
Abstract
Monoamine storage in secretory granules is mediated by the vesicular monoamine transporters 1 and 2 (VMAT1 and VMAT2). The aim of our study was to identify monoamine-handling normal and neoplastic inflammatory cells in the skin by their expression of VMAT1 and VMAT2. Normal skin from various parts of the body, as well as 21 cases of cutaneous mastocytosis and 10 cases of cutaneous Langerhans cell histiocytosis were analyzed by immunohistochemistry, radioactive in situ hybridization, and double-fluorescence confocal microscopy. VMAT2-positive cells in the subepidermal layer were identified as mast cells by their expression of tryptase. Neoplastic mast cells in all cases of cutaneous mastocytosis retained their VMAT2 positivity. The intraepidermal VMAT2-expressing cells were identified as Langerhans cells by their CD1a positivity. VMAT2 was absent from Langerhans cell histiocytosis. VMAT2 is an excellent marker for normal and neoplastic mast cells. The expression of VMAT2 demonstrates the capacity of mast cells for monoamine storage and handling. The presence of VMAT2 in epidermal Langerhans cells revealed a previously unrecognized monoamine-handling phenotype of these cells and indicates possible involvement of amine storage and release associated with antigen presentation. Absence of VMAT2 in neoplastic Langerhans cells indicates a loss of monoamine handling capacity of these cells during tumorigenesis. (
Keywords
M
The uptake of monoamines from the cytoplasm into secretory granules is mediated by vesicular monoamine transporters (VMATs) along an ATPase-generated proton gradient (Schulinger et al. 1995; Erickson et al. 1996; Eiden 2000; Erickson and Varoqui 2000; Travis et al. 2000). Two isoforms of the vesicular monoamine transporter (VMAT1 and VMAT2) were isolated from rat and human tissue (Erickson et al. 1992; Liu et al. 1992; Erickson and Eiden 1993).
Extensive studies employing immunohistochemistry (IHC) and in situ hybridization (ISH) have shown that these two transporters are differentially expressed in monoamine-handling cells of the central nervous system, the autonomic nervous system, and in neuroendocrine cells and related neuroendocrine tumors (Weihe et al. 1994; Schutz et al. 1998; Eissele et al. 1999; Weihe and Eiden 2000; Jakobsen et al. 2001; Eiden et al. 2002; Anlauf et al. 2003a, b). In addition to their roles as vesicular transporters for monoamines in neurons and neuroendocrine cells, they may also be present in immune cells, because VMAT2 was originally cloned from a rat basophilic leukemia cell line (Erickson et al. 1992). There is some evidence that VMATs may be expressed in blood lymphocytes (Watson et al. 1999; Amenta et al. 2001). The differential in vivo expression of VMATs in cells similar to basophilic cells, such as the mast cells, has not been examined in detail (Weihe and Eiden 2000). It was therefore our aim to analyze the expression of VMAT1 and VMAT2 mRNA and protein in inflammatory cells of the primate skin. Furthermore, to investigate whether VMAT1 and VMAT2 may serve as potential markers for tumors derived from mast cells and Langerhans cells of the skin, a series of cases of cutaneous mastocytosis and of cutaneous Langerhans cell histiocytosis were analyzed for their expression of VMAT1 and VMAT2.
Materials and Methods
Tissue Collection and Preparation
Various samples of human skin were collected from 39 patients during surgery. The patients consisted of 17 men and 22 women with an average age of 61 years and an age range of 4 to 83 years. The expression patterns of VMAT1 and VMAT2 were analyzed in non-tumorous normal human skin in various regions: head (n = 3), nose (n = 3), lip (n = 2), other facial regions (n = 5), neck (n = 2), back (n = 2), thoracic and abdominal skin (n = 3), upper and lower extremities (n = 9), scrotum and prepuce of penis (n = 2), vulva (n = 2), and the anal skin region (n = 4). Furthermore, multiple skin specimens from various regions were obtained from five rhesus monkeys, as described previously (Rausch et al. 1994).
Samples from cases of cutaneous mastocytosis, including patients with urticaria pigmentosa (n = 10), diffuse cutaneous mastocytosis (n = 8), and solitary mastocytoma (n = 3) were examined for their VMAT expression patterns. The patients with cutaneous mastocytosis consisted of nine males and 12 females with an average age of 14 years and an age range of 2 to 69 years. All these patients exhibited the typical clinical signs of cutaneous mastocytosis with its accompanying skin infiltrates. None of these patients exhibited evidence of extracutaneous involvement or leukemia. Furthermore, tissues from 10 children with CD1a-positive Langerhans cell histiocytosis of the skin were examined. These patients consisted of three boys and seven girls, with an average age of 3 years and an age range of 9 months to 5 years.
Adrenal gland tissues from two rhesus monkeys and from two patients who had undergone unilateral adrenalectomy, one suffering from pheochromocytoma (m/63) and one from Conn's syndrome (w/70), were used as control tissues.
For IHC analysis the human tissues were fixed in either Bouin-Hollande fixative, 10% formalin, or 4% formaldehyde/PBS for 24–48 hr and then embedded in paraffin. The monkey tissues were perfused with 4% formaldehyde/PBS before postfixation in Bouin-Hollande for 24–48 hr, as described previously (Rausch et al. 1994).
After dehydration in a graded series of 2-propanol solution, the tissues were embedded in Paraplast Plus (Merck; Darmstadt, Germany). Adjacent sections (3 μm or 7 μm thick) were cut and deparaffinized. Antigen retrieval to increase the sensitivity of immunodetection was performed by heating the sections at 92–95C for 15 min in 0.01 M citrate buffer (pH 6) according to the DAKO protocol (Hamburg, Germany). Nonspecific binding sites were blocked with 5% bovine serum albumin (BSA; Serva, Heidelberg, Germany) in PBS (pH 7.45), followed by an avidin-biotin blocking step (avidin–biotin blocking kit; Boehringer, Ingelheim, Germany).
