
Abstract
Uptake of monoamines into secretory granules is mediated by the vesicular monoamine transporters VMAT1 and VMAT2. In this study, we analyzed their expression in inflammatory and hematopoietic cells and in patients suffering from systemic mastocytosis (SM) and chronic myelogenous leukemia (CML). Normal human and monkey tissue specimens and tissues from patients suffering from SM and CML were analyzed by means of immunohistochemistry, radioactive in situ hybridization, real time RT-PCR, double fluorescence confocal laser scanning microscopy, and immunoelectron microscopy. In normal tissue specimens, VMAT2, but not VMAT1, was expressed in mast cells, megakaryocytes, thrombocytes, basophil granulocytes, and cutaneous Langerhans cells. Further hematopoietic and lymphoid cells showed no expression of VMATs. VMAT2 was expressed in all types of SM, as indicated by coexpression with the mast cell marker tryptase. In CML, VMAT2 expression was retained in neoplastic megakaryocytes and basophil granulocytes. In conclusion, the identification of VMAT2 in mast cells, megakaryocytes, thrombocytes, basophil granulocytes, and cutaneous Langerhans cells provides evidence that these cells possess molecular mechanisms for monoamine storage and handling. VMAT2 identifies normal and neoplastic mast cells, megakaryocytes, and basophil granulocytes and may therefore become a valuable tool for the diagnosis of mastocytosis and malignant systemic diseases involving megakaryocytes and basophil granulocytes.
Keywords
M
VMAT1 and VMAT2 were found to be differentially expressed in monoamine-handling neurons of the central and peripheral nervous system (Weihe et al. 1994; Nirenberg et al. 1995; Peter et al. 1995; Schütz et al. 1998; Weihe and Eiden 2000; Anlauf et al. 2003b; Berkley et al. 2004), enterochromaffin- and enterochromaffin-like cells of the gastrointestinal tract (Weihe et al. 1994; Peter et al. 1995; Eissele et al. 1999; Weihe and Eiden 2000), and pancreatic beta cells (Anlauf et al. 2003a). A few studies have shown that VMAT1 and VMAT2 serve as excellent markers for monoaminehandling tumors originating from monoaminergic cells of the gastroenteropancreatic system (GEP), the adrenal medulla, and the paraganglia (Eissele et al. 1999; Rindi et al. 2000; Jakobsen et al. 2001; Anlauf et al. 2003a; Nilsson et al. 2004). In addition, specific radioligands have been developed and used for in vivo positron emission tomography to diagnose degenerative diseases of the central nervous system (CNS) and neoplastic monoamine-handling endocrine cells of the GEP (Kilbourn 1997; Efange 2000; Frey et al. 2001; Kolby et al. 2001, 2003; Fuente-Fernandez et al. 2003).
VMAT2 was originally cloned from a rat basophilic leukemia cell line (Erickson et al. 1992). This suggested that monoamine storage in this cell type is mediated by VMAT2. However, neither the in vivo expression of VMAT1 and VMAT2 in cells of the hematopoietic and lymphoid system nor their possible role as markers for myeloproliferative disorders has been examined in detail. Recently, we found VMAT2 to be expressed in mast cells and cutaneous Langerhans cells of the skin (Anlauf et al. 2004). Our present study addressed three main issues: (1) do the extradermal mast cell also express VMAT2, (2) are there other cells of the bone marrow that are positive for VMAT1 or VMAT2, and (3) are VMATs possible diagnostic tools for mastocytosis and for chronic myelogenous leukemia (CML), as representatives of myeloproliferative disorders? To answer these questions, we analyzed a series of different normal human and monkey tissues and specimens from patients suffering from mastocytosis and CML.
Materials and Methods
Normal Tissue Specimens
Normal human tissue samples from various skin regions (n = 10); the salivary glands; gastrointestinal tract and greater omentum (n = 15); pancreas (n = 5); liver, gallbladder, and bile duct system (n = 10); spleen (n = 5); lymph nodes (n = 10); tonsils (n = 10); thymus (n = 3); bone marrow (n = 20); nose, trachea, and lung (n = 7); kidney and ureter (n = 5); testis and prostate (n = 5); uterus, tube, and ovaries (n = 5); heart and large blood vessels (n = 5); thyroid gland and pituitary (n = 5); various regions of the central nervous system (n = 5); autonomous and spinal cord ganglia (n = 5); and peripheral nerves (n = 3) were used to analyze the expression of VMAT1 and VMAT2. Tissue samples similar in anatomic localization to those of humans were obtained from five rhesus monkeys, as described previously (Rausch et al. 1994). In addition, representative blood and bone marrow smears and buffy coat–derived pooled platelets concentrates were analyzed. As positive control for VMAT1 and VMAT2, adrenal gland tissues from two rhesus monkeys and from two patients with unilateral adrenalectomy, one suffering from pheochromocytoma (m/63) and one from Conn's syndrome (f/70), were used.
For immunohistochemical analysis, the human tissues were fixed in either Bouin-Hollande fixative, 10% formalin, or 4% formaldehyde/PBS for 24–48 hr and then embedded in paraffin. The monkey tissues were perfused with 4% formaldehyde/PBS before postfixation in Bouin-Hollande for 24–48 hr, as described previously (Rausch et al. 1994). After dehydration in a graded series of 2-propanol solution, the tissues were embedded in Paraplast Plus (Merck; Darmstadt, Germany). Adjacent sections (3 μm or 5 μm thick) were cut and deparaffinized. Antigen retrieval to increase the sensitivity of immunodetection was performed by heating the sections at 92–95C for 15 min in 0.01 M citrate buffer (pH 6) according to the DAKO protocol (Hamburg, Germany). Nonspecific binding sites were blocked with 5% BSA (Serva; Heidelberg, Germany) in PBS (pH 7.45) followed by an avidin-biotin blocking step (avidin-biotin blocking kit; Boehringer Ingelheim, Germany).
For in situ hybridization, human and monkey tissue specimens were immediately frozen in dry ice and stored at −80C. Cryosections (12–16 μm thick) were placed on presilanized glass slides, fixed in 4% phosphate-buffered formaldehyde for 60 min, followed by three 10-min washes in 50 mM PBS (pH 7.4). The slides were then briefly rinsed in distilled water, incubated in 0.1 M triethanolamine (pH 8.0) for 1 min and for 10 min in the same solution containing 0.25% v/v acetic anhydride under rapid stirring. They were then quickly rinsed in 2X SSC, dehydrated in 50% and 70% ethanol, and air dried.
Immunohistochemistry
Rabbit antiserum VMAT1/10 raised against the C-terminal sequence of human VMAT1 and antiserum no. 80182 raised against the C-terminal sequence of human VMAT2 were employed for the immunohistochemical investigations (Table 1). Poststaining with Giemsa and the antiserum against tryptase were used to identify mast cells of all organs (Horny et al. 1998). The CD25 antibody was used to discriminate between normal and neoplastic mast cells in patients with systemic mastocytosis (Sotlar et al. 2004). Further, previously very well characterized antibodies were used to analyze the cell-specific expression of VMATs in bone marrow, lymph node, tonsil, and thymus as summarized in Table 1.
The tissue sections were incubated with the primary antibodies overnight at 18C (diluted as shown in Table 1) followed by a 2 hr incubation at 37C. The sections were then washed in distilled water and in 50 mM PBS and incubated with species-specific biotinylated secondary antibodies (Dianova; Hamburg, Germany) for 45 min at 37C, washed several times and incubated for 30 min with the ABC reagents (Vectastatin Elite ABC kit; Boehringer Ingelheim). Immunoreactions with polyclonal antibodies were visualized with 3′3-diaminobenzidine or enhanced by the addition of 0.08% ammonium nickel sulfate, as previously described (Anlauf et al. 2003a). Monoclonal antibodies were visualized by using an alkaline phosphatase system, as described previously (Kreipe et al. 1993).
