
Abstract
We investigated the histochemical characteristics of mast cell tryptase in different mouse tissues. By use of peptide substrates, tryptase activity could be demonstrated in unfixed connective tissue mast cells in different tissues, including the stomach. Tryptase activity was better localized after aldehyde fixation and frozen sectioning, and under such conditions was also demonstrated in mucosal mast cells of the stomach but not in those of the gut mucosa. Double staining by enzyme histochemistry followed by toluidine blue indicated that the tryptase activity was present only in mast cells and that all mast cells in the stomach mucosa contained the enzyme. The peptide substrates z-Ala-Ala-Lys-4-methoxy-2-naphthylamide and z-Gly-Pro-Arg-4-methoxy-2-naphthlyamide, which are substrates of choice for demonstrating tryptase in other species, were most effective for demonstrating mouse tryptase. The use of protease inhibitors further indicated that activity present in all mast cells was tryptase. Safranin O did not stain stomach mucosal mast cells, suggesting that the tryptase present in these cells was active in the absence of heparin sulfate proteoglycan.
T
Materials and Methods
Tissues
Gut, lung, prostate, skin, and stomach were removed from adult (12-week-old) male C3H, WHT, and C3A mice under terminal pentobarbital (IP) anesthesia.
Fixation
Pieces of the different tissues were snap-frozen in hexane cooled on solid CO2 and 10-μm frozen sections were cut in a cryostat. Different fixatives were assessed for the fixation of tryptase activity in other pieces of the tissues taken. Aldehyde fixation was performed either by (a) immersion of the material for 6-18 hr at 0-4 C in formol-sucrose containing 4% (w/v) formaldehyde freshly prepared from paraformaldehyde, 7% (w/v) sucrose in 0.05 M cacodylate buffer, pH 7.2, or (b) immersion in a mixture of formaldehyde-glutaraldehyde containing 2.5% (w/v) formaldehyde freshly prepared from paraformaldehyde, 0.2% (w/v) distilled glutaraldehyde in 0.05 M cacodylate buffer, pH 7.2. After fixation the tissues were washed overnight at 0-4C in 0.05 M cacodylate buffer containing 7.5% sucrose, pH 7.2. Nonaldehyde fixation was performed in Carnoy fixative (standard protocol 60% ethanol, 30% chloroform, 10% acetic acid) for 4 hr or overnight or in methanol overnight. Aldehyde-fixed tissues were either paraffin-embedded and sectioned (5 (μm) or frozen and cryostat-sectioned (10 (μm). Carnoy- and methanol-fixed tissues were paraffin-embedded.
Dye Binding
Sections were stained for 30 min in solutions of either alcian blue-safranin O (0.36% and 0.18%, respectively) in 1 M HCl-sodium acetate buffer (Csaba 1969) or 1% toluidine blue in 0.5 M HCl, pH 0.5. Alcian blue-safranin O-stained sections were rinsed in water and then dehydrated in tertiary butyl alcohol, cleared in xylene, and mounted in DPX, whereas toluidine blue stained sections were not rinsed before dehydration. Mast cells were also stained for 20 min in a 0.02% aqueous solution of Berberine at low pH, as described by Aldenborg and Enerbäck (1988). After staining, sections were rinsed in distilled water, mounted in unbuffered glycerol, and fluorescent mast cells were viewed under UV light microscopy using a 510-nm barrier filter.
Enzyme Histochemistry
For demonstration of tryptase amidolytic activity, tissue sections were incubated in 0.25 mg/ml of the substrate z-Ala-Ala-Lys-4-methoxy-2-naphthylamide (z-AAK-mna; Enzyme System Products; Dublin, CA) in 0.1 M phosphate buffer, pH 6.5, 5.5, or 5.0 for 60-120 min at 37C. The incubation medium contained Fast Blue B salt (final concentration 3.2 mM) as a capture reagent and incubation was followed by a 5-min wash in 1% CuSO4. The same protocol was applied for the substrates z-Gly-Pro-Arg-4-methoxy-2-naphthylamide (z-GPR-mna) and D-Val-Leu-Arg-4-methoxy-2-naphthylamide (D-VLR-mna). To demonstrate chymase amidolytic activity with the substrate suc-Ala-Ala-Phe-4-methoxy-2-naphthylamide (s-AAF-mna), the same protocol was followed at pH 7.8 in 0.1 M Tris-HCl, and for chymase esterolytic activity sections were incubated with N-acetyl-L-methionine-α-naphthyl ester (α- N-O-Met) for 0.5 hr at pH 7.8. After staining, the sections were rinsed in double-distilled water, dehydrated, cleared, and mounted in DPX.