For ISH, human non-neoplastic skin tissues from six patients and three patients with cutaneous mastocytosis were immediately frozen on dry ice and stored at −80C. Cryosections (14–16 μm thick) were placed on presilanized glass slides, fixed in 4% phosphate-buffered formaldehyde for 60 min, followed by three 10-min washes in 50 mM PBS (pH 7.4). The slides were then briefly rinsed in distilled water, incubated in 0.1 M triethanolamine (pH 8.0) for 1 min and for 10 min in the same solution containing 0.25% v/v acetic anhydride under rapid stirring. They were then quickly rinsed in 2 × SSC, dehydrated in 50% and 70% ethanol, and air-dried.
List of primary antibodies

dilution for immunofluorescence
polyclonal
monoclonal
Immunohistochemistry
Rabbit antiserum VMAT1/10 raised against the C-terminal sequence of human VMAT1 and antiserum 80182 raised against the C-terminal sequence of human VMAT2 were employed for the IHC investigations (Table 1). Poststaining with Giemsa and antisera against histidine decarboxylase (HDC) and tryptase was used to identify mast cells. Langerhans cells of the skin were labeled with their specific marker CD1a (Table 1).
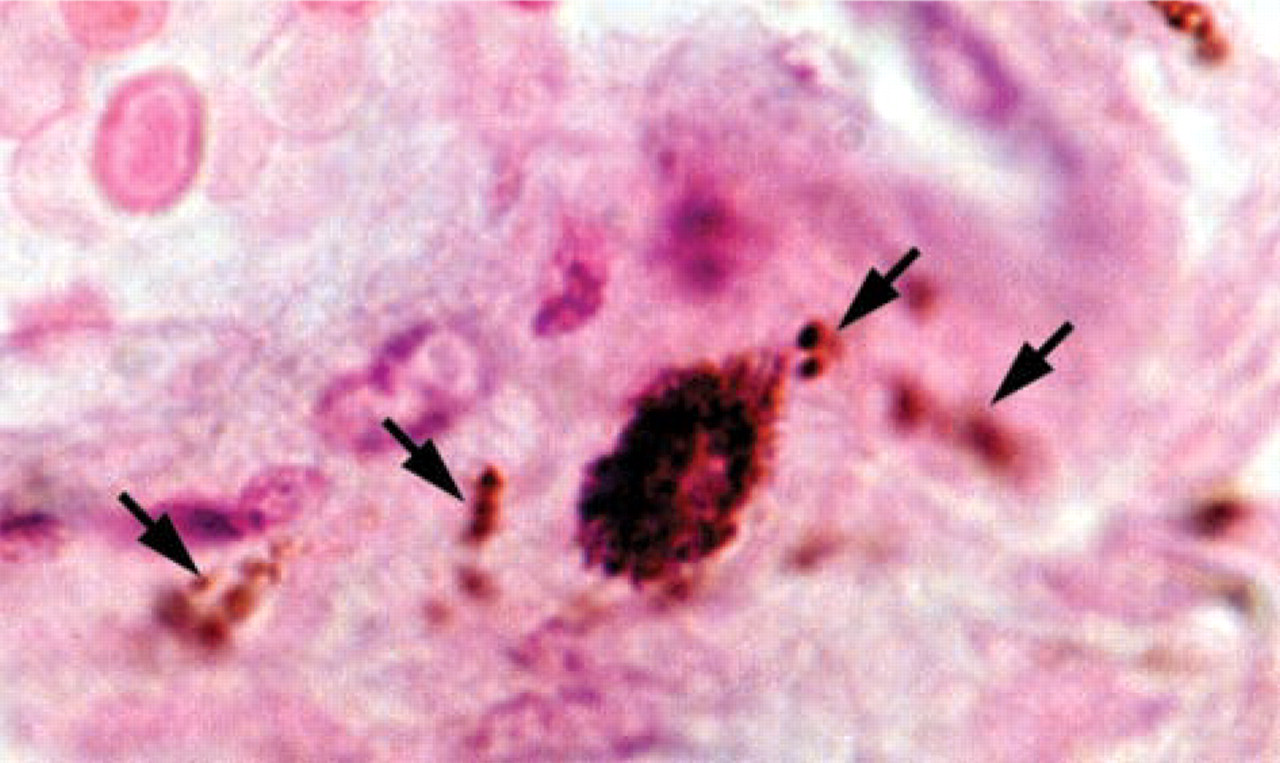
VMAT2 expression in a perivascular immune cell, high-power magnification revealing a granular distribution pattern of VMAT2 in the cytoplasm of a perivascular immune cell. Note that nerve fibers of the perivascular plexus (labeled by arrows) are also positive for VMAT2.
Tissue sections were incubated with the primary antibodies overnight at 18C (diluted as shown in Table 1) and further incubated for 2 hr at 37C. The sections were then washed in distilled water and in 50 mM PBS and incubated with species-specific biotinylated secondary antibodies (Dianova; Hamburg, Germany) for 45 min at 37C, washed several times, and incubated for 30 min with the ABC reagents (Vectastatin Elite ABC kit; Boehringer, Ingelheim, Germany). Immunoreactions were visualized with 3′3-diaminobenzidine (DAB; Sigma, Deisenhofen, Germany) enhanced by the addition of 0.08% ammonium nickel sulfate (Fluka; Buchs, Switzerland), resulting in dark blue staining. No binding was detected in the absence of the primary antibody. The specificity of the IHC staining was observed by preabsorbing the antisera with 25 μmol of the C-terminal human VMAT1 and VMAT2 peptide, respectively.