List of primary antibodies
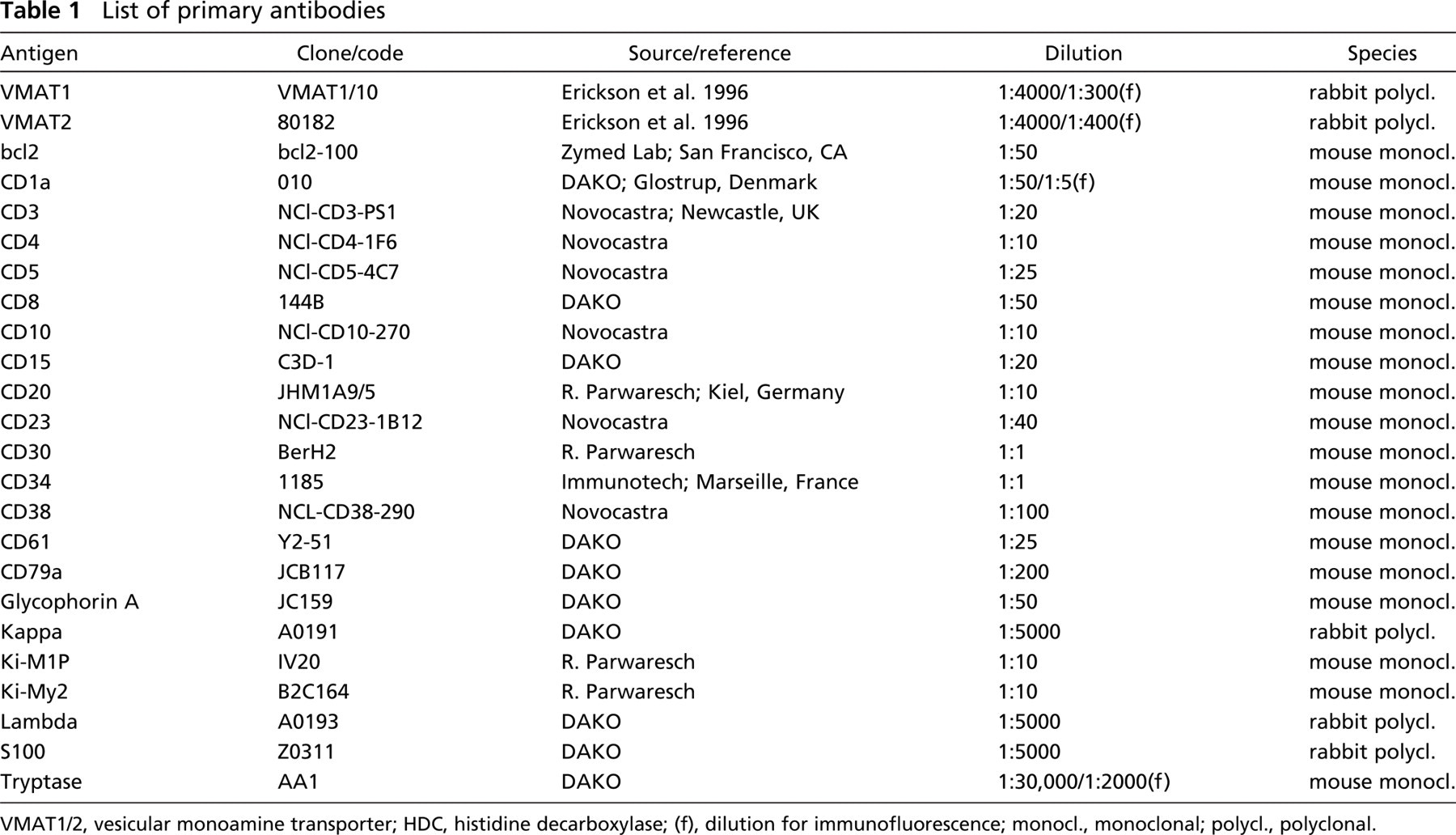
VMAT1/2, vesicular monoamine transporter; HDC, histidine decarboxylase; (f), dilution for immunofluorescence; monocl., monoclonal; polycl., polyclonal.
The specificity of the immunohistochemical detection of VMAT1 and VMAT2 was analyzed by preabsorbing the antisera with 25 μM of the C-terminal human VMAT1 and VMAT2 peptide, respectively.
Generation of 35S-labeled Riboprobes
To generate specific radioactive cRNA probes for the detection of VMAT1 mRNA, a 247-bp long DNA restriction fragment of hVMAT1 cDNA corresponding to nt. 436–682 was subcloned into Bluescript II KS+ (Stratagene; Heidelberg, Germany) (Erickson et al. 1996). For VMAT2 mRNA detection, a 269-bp long DNA fragment of hVMAT2 corresponding to nt. 244–512 was subcloned into pCDNAI (Invitrogen; Leek, The Netherlands) (Erickson and Eiden 1993). The inserted sequences were verified by double-stranded DNA sequencing. In vitro transcription of the two vector constructs using 35S-UTP-labeled nucleotide yielded anti-sense riboprobes for hVMAT1 after linearization with XhoI and incubation with T7 RNA polymerase, and for hVMAT2 after linearization with XbaI and incubation with SP6 RNA polymerase. To increase the tissue penetration of the probes, the generated transcripts were reduced to ∼200 nucleotide fragments by limited alkaline hydrolysis, as described previously (Angerer et al. 1987).
Radioactive In Situ Hybridization
To each section hybridization buffer (3X SSC, 50 mM NaPO4, pH 7.4, 1X Denhardt's solution, 0.25 mg/ml yeast tRNA, 10% dextran sulfate, 50% formamide, 10 mM dithiothreitol) was applied. The hybridization mix contained 50,000 dpm/ml of 35S-labeled RNA probes. The sections were cover slipped and incubated in moist chambers at 60C for 16 hr. The cover slips were then removed in 2X SSC and the sections were subjected to the following post-hybridization steps: RNase treatment (20 μg/ml RNase A and 1 U/ml RNase T1 in 10 mM Tris, pH 8.0, 0.5 M NaCl, 1 mM EDTA) for 60 min at 37C and successive washes in decreasing salt concentrations (2X, 1X, 0.5X, and 0.2X SSC) for 10 min each, followed by incubation in 0.2X SSC at 60C for 60 min.
For autoradiography, the slides were dipped in NTB-2 nuclear emulsion (Eastman Kodak; Rochester, NY) and developed after 3 weeks of exposure time. The developed sections were stained with hematoxylin and eosin, analyzed, and photographed in dark-field and bright-field modus with an AX 70 microscope (Olympus; Hamburg, Germany).
RNA Extraction and Real-time RT-PCR
From each cryo-protected normal human tissue specimen, 10 sections (10-μm thick) were cut and the first section was stained with hematoxylin and eosin. The other sections were used for dissection using a 24-gauge needle or a razor blade. Total RNA was extracted using Trizol according to the manufacturer's recommendations (Invitrogen; Karlsruhe, Germany). Equal amounts of RNA from each sample were reverse transcribed using random hexamer primers (Amersham Biosciences; Freiburg, Germany), and the cDNAs were used as templates in subsequent quantitative PCR reactions. Real-time PCR was performed on an ABI Prism 7000 Sequence Detection System using commercially available specific primers and probes for the target, VMAT2, and the endogenous control, eukaryotic 18s rRNA (assays-on demand Hs00161858_m1, and Hs99999901_s1, respectively), and reagents as recommended by the manufacturer (all Applied Biosciences; Darmstadt, Germany). The relative levels of gene expression were calculated using the ΔΔCt method as described by Cho et al. (2003).
