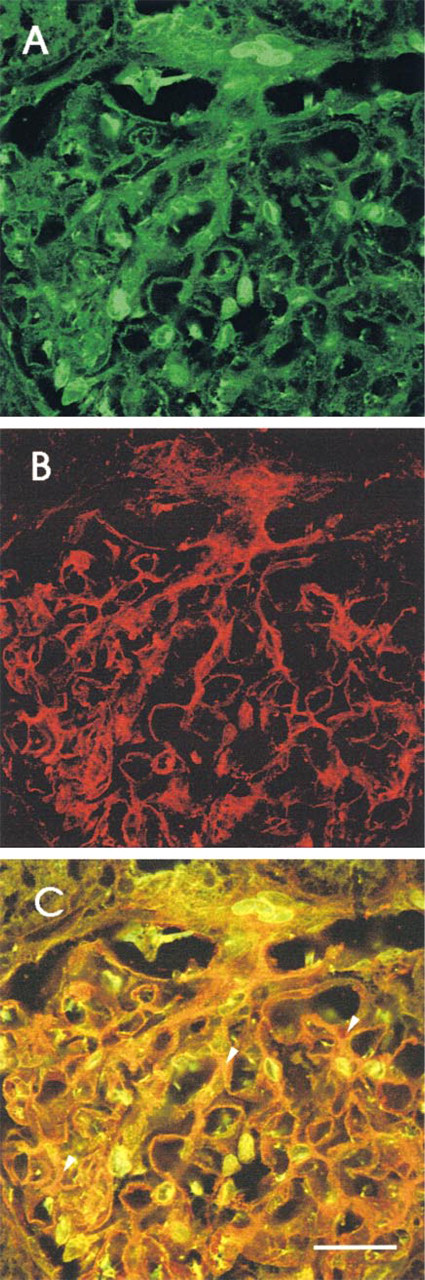
Indirect immunofluorescence micrograph showing a rat glomerulus. (
To differentiate the SAH hydrolase content in the different cells of glomeruli, cryosections of a whole kidney were examined by LS microscopy. To discriminate more precisely between podocytes and mesangium, double staining was used, and it was possible to clearly demonstrate that mesangial cells also express SAH hydrolase. To further discriminate between endothelial cells and podocytes in single glomerulus capillary loops, double staining with von Willebrand factor and SAH hydrolase was used. The result clearly shows that SAH hydrolase is also present in endothelial cells. These observations, even in the absence of podocyte double staining, strongly favor the conclusion that the intense green staining shown in Figures 4C and 5C represents other cells than podocytes. However, evidence for podocytes is provided by the localization of these cells outside the glomerular interstitium and near the glomerular basement membrane.
Podocytes stabilize the folding pattern of glomerular capillaries by fixing the turning points of the glomerular basement membrane to neighboring capillaries (Kriz et al. 1995). Podocytes contain large numbers of microtubules, microfilaments, and intermediate filaments in the cytoplasm, and actin filaments are especially abundant in the pedicels (Andrews and Bates 1984; Vasmant et al. 1984; Buniatian et al. 1998). Podocytes have several functions, including glomerular basement membrane turnover, maintenance of the glomerular filtration barrier, regulation of glomerular filtration, and immunological functions (Mundel and Kriz 1995). The high content of SAH hydrolase in the podocytes might be indicative of higher turnover in transmethylation reactions.
For the possible physiological role of SAH hydrolase, it is important to mention that protein methylation in the kidney may be involved in the action of aldosterone on tubules (Bastl and Hayslett 1992). When SAH hydrolase is overexpressed in oocytes to lower cytosolic SAH levels, aldosterone action was enhanced, whereas inhibition of SAH hydrolase resulted in almost complete obviation of aldosterone action (Stockand et al. 1998). These observations support the concept that SAH concentrations in the cytosol participate in the transmethylation activity. Whether increased transmethylation activity is associated with enhanced SAH hydrolase activity in vivo under different conditions of electrolyte transport (e.g., high-potassium diet, low-sodium diet) remains to be determined.
The data show that SAH hydrolase activity is present in the whole kidney, glomeruli, and tubules. Differentiation of glomeruli cells by double staining revealed that SAH hydrolase immunoreactivity is present in endothelial cells, mesangial cells, and podocytes. Further study is required to determine whether different transmethylation activities in tubules under physiological and pathophysiological conditions are associated with different distribution patterns of SAH hydrolase in the kidney.
Footnotes
Acknowledgments
Supported by a grant from the Federal Ministry of Education, Science, Research and Technology (Fö. 01 KS 9602) and by the Interdisciplinary Clinical Research Center (IKFZ) Tübingen.