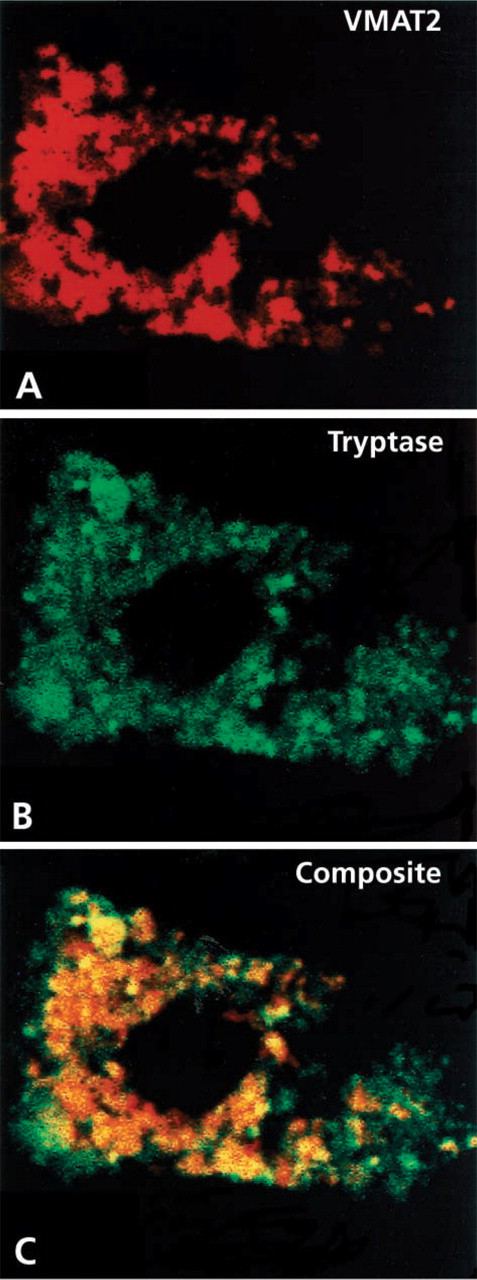
Identification of the cellular phenotype of VMAT2-immunoreactive connective tissue immune cells and analysis of the subcellular distribution of VMAT2 revealed by confocal laser scanning microscopy (
Radioactive ISH
To generate specific probes for the localization of VMAT1 mRNA, a 247-bp DNA restriction fragment of the hVMAT1 cDNA corresponding to nt 436–682 was subcloned into Bluescript II KS+ (Stratagene; Heidelberg, Germany) (Erickson et al. 1996). For VMAT2 mRNA detection, a 269-bp DNA fragment of the hVMAT2 corresponding to nt 244–512 was subcloned into pCDNAI (Invitrogen; Leek, The Netherlands) (Erickson and Eiden 1993). Inserted sequences were verified by double-stranded DNA sequencing. In vitro transcription of the two vector constructs using [35S]-UTP-labeled nucleotide yielded antisense riboprobes for hVMAT1 after linearization with XhoI and incubation with T7 RNA polymerase, and for hVMAT2 after linearization with XbaI and incubation with SP6 RNA polymerase. To increase the tissue penetration of the probes, generated transcripts were reduced to ∼200 nucleotide fragments by limited alkaline hydrolysis, as described by Angerer et al. (1987).
To each section, hybridization buffer (3 × SSC, 50 mM NaPO4, pH 7.4, 1 × Denhardt's solution, 0.25 mg/ml yeast tRNA, 10% dextran sulfate, 50% formamide, 10 mM dithiothreitol) was applied. The hybridization mix contained 50,000 dpm/ml of 35S-labeled RNA probes. The sections were coverslipped and incubated in moist chambers at 60C for 16 hr. The coverslips were then removed in 2 × SSC and the sections were subjected to the following posthybridization steps: RNase treatment (20 μg/ml RNase A and 1 U/ml RNase T1 in 10 mM Tris, pH 8.0, 0.5 M NaCl, 1 mM EDTA) for 60 min at 37C and successive washes in decreasing salt concentrations (2 ×, 1 ×, 0.5 ×, and 0.2 × SSC) for 10 min each, followed by incubation in 0.2 × SSC at 60C for 60 min. For autoradiography, slides were dipped in NTB-2 nuclear emulsion (Eastman Kodak; Rochester, NY) and developed after 3 weeks of exposure time. Developed sections were stained with hematoxylin and eosin, analyzed, and photographed in darkfield and brightfield modus with an AX 70 microscope (Olympus; Hamburg, Germany).
Confocal Laser Scanning Microscopy
Double-immunofluorescence detection was performed by covering the sections with a mixture of the two different primary antibodies in appropriate dilutions (Table 1) and by subsequent labeling with the species-specific secondary antibodies bearing the Alexa fluorochrome A647, A594, or A 488 (MoBiTec; Göttingen, Gemany) or the fluorochromes Cy2 or Cy3 (Dianova; Hamburg, Germany). Furthermore, streptavidin coupled with Alexa fluorochrome A647, A488, or A594 (MoBiTec) or with Cy2 or Cy3 was used in combination with biotinylated species-specific secondary antisera bearing the appropriate fluorochrome. For conventional double immunofluorescence, sections were analyzed and photographed with an AX 70 microscope (Olympus) equipped with the appropriate filter cubes for discriminating among the different fluorochromes. Confocal double-immunofluorescence analysis was performed with the Fluoview laser scanning microscope (Olympus). Digital confocal images were presented in false color.