Confocal Laser Scanning Microscopy
Double immunofluorescence was performed by covering the sections with a mixture of the two different primary antibodies in appropriate working dilutions (Table 1) and by subsequent labeling with the species-specific secondary antibodies coupled to the Alexa fluorochromes A647, A594, or A488 (MoBiTec; Göttingen, Germany) or the fluorochromes Cy2 or Cy3 (Dianova). Streptavidin coupled to Alexa fluorochromes A647, A488, or A594 (MoBiTec) or to Cy2 or to Cy3 was used in combination with biotinylated species-specific secondary antisera bearing the appropriate fluorochrome. For conventional double immunofluorescence, the sections were analyzed and photographed with an AX 70 microscope (Olympus) equipped with the appropriate filter cubes for discriminating between the different fluorochromes. Confocal double immunofluorescence analysis was performed with a Fluoview laser scanning microscope (Olympus). Digital confocal images were presented in false color.
Immunoelectron Microscopy
Ultrathin sections were incubated overnight at room tempperature with the polyclonal antibody against VMAT2 (1:1000) as previously described (Anlauf et al. 2000). After six 5-min rinses in PBS, pH 7.6, anti-rabbit goat IgG conjugated with 10 nm gold particles (Dianova) was applied at a dilution of 1:30. After a further incubation period of 90 min, the sections were rinsed with distilled water and contrasted with uranyl acetate and lead citrate (Weihe et al. 1991). Controls were performed by preabsorbing the antisera with 25 μM of the C-terminal human VMAT2 peptide, respectively (Weihe et al. 1991). The sections were analyzed with a Philipps EM 400 electron microscope (Eindhoven, the Netherlands).
Tissue from Patients with Mastocytosis or CML
Formalin-fixed tissue specimens from 31 patients suffering from systemic mastocytosis were retrieved from the consultation files of the Department of Pathology of the University of Lübeck and the Department of Hematopathology of the University of Kiel. In all of the patients with systemic mastocytosis, the neoplastic character of the mast cells had been confirmed on the basis of their CD25 expression, elevated tryptase serum levels, and evident c-kit mutation D816V (Sotlar et al. 2004). The diagnoses followed the World Health Organization classification system for mastocytosis (Valent et al. 2001b): indolent systemic mastocytosis (n = 11), systemic mastocytosis with associated clonal hematological nonmast cell lineage disease (n = 11), aggressive systemic mastocytosis (n = 6), mast cell leukemia (n = 2), and mast cell sarcoma (n = 1). In nine additional patients with typical clinical signs of cutaneous mastocytosis skin and bone marrow, specimens were analyzed prospectively to confirm the clinical diagnosis and to determine whether or not they showed systemic involvement. To study the expression of VMAT1 and VMAT2 in CML, bone marrow biopsies and smears from 15 patients diagnosed according to the World Health Organization criteria were obtained (Vardiman et al. 2001).
Ethics
The procurement of human material during surgery was approved by the Ethics Committees of the Medical Faculties of the University of Marburg and the Universities of Kiel and Lübeck. Oral informed consent was obtained from each patient before surgery. Monkey tissues were obtained in accordance with the approved animal care and use protocols of Bioqual, Inc. (Rockville, MD).
Results