Inhibitors
Selective inhibitors were used to characterize the enzyme activity detected (Kennett et al. 1993). Tissue sections were preincubated for 0.5 hr at 37C with the following trypsinlike protease inhibitors: 2 mM Tos-Lys-chloromethylketone or 0.03 mM D-Val-Leu-Arg-chloromethylketone or the chymotrypsin-like enzyme inhibitor Gly-Lys-Pro-chloromethylketone (at 0.6 mM), and then enzyme activity was detected in substrate medium (see above) containing the same concentration of inhibitor.
Double Staining
To estimate the number and distribution of tryptase-containing mast cells, sections were stained with z-Ala-Ala-Lys-mna, dehydrated, mounted, photographed, and afterwards were hydrated again and stained with 1% toluidine blue in 0.5 M HCl, pH 0.5, for 30 min, followed by dehydration, mounting, and photomicrography.
Results
Enzyme Histochemistry
With the use of amidolytic substrates, tryptase activity could be demonstrated in connective tissue mast cells in unfixed frozen sections of all tissues. Of the substrates tested, z-GPR-mna (Figure 1A) gave intense staining, and similar results were obtained with z-AAK-mna, whereas inferior faint staining was obtained with D-VLR-mna. Occasional diffuse staining, suggestive of mast cells, was seen in the unfixed stomach mucosa using z-AAK-mna and z-GPR-mna (Figure 1B). When tissues were fixed with formaldehyde-glutaraldehyde and frozen-sectioned, tryptase activity in connective tissue mast cells, and particularly in stomach mucosal mast cells, was better localized within distinct granules and the mucosal mast cells were seen to be present in intraepithelial sites (Figures 2A and 2B). Tryptase activity could not be demonstrated in aldehyde-, Carnoy-, or methanol-fixed tissues after paraffin embedding. Tryptase-containing mucosal mast cells were not observed in either unfixed or fixed gut. The tryptasecontaining mucosal mast cells in the stomach were examined in more detail. Like tryptase in connective tissue mast cells, the enzyme in stomach mucosal mast cells was best demonstrated with the substrates z-AAK-mna and z-GPR-mna, and when the pH of the enzyme histochemical incubation was lowered from 6.3 to 5.5 or 5.0 the intensity of the reaction product was decreased in mucosal mast cells of the stomach as in connective tissue mast cells (Figure 3). Incubation of the tissue sections with Tos-Lys-chloromethylketone, an inhibitor of trypsin-like enzymes, blocked the reaction with substrates z-Ala-Ala-Lys-mna, z-Gly-Pro-Arg-mna, and D-Val-Leu-Arg-mna because mast cells remained unstained (not shown). The amidolytic substrate s-AAF-mna and the esterolytic substrate α- N-O-Met demonstrated chymase activity in mast cells and the intensity of the reaction product was unaffected by incubation with the trypsin-like enzyme inhibitor Tos-Lys-chloromethylketone (Figure 4). However, such chymase activity was greatly reduced by the specific chymotryptic enzyme inhibitor Gly-Lys-Pro-chloromethylketone (not shown).