Ethics
The procurement of human material during surgery was approved by the Ethics Committees of the Medical Faculties of the University of Marburg, the University of Kiel, and the University of Heidelberg. Oral informed consent was obtained from each patient before surgery. Monkey tissues were obtained in accordance with the approved animal care and use protocols of Bioqual (Rockville, MD).
Results
Expression of VMAT2 in Cutaneous Mast Cells and Cutaneous Mastocytosis
In all regions of the human and monkey skin, VMAT2 but not VMAT1 immunoreactivity was present in a considerable subpopulation of connective tissue immune cells and in varicose nerve fibers innervating the blood vessels (Figure 1). The VMAT2-positive cells were usually found in the dermis, with some enrichment around the superficial vascular plexus. They measured 8–15 μm in diameter, were round, oval, or fusiform, and possessed the morphological features of mast cells. Their number varied in the skin of different parts of the body, being most abundant in the scrotum and the prepuce of penis (data not shown). This pattern was seen in tissues fixed either in Bouin-Hollande or in 4% (para)formaldehyde with or without antigen retrieval.
Radioactive ISH revealed a corresponding expression pattern for VMAT2 mRNA in cells of the dermal layer. VMAT2-positive cells were identified as mast cells by poststaining with Giemsa and by double immunofluorescence, which revealed full co-positivity of VMAT2 and the mast cell marker tryptase in the granules of mast cells (Figure 2). All other (tryptase-negative) immune cells of the connective tissue were also negative for VMAT2, including lymphocytes and macrophages.