Analysis of VMAT2 Protein and mRNA Expression in Normal Tissue Specimens
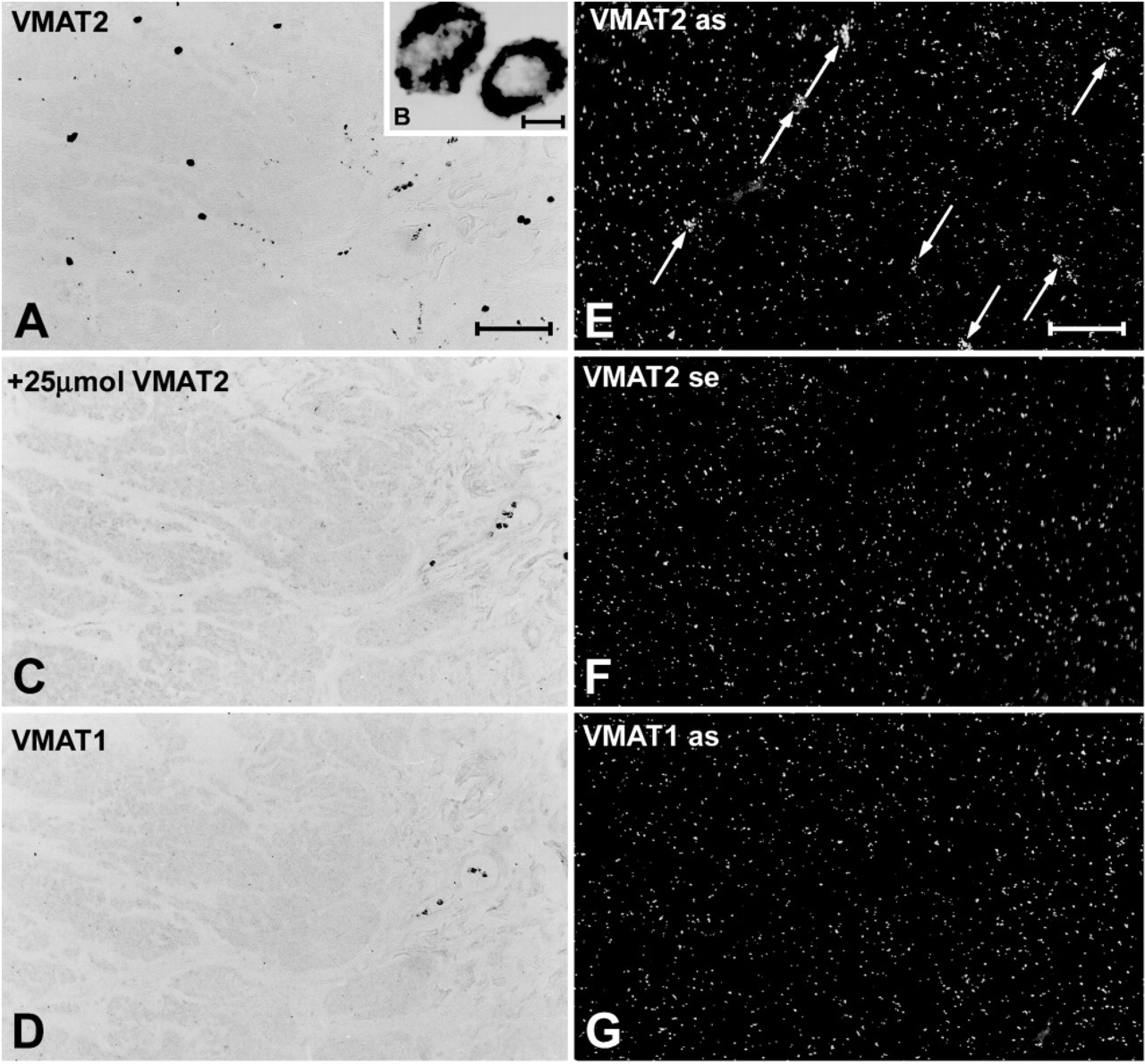
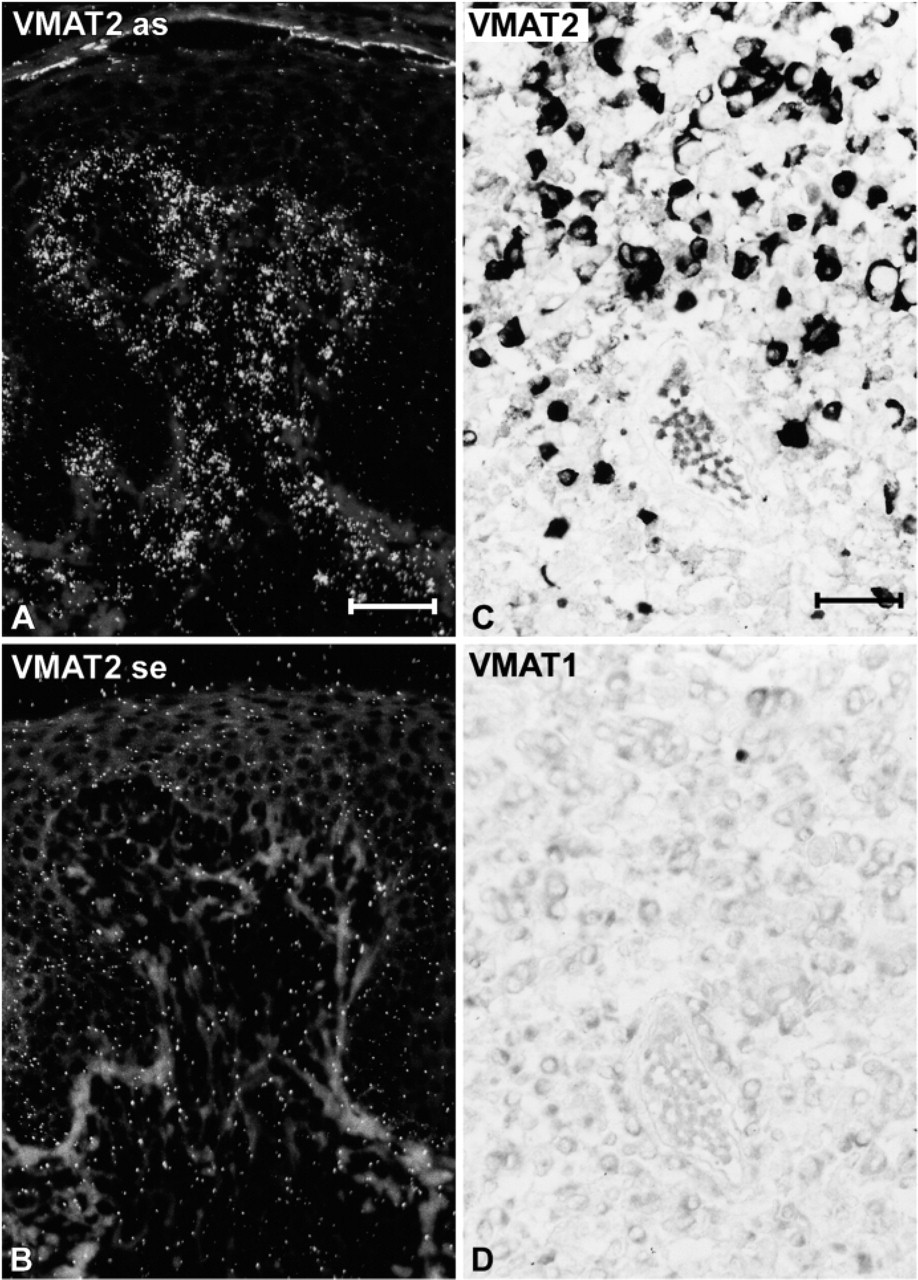
Immunohistochemistry and radioactive in situ hybridization revealed an identical distribution pattern for VMAT2 protein and mRNA in a sizeable subpopulation of connective tissue- and mucosa-associated inflammatory cells (Figure 1 and Figure 2). In paraffin-embedded sections, the VMAT2-positive cells measured 8–15 μm in diameter, were round, oval, or fusiform in shape and revealed granular cytoplasmic VMAT2 immunoreactivity, suggesting that these cells are mast cells (Figure 1).
Real time RT-PCR analysis revealed VMAT2 mRNA in all tissues observed. It was most abundant in the skin, spleen, lung, and the uterine cervix. Lower levels were found in the esophagus, the large and small intestine, and the lymph nodes (Figure 3).
The results of immunohistochemistry and in situ hybridization corresponded to each other. The highest density of VMAT2-positive cells was seen in the dermis, the lung, the tonsil, and the spleen. In the dermal layer of the skin, the number of VMAT2-positive cells varied in different parts of the body, being most abundant in the scrotum and the prepuce of the penis. VMAT2 was also expressed in cutaneous Langerhans cells, as previously reported by our group (Anlauf et al. 2004) (data not shown).
All parts of the gut contained a sizeable subpopulation of VMAT2-positive cells in the lamina propria mucosae and in the connective tissue of the outer layers and the omenta (Figure 1, Figure 2, and Figure 4). Scattered VMAT2-positive cells were seen along the ducts of the salivary glands and the pancreas and within the connective tissue of the bile duct system, but they were sparse in the acinar parenchyma. There were small numbers of VMAT2-positive cells in the portal tracts and sinusoids of the liver (data not shown).
Expression of VMAT2 in the connective tissue of the gut (
In lymphoid organs and the bone marrow, the highest densities of VMAT2-positive cells were seen in the peritonsillar connective tissue and in close association with fibrous septa of the tonsil and the spleen. The connective tissue of the thymus and the lymph nodes revealed scattered VMAT2-positive cells, but lymphocytes were VMAT2-negative (Figure 4). In the bone marrow all megakaryocytes were VMAT2 positive (Figure 4D). In addition, a minor subpopulation of less than 0.5% of the hematopoietic cells, mainly localized around the small blood vessels, were VMAT2-positive (Figure 4D).
High densities of VMAT2-positive cells were seen in all parts of the respiratory system. In the nose, the trachea, and the bronchi, some of these cells had migrated into the respiratory epithelium, whereas in the lung parenchyma, most VMAT2-positive cells were localized within the connective tissue of the alveolar septa (Figure 4).
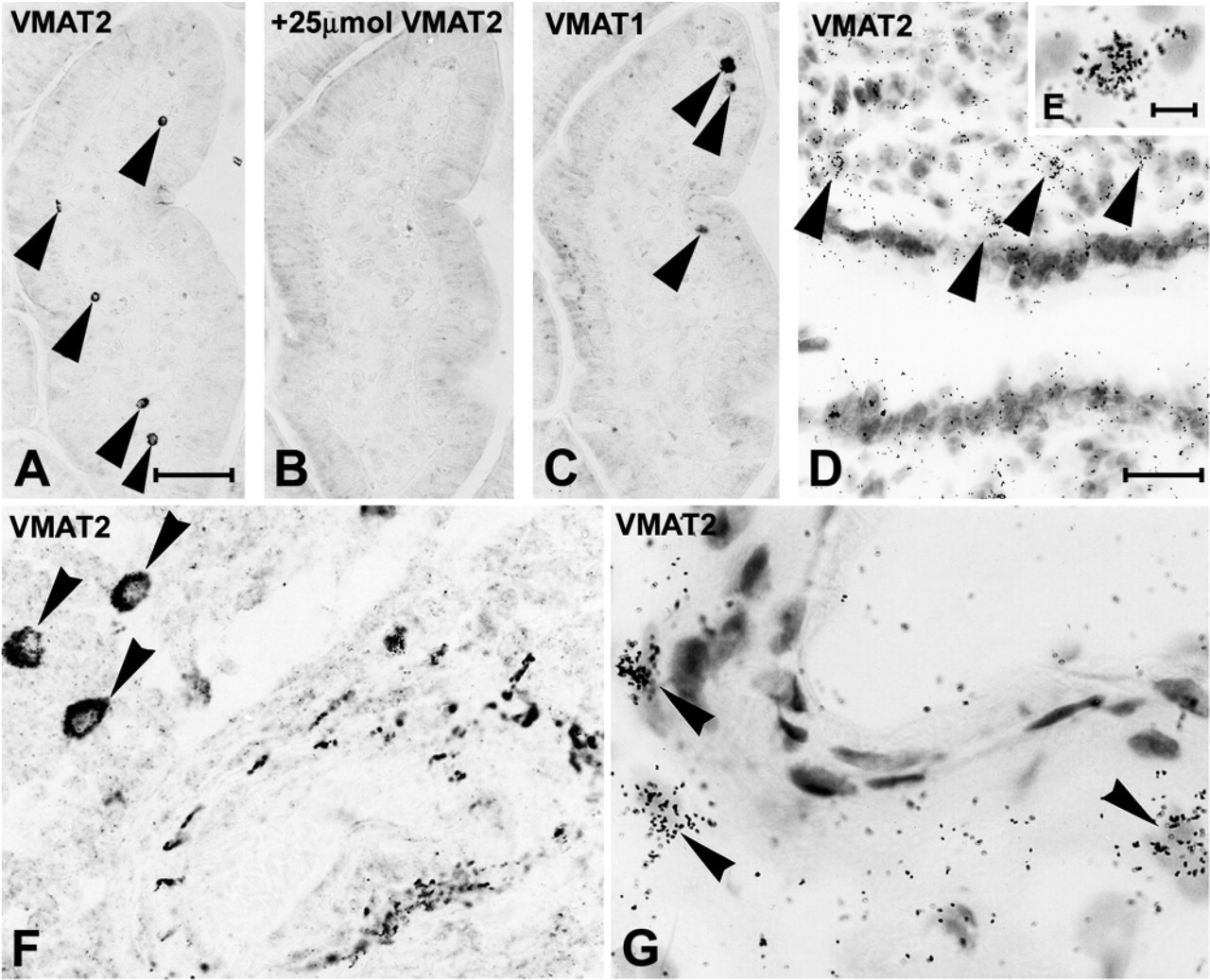
Expression of VMAT and VMAT2 in the mucosa and submucosa of the gut (
In the nervous system, VMAT2-positive cells were extremely sparse. They were found in close association with nerve cells of the autonomic ganglia and peripheral nerves and close to blood vessels of the CNS, with the highest densities being seen within the leptomeninges. In all other organs (heart and large blood vessels, urogenital tract, and endocrine organs), VMAT2-positive cells were sparse. Most of them were localized within the connective tissue (data not shown).