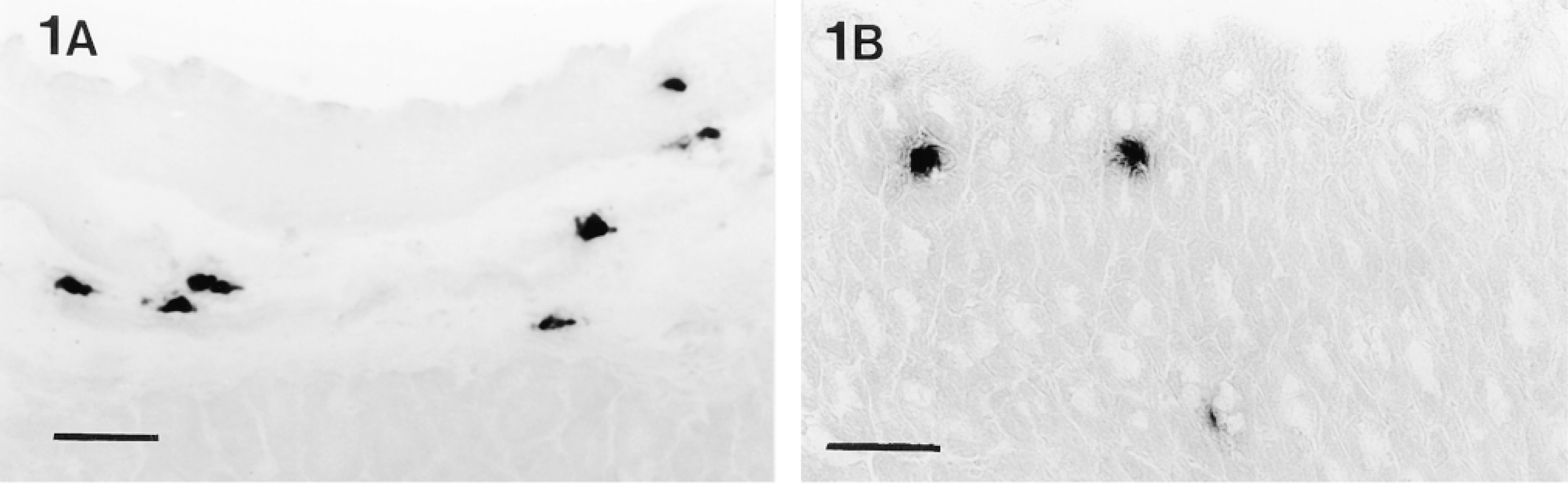
Tryptase activity, demonstrated with the substrate z-Gly-Pro-Arg-mna, in (A) submucosa and (B) mucosa of a frozen section (10-μm) of unfixed stomach. Staining of connective tissue mast cells in the submucosa is localized, whereas only faint diffuse staining is seen in mucosal mast cells. Bars = 50 μm.
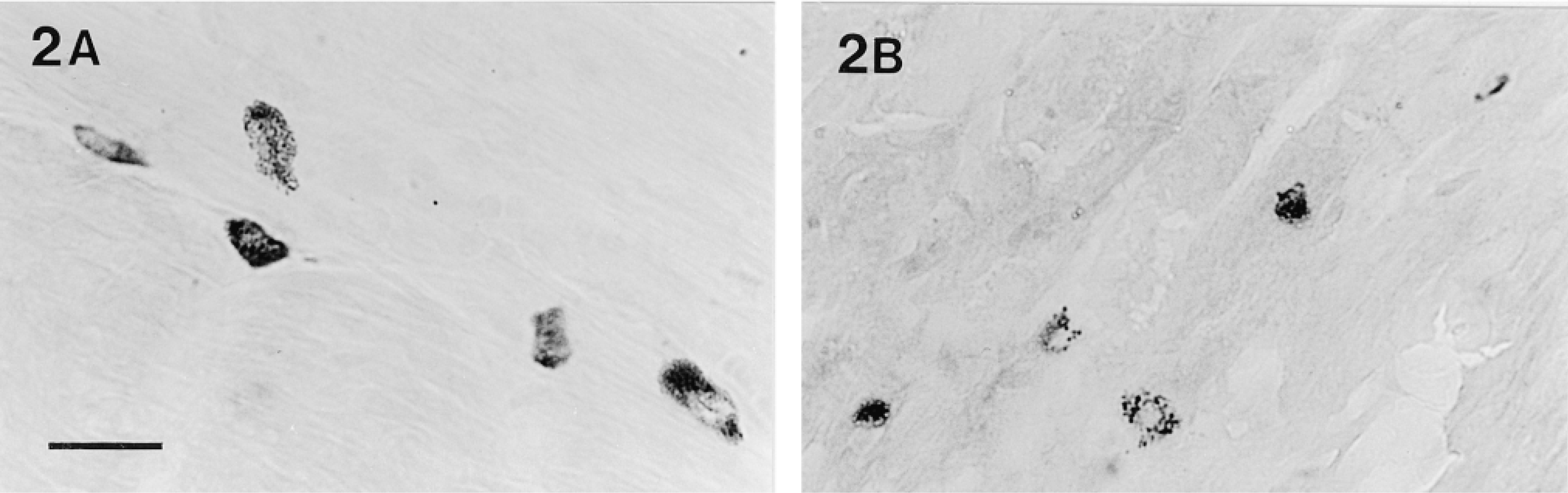
Tryptase activity, demonstrated with the substrate z-Ala-Ala-Lys-mna, in (A) submucosa and (B) mucosa of a frozen section (10-μm) of glutaraldehyde-fixed stomach. Staining of both the mucosal and connective tissue mast cells is well localized in cytoplasmic storage granules, and mucosal mast cells are localized in intraepithelial sites. Bar = 20 μm.