Tumor mast cells in patients with urticaria pigmentosa, solitary mastocytoma and diffuse cutaneous mastocytosis were fully co-positive for VMAT2, tryptase, and HDC (Figure 3). The specificity of the IHC staining was demonstrated by preabsorbing the VMAT2 antisera with 25 μmol of the recognizing C-terminal human VMAT2 peptide (Figure 3). In contrast, VMAT1 was absent from tumor mast cells in all cases of cutaneous mastocytosis (Figure 3).
Expression of VMAT2 in Cutaneous Langerhans Cells but Absence of VMATs in Langerhans Cell Histiocytosis
A considerable subpopulation of epidermal cells situated in the epidermis above the basal layer and within the pilosebaceous epithelium of human skin showed gene and protein expression of VMAT2 but not of VMAT1, as demonstrated by IHC and radioactive ISH (Figures 4 and 5). The VMAT2-immunoreactive cells exhibited dendritic features, with thin irregularly shaped cell processes (Figure 4). On the basis of their CD1a co-positivity, the VMAT2-positive cells were identified as epidermal Langerhans cells (Figure 6). In the epidermal layer, VMAT2 immunoreactivity was seen only in CD1a-positive cells. All other cell types of the epidermal layer were negative for VMAT2 and for VMAT1. High-resolution confocal laser scanning microscopy demonstrated three subcellular staining patterns of VMAT2 and CD1a. The cell surface was exclusively VMAT2-/CD1a+, some organelles in close association with the cellular surface were VMAT2+/CD1a+, and the majority of cytoplasmic granules were VMAT2+/CD1a– (Figure 6). In contrast to the normal skin, VMAT2 was absent from all tumor CD1a-positive cells in Langerhans cell histiocytosis (Figure 7).
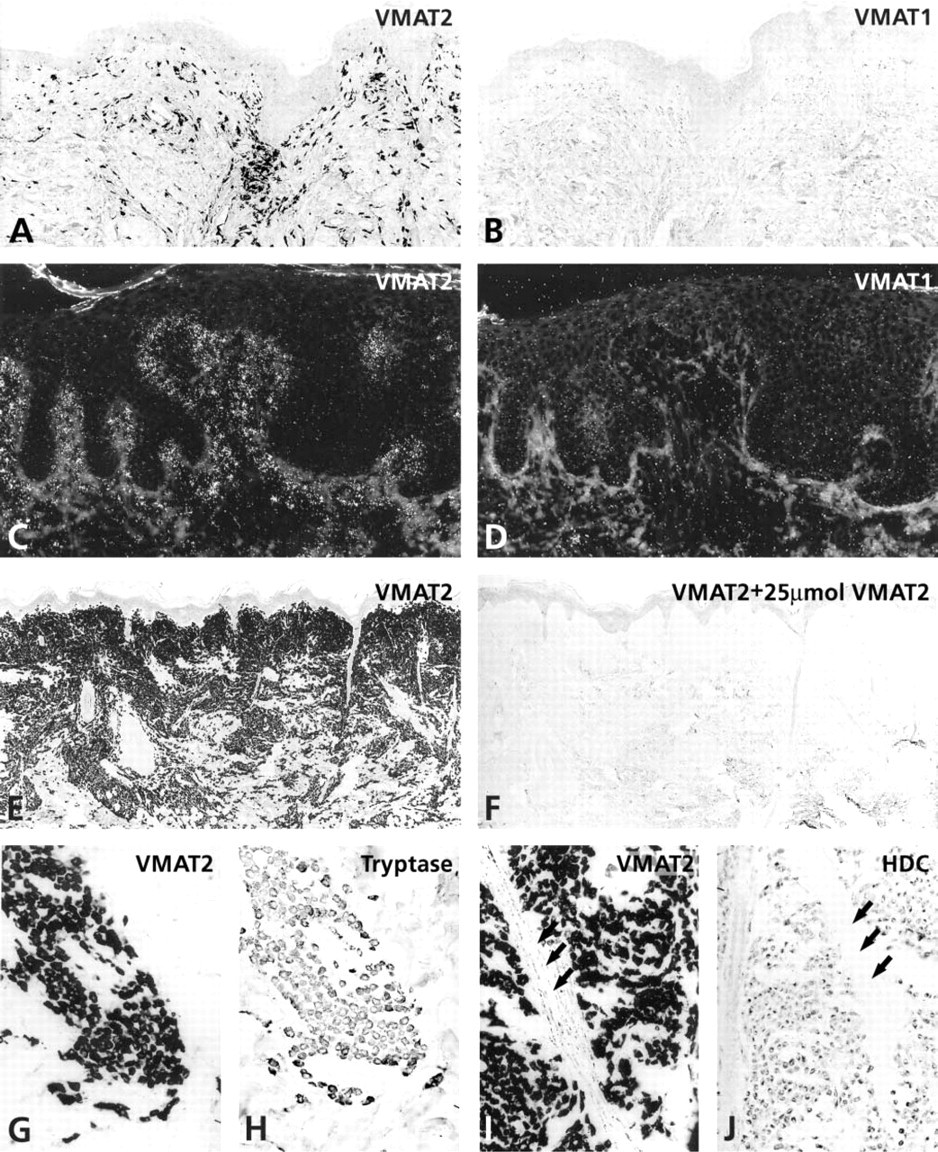
Differential expression of VMAT1 and VMAT2 in different types of cutaneous mastocytosis. (
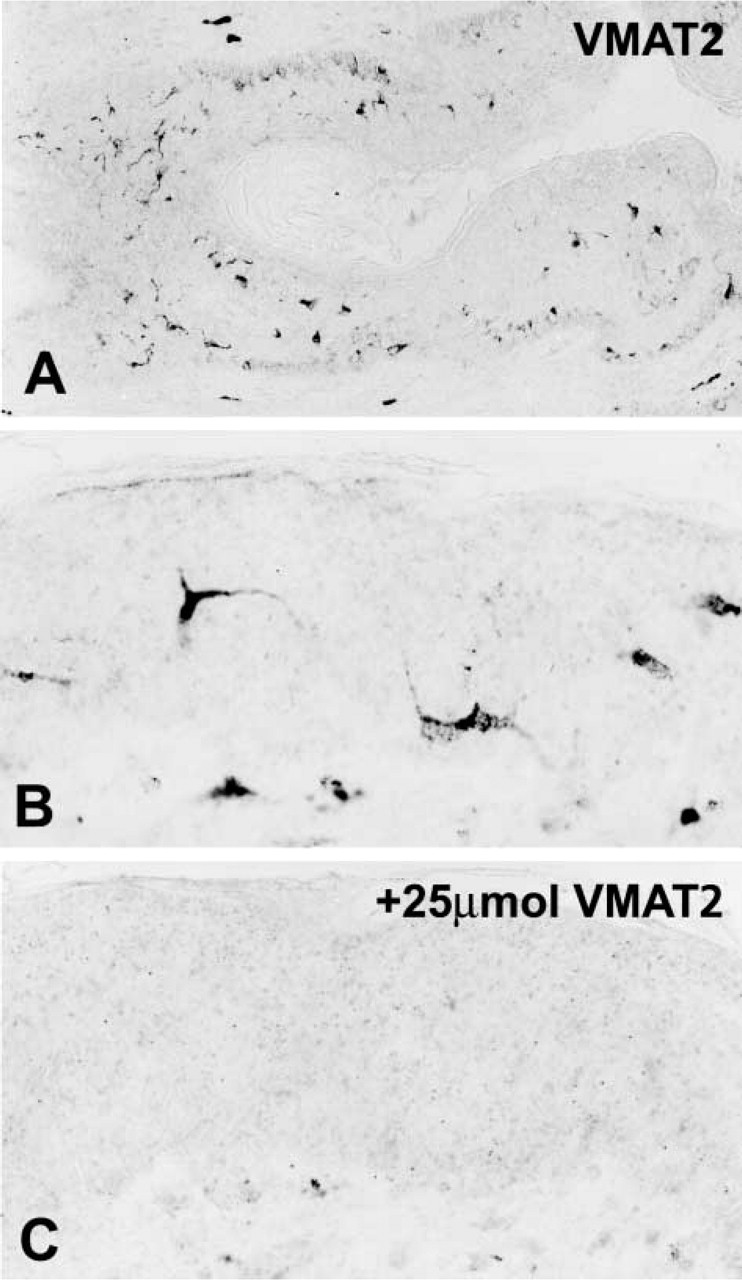
Immunohistochemistry for VMAT2 in the epidermal layer. (