Identification of the Cellular Phenotype of VMAT2-positive Cells
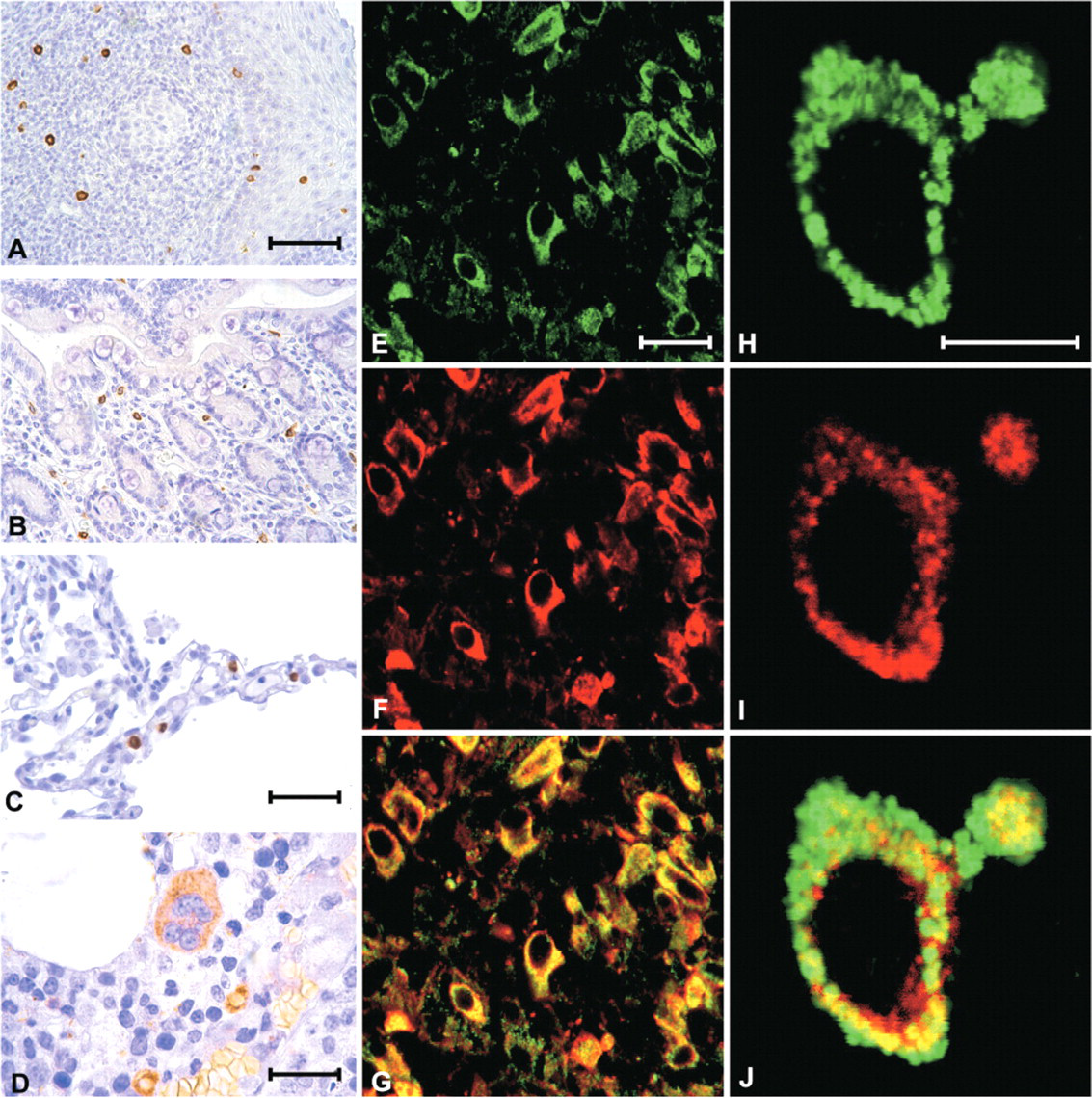
In all lymphoid organs and in the lamina propria mucosae and the connective tissue of other organ systems, VMAT2 was expressed in mast cells, as demonstrated by confocal laser scanning double fluorescence microscopy, which revealed full coexpression of VMAT2 and tryptase (Figures 4E–4G). In contrast, VMAT1 was absent from inflammatory cells, but was expressed in some neuroendocrine cells of the epithelial layers. High-resolution confocal laser scanning microscopy and immunoelectron microscopy showed that VMAT2 was localized at the membrane of the secretory granules of mast cells (Figures 4H–4J and Figure 5). Other inflammatory cells, including various maturation stages of B-lymphocytes and T-lymphocytes (CD 3, 4, 5, 8, 10, 15, 20, 23, 30, 79a, and bcl2) and plasma cells (kappa, lambda, CD38, and CD79a), were negative for VMAT2. With the exception of CD1a-positive Langerhans cells of the epidermal layer, macrophages and dendritic cells of all lymphoid organs did not reveal any immunoreactivity for VMAT1 and VMAT2 (data not shown).