Tryptase activity, demonstrated with the substrate z-Ala-Ala-Lys-mna, in a frozen section (10-μm) of glutaraldehyde-fixed stomach. Enzyme histochemistry was performed at pH 5.0 and staining is much fainter than that seen at pH 6.3 (Figure 2). Bar = 20 μm.
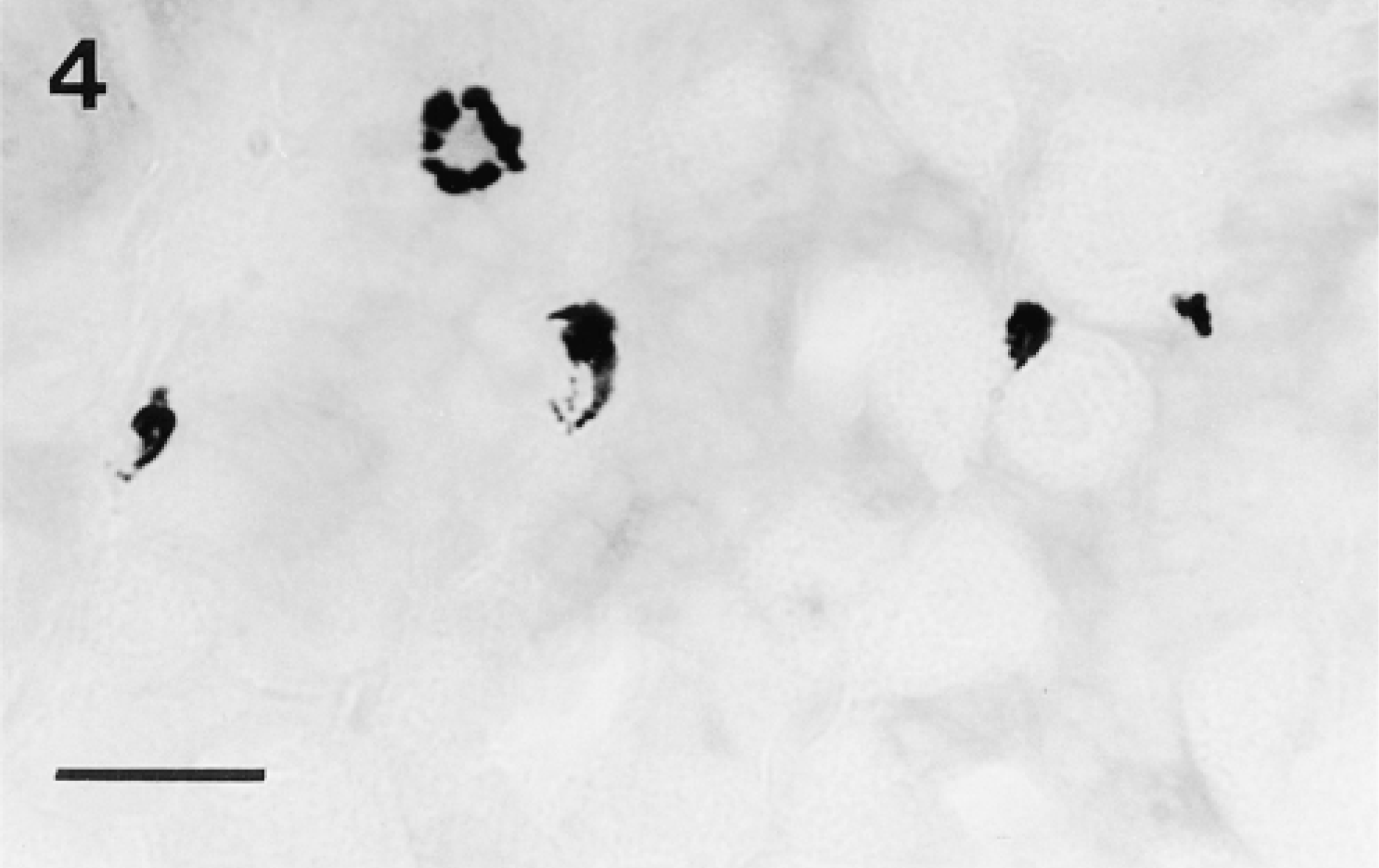
Chymase activity demonstrated with the substrate N-acetyl-L-methionine-alphanaphthyl ester in the presence of the trypsin-like enzyme inhibitor Tos-Lys-chloromethylketone in a frozen section (10-μm) of glutaraldehyde-fixed stomach. Staining is well localized in mucosal mast cells and is as intense as that seen in sections incubated in the absence of the inhibitor (not shown). Bar = 20 μm.
Dye Binding
To examine whether the tryptase-containing mast cells also contain heparin sulfate PG in their granules, aldehyde- and Carnoy-fixed tissues were stained with alcian blue-safranin O or Berberine sulfate. All connective tissue mast cells