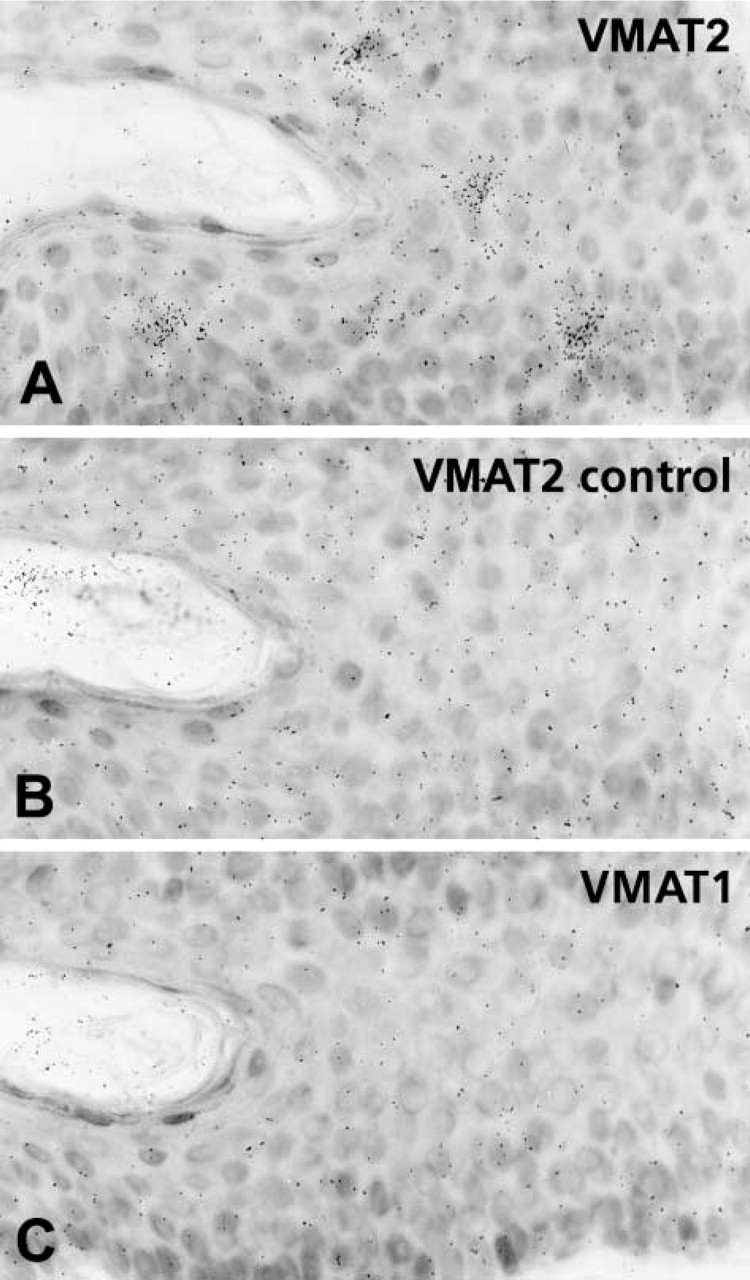
Radioactive in situ hybridization for VMAT2 in the epidermal layer. (
Discussion
Monoamines are stored in and secreted from immune cells. Storage of monoamines in secretory granules is mediated by either VMAT1 or VMAT2. Although VMAT2 was originally cloned from a rat basophilic leukemia cell line (Erickson et al. 1992), neither the expression and cellular localization of VMAT1 and VMAT2 in vivo nor their possible role as markers for immune cells in pathology has been examined. Our present study showed that the storage of monoamines in secretory granules of normal and tumor cutaneous mast cells is mediated by VMAT2. In addition, we found that CD1a-positive Langerhans cells of the human and monkey skin also express VMAT2. However, all cases of Langerhans cell histiocytosis were negative for VMAT2.
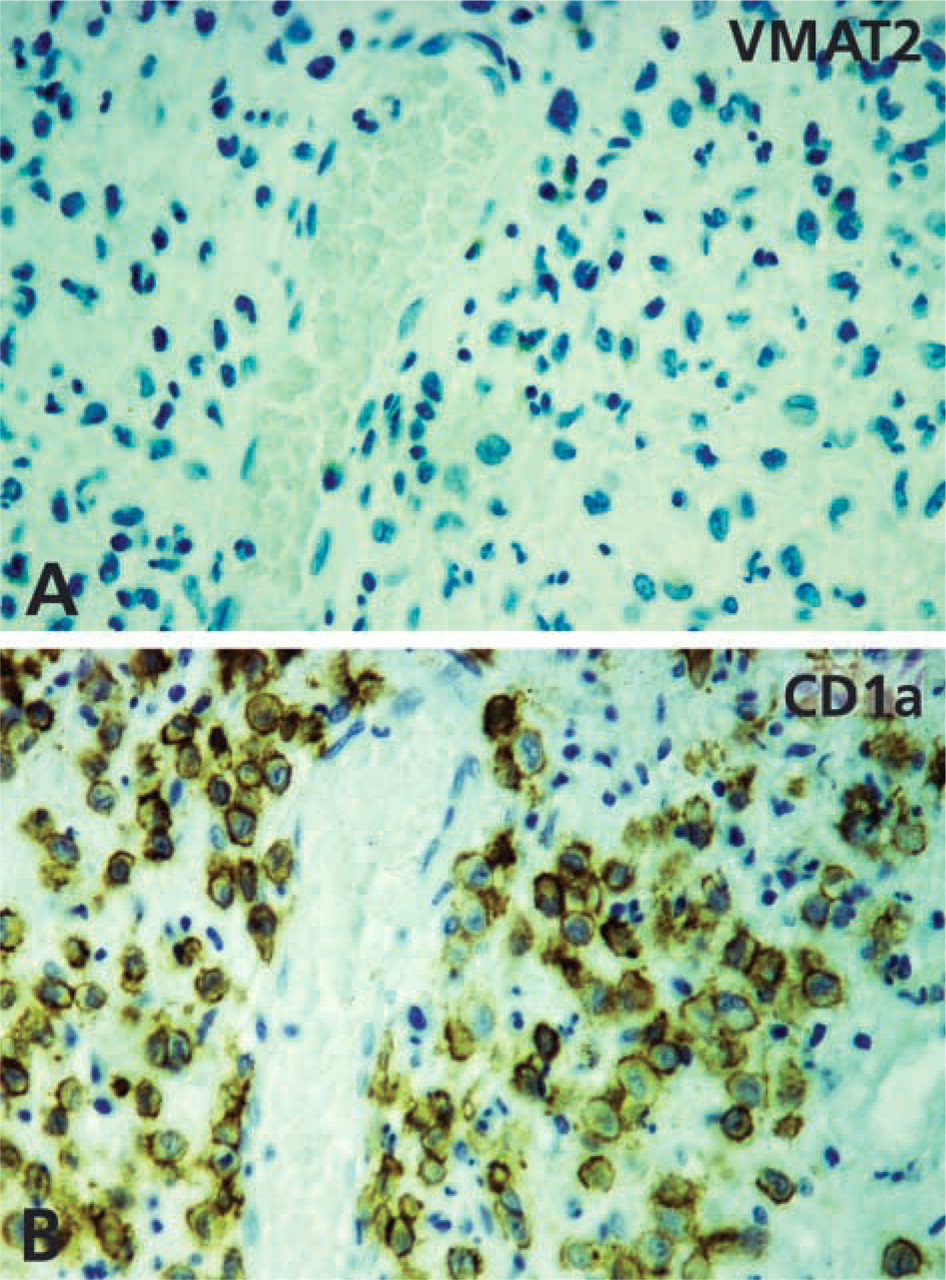
Absence of VMAT2 from CD1a-positive Langerhans cell histiocytosis. (
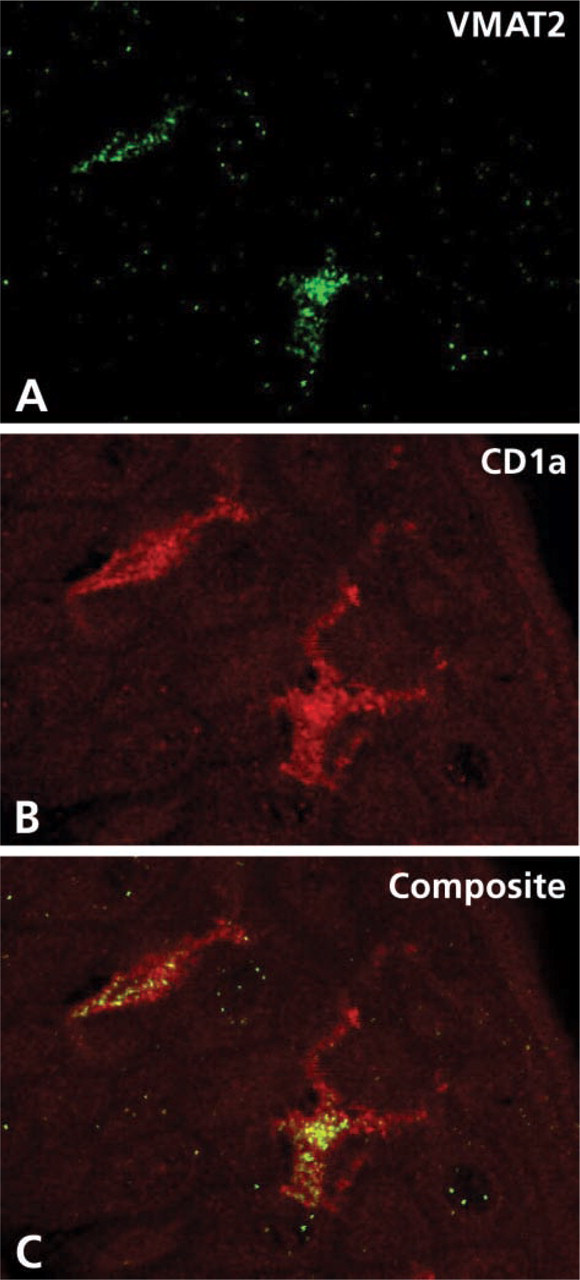
Identification of the cellular phenotype of VMAT2-immunoreactive immune cells of the epidermal layer and analysis of the subcellular distribution of VMAT2 shown by confocal laser scanning microscopy. (
Mast cells, which derive from CD34-positive bone marrow stem cells, produce and liberate a variety of biologically active agents, especially monoamines such as histamine. They appear to participate in a variety of inflammatory dermatoses and in some tumors (Atkins and Clark 1987; Melman 1987; Longley et al. 1995; Maurer et al. 1997; Tada et al. 2000). Corresponding to our finding that VMAT2 is present in all cutaneous mast cells of the primate skin, observations in knockout mice lacking the VMAT2 isoform have shown that the quantal release of histamine and serotonin from mast cells is mediated by VMAT2 (Travis