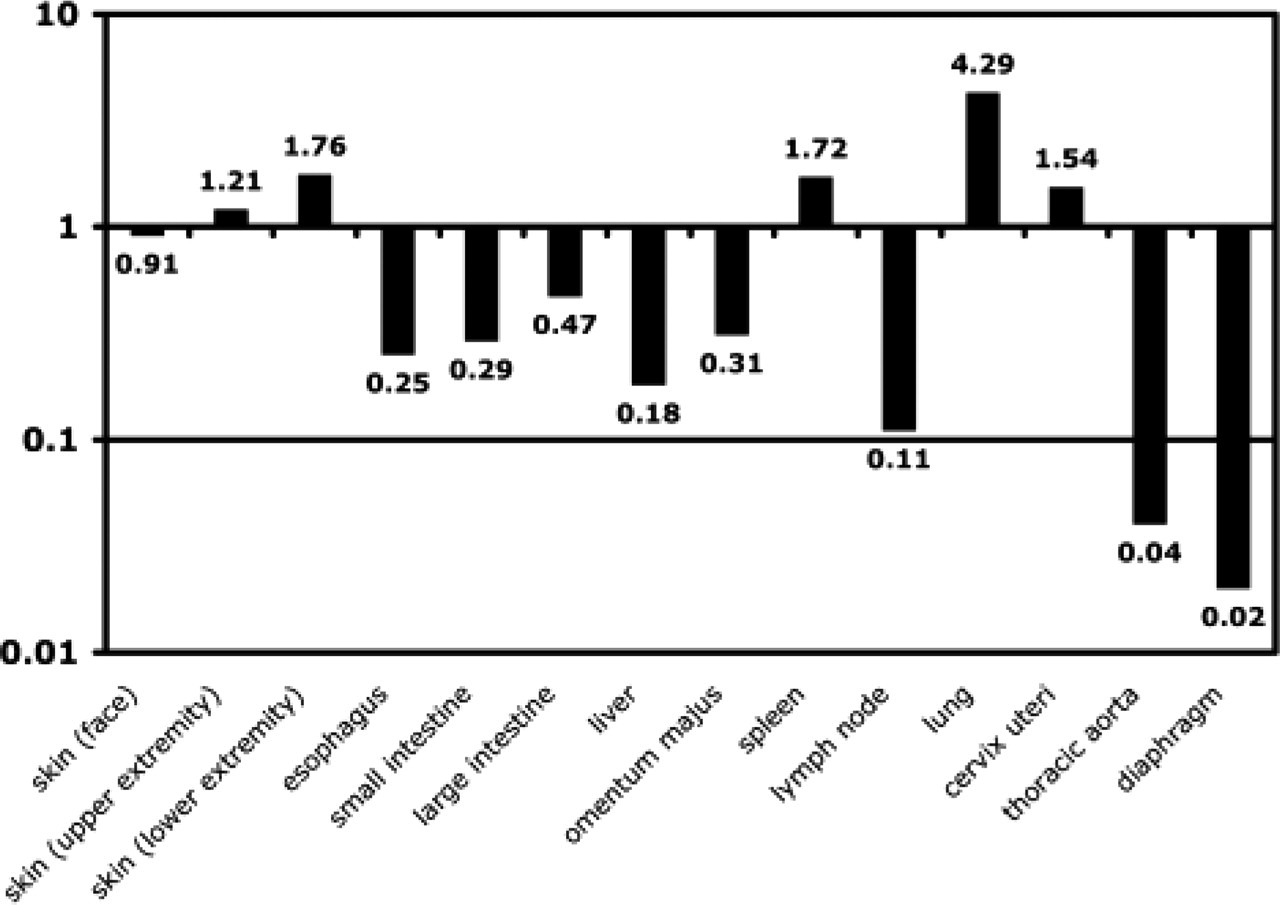
Detection of VMAT2 mRNA in various tissue specimens by real-time RT-PCR. The diagram shows the difference in VMAT2 expression in the respective tissues in comparison with a skin specimen from the trunk, which was defined as standard and has a relative value of 1. Significant amounts of VMAT2 mRNA were detected in all tissues investigated, with the expression level ranging from 0.02 (diaphragm) to 4.29 (lung).
In the hematopoietic system, VMAT2 was expressed in CD61-positive megakaryocytes and in thrombocytes (Figure 4D and Figure 6). Further VMAT2 expressing cells of the bone marrow comprised less than 0.5% hematopoietic cells and were identified as mast cells (VMAT2+/tryptase+) and basophil granulocytes (VMAT2+/tryptase−). Other hematopoietic cells—CD34-positive cells, erythropoietic cells (glycophorin A), granulopoietic precursor cells (ki-My2), and eosinophil granulocytes, which were identified by their typical granules—were negative for VMAT1 and VMAT2 (data not shown).
Expression of VMAT2 in Patients with Mastocytosis and CML
VMAT2 but not VMAT1 was found to be expressed in all patients with cutaneous and systemic mastocytosis, irrespective of the infiltration pattern, subtype of systemic mastocytosis and granule content (Figure 7 and Figure 8). The number of VMAT2-positive mast cells almost equaled that of tryptase-immunoreactive cells (Figure 8). The immunohistochemical detection of VMAT2 and tryptase was of equal diagnostic relevance, with the exception of two aggressive systemic mastocytosis cases that revealed reduced immunoreactivity for tryptase but strong VMAT2 expression (Figures 8D–8G).
In bone marrow specimens from patients suffering from CML, VMAT2 expression was seen in all megakaryocytes, in particular in cases with extensive proliferation of small hypolobulated megakaryocytes. There was a slight increase in mast cells (VMAT2+/tryptase+) in most specimens. Basophilia (VMAT2+/tryptase−) identified in blood and bone marrow smears was present in all cases. VMAT2 was absent from all other neoplastic cell populations (e.g., neutrophils and eosinophils, and erythropoietic cells) (data not shown).
Discussion
In a recent study, we showed that the expression of VMAT2 characterizes cutaneous mast cells and epidermal Langerhans cells (Anlauf et al. 2004). We can extend this observation by showing that VMAT2 is expressed in mast cells from all human and monkey organs. In addition, VMAT2 but not VMAT1 was found to be expressed in megakaryocytes, thrombocytes, and basophil granulocytes. Both VMAT2 and VMAT1 were absent from lymphocytes, plasma cells, neutrophil and eosinophil granulocytes, erythropoietic cells, and, with the exception of cutaneous Langerhans cells, also from dendritic cells and macrophages. VMAT2 is expressed in all mast cells, megakaryocytes, and basophil granulocytes, regardless of whether they are derived from patients or healthy controls.
It is well known that mast cells, which derive from CD34-positive bone marrow stem cells, store monoamines. Mediators of mast cells stored and released in granules (e.g., histamine) function as amplifiers of the inflammatory response (Kaliner 1979; Serafin and Austen 1987; Kunzmann et al. 2003). Among the differential immunohistochemical markers for mast cells, tryptase is the most sensitive and ubiquitous (Horny et al. 1998). Our results showed, on the basis of its co-expression with tryptase, that VMAT2 is expressed in mast cells of all human and monkey organs. The immunohistochemical distribution pattern of VMAT2 matched the results of the radioactive in situ hybridization and RT-PCR. By using high-resolution confocal laser scanning microscopy and immunoelectron microscopy, we were able to localize VMAT2 to the membrane of secretory granules in mast cells. These observations, together with the previous pharmacological finding that VMAT2 knockout mice lack the capacity to release histamine and serotonin from mast cells (Travis et al. 2000), further supports the view that VMAT2 plays an active role in monoamine handling in mast cells of rodents and primates.
Expression of VMAT2 in various lymphoid tissues and the bone marrow (
The expression of VMAT2 by megakaryocytes and basophil granulocytes indicates sequestration of monoamines in these cells. In contrast, absence of VMAT1 and VMAT2 from other cells of the hematopoietic and lymphoid lineage (e.g., eosinophil and neutrophil granulocytes, erythropoietic cells, lymphocytes and plasma cells) shows that vesicular monoamine storage mechanisms are restricted to megakaryocytes and basophil granulocytes. Though it has been suggested that VMATs can be transcriptionally activated in a preB-cell line and may be present in peripheral blood lymphocytes (Watson et al. 1999; Amenta et al. 2001), we could not confirm these observations in situ. Although there is strong evidence in the literature that lymphocytes synthesize catecholamines (Bergquist et al. 1994; Josefsson et al. 1996; Musso et al. 1996), our results suggest that storage of monoamines in lymphocytes is not dependent on VMATs. On the other hand, it is well established that basophil granulocytes store histamine and megakaryocytes accumulate serotonin (Dvorak et al. 1995; Dvorak et al. 1996; Holtje et al. 2003). Our results showed that VMAT2 is responsible for the sequestration of monoamines in these cell types. The reported absence of VMAT2 from macrophages other than cutaneous Langerhans cells implies that macrophages obtain a monoamine-handling phenotype specifically when they have entered the epidermis. Langerhans cells seem to lose their VMAT2 phenotype during their migration to lymphoid organs.