showed safranin or Berberine sulfate affinity, whereas the mucosal mast cells, including the tryptase-positive stomach mucosal mast cells, were negative (not shown). Both connective tissue and mucosal mast cells were demonstrated by toluidine blue staining of aldehyde-fixed, frozen-sectioned tissues. When used after enzyme histochemistry in a double staining protocol, toluidine blue staining showed that all tryptase activity was present only in mast cells and that all toluidine blue-positive stomach mucosal mast cells contained active tryptase (Figures 5A and 5B).
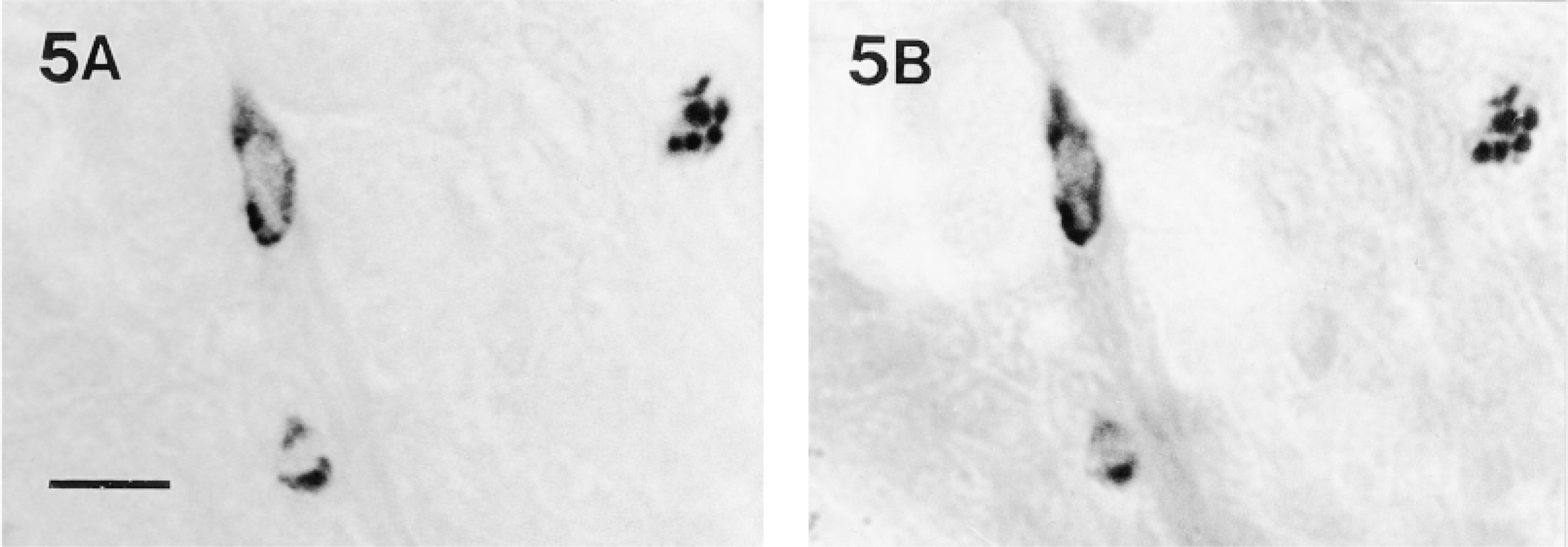
(A) Tryptase activity, demonstrated with the substrate z-Ala-Ala-Lys-mna, in a frozen section (10-μm) of formaldehyde-fixed stomach. (B) The same section as A after further staining with toluidine blue at low pH. All of the stomach mucosal mast cells demonstrated by toluidine blue contained tryptase activity. Bar = 20 μm.