et al. 2000). This finding, together with our results, suggests that VMAT2 does indeed play an active role in monoamine handling by cutaneous mast cells and, in addition, indicates that there is a developmental conservation of the VMAT2 molecule in mast cells between rodents and primates.
Recently, VMAT1 and VMAT2 were found to be excellent markers for the diagnosis of certain monoamine-handling endocrine tumors, such as tumors deriving from histamine-containing enterochromaffin-like cells and serotonin-containing enterochromaffin cells of the gastroenteropancreatic system (Eissele et al. 1999; Jakobsen et al. 2001; Anlauf et al. 2003a). We can extend this observation to various forms of cutaneous mastocytosis, including urticaria pigmentosa, solitary mastocytoma, and diffuse cutaneous mastocytosis. All these benign forms of mastocytosis revealed expression of VMAT2 but not of VMAT1. The diagnosis mastocytosis is traditionally based on the demonstration of focal accumulation of mast cells with typical histological and cytomorphological properties (Lennert and Parwaresch 1979; Horny and Valent 2001; Valent et al. 2001). However, depending on the organ system examined, type of mast cell disease, and presence of other diseases, it is sometimes difficult to diagnose mastocytosis histologically (Horny et al. 1985,1989,1992; Horny and Kaiserling 1987). Our study demonstrates that VMAT2 can be used as an excellent marker for cutaneous mastocytosis comparable to the well-established mast cell marker antibody against tryptase (Li et al. 1996; Horny et al. 1998). Because we found that VMAT2 was also expressed in mast cells of organ systems other than the skin (data not shown), it is likely that VMAT2 may also serve as a diagnostic marker for more aggressive forms of mastocytosis.