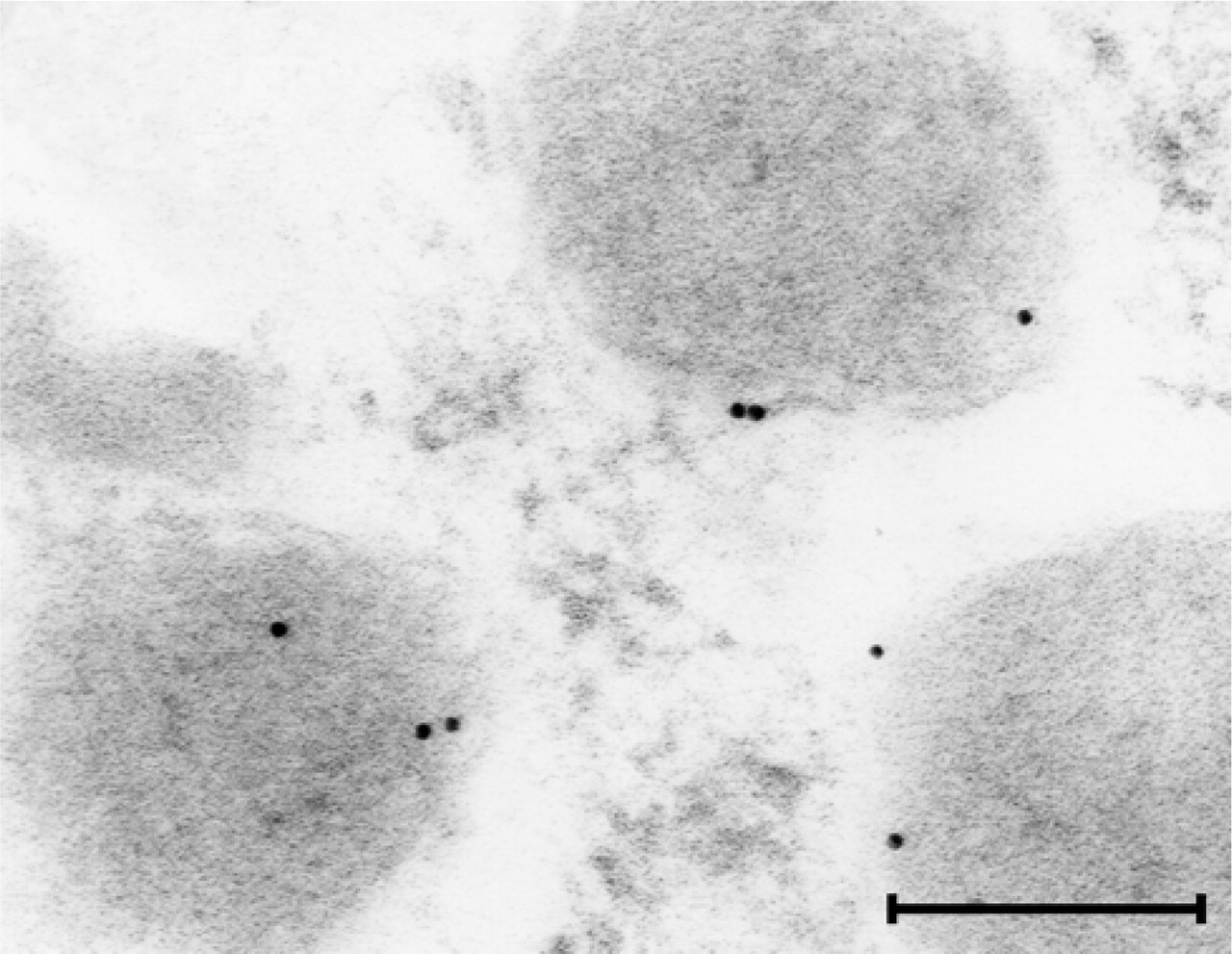
Immunoelectron microscopic localization of VMAT2 protein to secretory granules of a cutaneous mast cell. Gold particles representing the VMAT2 immunogold precipitate are closely associated with the membrane of secretory granules within a mast cell. Bar = 150 nm.
Recently, VMATs were found to be excellent tools for the pathological and scintigraphical diagnosis of degenerative diseases of the CNS and certain neoplastic monoamine-handling endocrine cells of the GEP (Eissele et al. 1999; Jakobsen et al. 2001; Kolby et al. 2001, 2003; Nilsson et al. 2004). Based on the observation that neoplastic mast cells retain their VMAT2 phenotype in cutaneous mastocytosis (Anlauf et al. 2004), we anticipated that VMAT2 might also serve as a marker for systemic mastocytosis, which is a persistent clonal disease of bone marrow–derived myelomastocytic progenitor cells and therefore differs from cutaneous mastocytosis (Valent et al. 2001a, 2004). The diagnosis mastocytosis is traditionally based on the demonstration of focal accumulations of mast cells with typical histological and cytomorphological properties (Lennert and Parwaresch 1979; Horny and Valent 2001; Valent et al. 2001a). However, depending on the organ system examined, type of mast cell disease, and presence of other diseases, it is sometimes difficult to diagnose mastocytosis histologically (Horny et al. 1985; Horny and Kaiserling 1988; Horny et al. 1989, 1992). Our study demonstrated that VMAT2 can be used as an excellent pathological marker not only of cutaneous, but also of systemic mastocytosis, comparable to the well established mast cell marker tryptase (Li et al. 1996; Horny et al. 1998).
CML is a myeloproliferative disease that originates from an abnormal pluripotent bone marrow stem cell. Most CML patients have increased numbers of mast cells, basophil granulocytes, and megacaryocytes (Vardiman et al. 2001). Our finding that VMAT2 expression is retained in these neoplastic cell types indicates that monoamine-storage mechanisms are conserved in this condition and that VMAT2 can be used as a marker to identify monoamine-handling cell populations. However, because VMAT2 is expressed in all mast cells, regardless of whether they are derived from patients or healthy controls, a distinction between normal mast cells and different subtypes of mastocytosis based on their VMAT2 expression patterns is not possible. So far it is not known whether VMAT2 is also expressed in hematological disorders other than mastocytosis and CML. It would be of interest to analyze in which maturation stages neoplastic hematopoietic cells in acute myeloid leukemia obtain monoaminergic properties, because it has been reported that some types of acute myeloid leukemia showed tryptase and CD117 expression and elevated serum levels of circulating tryptase (Sperr et al. 2001, 2002). However, this should be the subject of a further study.
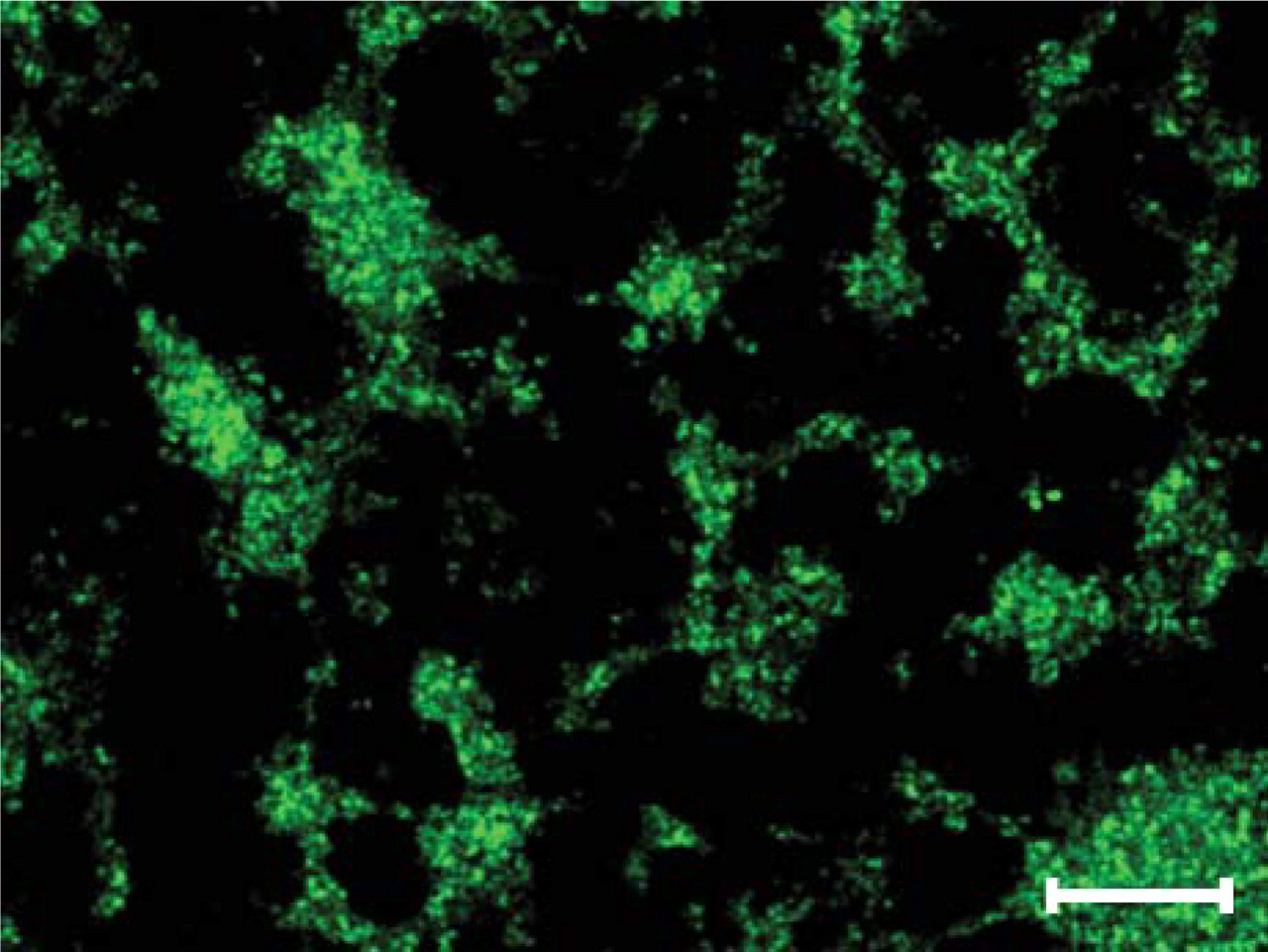
Expression of VMAT2 in thrombocytes. Confocal laser scanning double fluorescence microscopy for VMAT2 (green) showing a granular immunoreactivity of thrombocytes (pooled platelet concentrate). Bar = 150 μm.
Expression of VMAT2 in patients with cutaneous mastocytosis. (
It has been mentioned that increased VMAT2 binding can be visualized in the normal tonsils and in the nasal lymphoid tissue by positron emission tomography (Meyer et al. 2000). Our findings provide the morphological substrate for this observation, namely the enrichment of VMAT2-positive mast cells in these regions. Because we have shown in our study that VMAT2 is an excellent pathological marker for all types of mastocytoses and neoplastic monoamine handling cells, it should now be possible to develop positron emission tomography imaging strategies for the diagnosis of mastocytosis and myeloproliferative disorders.
In conclusion, the selective expression of VMAT2, which is responsible for the uptake and accumulation of biogenic amines in secretory granules to make them available for exocytotic release, is a new, important feature in the functional spectrum of mast cells, megakaryocytes, and basophil granulocytes. VMAT2 is a useful diagnostic tool for identifying normal mast cells and neoplastic mast cells in all different subtypes of mastocytosis. The identification of VMAT2 in cells of the hematopoietic system provides evidence of a molecular mechanism for monoamine storage and handling in these cells and provides a basis for imaging, as well as for promoting and interfering with monoamine loading of these cells, in myeloproliferative diseases such as CML.
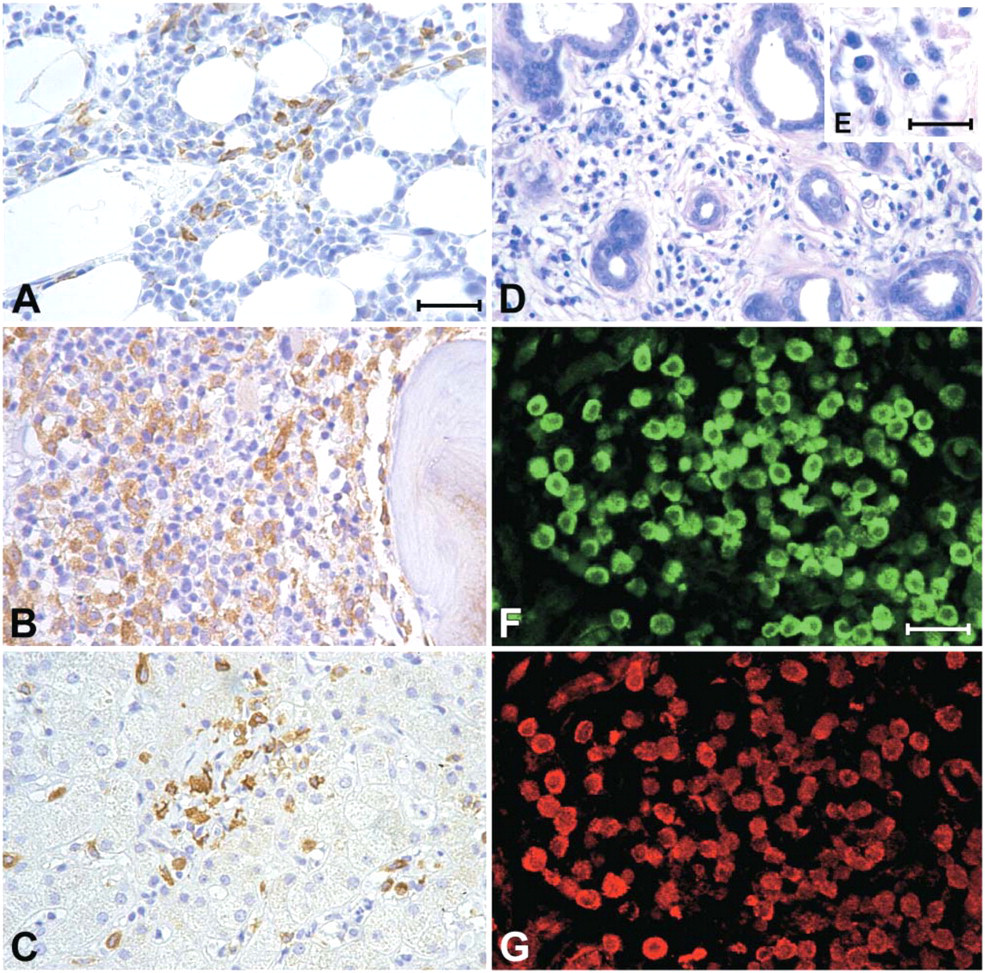
Expression of vesicular monoamine transporter (VMAT)2 in patients with systemic mastocytosis. (
Footnotes
Acknowledgements
Supported by the Volkswagen-Stiftung, Germany, and the University of Kiel, Germany (F 344101).
We would like to thank Maike Pacena, Anja Bredtmann, Tanja Hein, Heidi Hlavaty, Alice Johl, Heike Ristau, Elke Rodenberg-Frank, Petra Sack, and Marion Zibuschka for their excellent technical assistance, and Heidemarie Schneider and Waldemar Strauss for the brilliant photographic documentation. We are indebted to Katherine Dege for critically reading and editing the manuscript. We thank all colleagues who provided clinical data on the patients. Some of the results of this study are part of Viktoria Brand's MD thesis.