Surprisingly, VMAT2 but not VMAT1 was also expressed in a subpopulation of cells in the epidermal layer. According to their CD1a phenotype, these cells were identified as epidermal Langerhans cells (Klareskog et al. 1977; Krenacs et al. 1993; Moseley et al. 1997). Langerhans cells play an important role in the pathogenesis of contact dermatitis and in various other inflammatory dermatoses. They are thought to be specialized at processing and presenting antigens and to migrate to draining lymph nodes, where they initiate T-cell priming (Kripke et al. 1990; Taylor et al. 1991; Mommaas et al. 1992; Moll et al. 1993; Katou et al. 2000), albeit, a recent study suggests that a mechanism exists for priming virus-specific cytotoxic T-lymphocytes after skin infection that does not require antigen presentation by Langerhans cells (Allan et al. 2003).
The selective expression of VMAT2, which is responsible for the uptake and accumulation of biogenic amines to make them available for exocytotic release, adds a new important feature to the functional spectrum of this particular cell type. VMAT2 expression in epidermal Langerhans cells suggests that these cells constitutively accumulate, store, and release monoamines. The physiological action of monoamines such as catecholamines on skin immune function is well documented (Maestroni 2000; Seiffert et al. 2002; Kunzmann et al. 2003; Mazzoni et al. 2003). A few histochemical studies have demonstrated specific monoamine uptake in epidermal Langerhans cells (Axelsson et al. 1978; Sjoborg et al. 1978; Uno and Hanifin 1980; Crivellato et al. 1990; Falck et al. 1993). It is therefore tempting to speculate that Langerhans cells may, in addition to their function as antigen-presenting cells, modulate immune functions of the skin via regulated storage and secretion of monoamines to influence catecholamine acceptor bearing T-lymphocytes. Furthermore, monoamines secreted in a paracrine or autocrine manner may regulate Langerhans cell migration and antigen presentation (Maestroni 2000).
Analysis of the subcellular distribution of VMAT2 revealed a granular cytoplasmic pattern that was partially CD1a-positive. In contrast, the cell surface was exclusively CD1a-positive but VMAT2-negative. It has been shown that CD1a molecules traffic through the early recycling endosomal pathway (Salamero et al. 2001; McDermott et al. 2002). The endocytosis theory suggests that Birbeck granules originate from the cell membrane during receptor-mediated endocytosis as an elongation of CD1a-positive coated pits that pinch off to form intracellular Birbeck granules. Once formed, these structures could potentially deliver ligands to endosomal, prelysosomal, and lysosomal compartments (Ishii et al. 1984; Bartosik 1992). However, the function of Birbeck granules, which are so characteristic of Langerhans cells (Birbeck et al. 1961) remains enigmatic. On the basis of the endocytosis theory, it appears likely that the VMAT2-positive granules derived from CD1a-positive early endosomes form VMAT2-positive Birbeck granules as the intracellular compartment, able to accumulate monoamines. The hypothesis that Birbeck granules store monoamines via VMAT2 will require further support through detailed immunoelectron microscopic and functional studies.
Based on the expression of VMAT2 in cutaneous Langerhans cells, we anticipated that VMAT2 might also serve as a marker for the neoplastic counterparts of these cells in the skin. However, VMAT2 was absent from all cases of Langerhans cell histiocytosis examined. It has been demonstrated that Langerhans cell histiocytosis is a true neoplastic disease with clonal proliferation of theses cells (Willman et al. 1994). CD1a-positive Langerhans cells in histiocytosis may lose their ability to accumulate and secrete monoamines via VMAT2 during early tumorous transformation. Therefore, VMAT2 is not a useful marker for determining the cellular origin of Langerhans cell tumors. Whether the loss of VMAT2 phenotype in neoplastic Langerhans cells is causal to their tumorigenesis remains to be investigated. It is noteworthy that a loss of VMAT2 expression was also seen in a majority of tumors originating from pancreatic β-cells (Anlauf et al. 2003a).
In summary, the identification of VMAT2 in mast cells and Langerhans cells of the skin provides evidence of a molecular mechanism for monoamine storage and handling in these cells and provides a basis for imaging, as well as promoting and interfering with monoamine loading of these cells based on the known transport properties of human VMAT2 (Erickson et al. 1996). That monoamines may be critical for Langerhans cell function in host defense and inflammatory processes is now a subject that fully merits further investigation. The presence of VMAT2 in various forms of cutaneous mastocytosis indicates the importance of VMAT2 as a marker for normal and neoplastic mast cells and raises the important question of what monoamines might be stored in and secreted from human mast cells as a function of their intracellular concentrations and relative affinities for VMAT2.
Acknowledgments
We wish to thank Maike Pacena, Anja Paulus, Elke Rodenberg-Frank, Petra Sack, and Marion Zibuschka for their excellent technical assistance, and Heidemarie Schneider and Klaus Schoenheiz for photographic documentation. We are indebted to Katherine Dege for critically reading the manuscript. We thank Ivo Leuschner from the Institute for Paediatric Pathology, University of Kiel, for the generous gift of histiocytosis tissue specimens. The grant support of the Volkswagen-Stiftung (to EW, LEE) and of the DFG (to EW and MK-HS) is gratefully acknowledged.