Discussion
The amidolytic substrates z-AAK-mna and z-GPR-mna have previously been shown to be effective in specific demonstration of tryptase in mast cells of cat, human, ox, and rat, whereas the substrate D-VLR-mna has been shown to be less effective (Garrett et al. 1985; Proctor et al. 1991; Valchanov et al. 1998). With the same substrates, similar results have now been obtained in mouse. In a double staining technique with toluidine blue, tryptase enzymic activity has been demonstrated not only in murine connective mast cells but also in mucosal mast cells in the stomach, although not in mucosal mast cells of the gut. The enzyme in stomach mucosal mast cells has similar histochemical amidolytic substrate specificities and inhibitor characteristics to murine tryptase demonstrated in mouse connective tissue mast cells and tryptases of other species. Unlike human tryptase, but like rat tryptase, murine tryptase in both mucosal and connective tissue mast cells appears to be thermosensitive, because par affin embedding abolished activity. The optimal pH for trypsin-like enzymes is around neutrality, but to reduce nonspecific breakdown of the diazonium salt the histochemical coupling reaction is conducted at a pH below neutrality (Garrett et al. 1985). The neutral pH optimum of the enzyme in murine mast cells was demonstrated by the decreasing activity observed as pH was lowered from 6.3 to 5.5 and 5.0. The tryptic activity was also confirmed by using inhibitors of trypsin-like enzymes, which abolished staining, whereas inhibitors of chymotrypsin-like enzymes did not.
Human tryptase activity is dependent on heparin sulfate, with which it forms a complex (Schwartz and Bradford 1986). The link between the presence of tryptase activity in mast cells and heparin goes further because it has recently been found that heparin is required for processing of human tryptase to an active enzyme (Sakai et al. 1996). Heparin and tryptase appear to be present in all human mast cells (Irani et al. 1986; Craig et al. 1993). Safranin O and Berberine sulfate have been used as markers for heparin sulfate in connective tissue mast cells in rodents (Aldenborg and Enerbäck 1988), and we, like others, have found that stomach and gut mucosal mast cells are safranin O- and tryptase-negative in rat (Chen et al. 1993; Valchanov et al. 1998), indicating levels of heparin below the histochemical detection limit. Immunocytochemical localization of heparin in rat mast cells has also demonstrated an apparent absence from rat mucosal mast cells (Oliano et al. 1997). Therefore, there is strong evidence that the presence of tryptase in mast cells of most species is linked with the presence of heparin sulfate. The apparent absence of heparin sulfate PG from mucosal mast cells of the mouse, as indicated by our results and those of a previous study (Sonoda et al. 1986), suggests that the enzyme present in the stomach mucosal mast cells of mouse is not a conventional tryptase. At present, this conclusion remains tentative because we did not examine these mast cells for the presence of mouse mast cell protease 6 or 7, and there is evidence indicating that mouse mast cell proteases 6 and 7 cleave endogenous substrates in the absence of heparin (Huang et al. 1997,1998). However, others have found that mRNA for the most widespread mouse tryptase, mouse mast cell protease 6, is not present in stomach mucosal mast cells (Jippo et al. 1997). A difference between the tryptases present in mouse mucosal mast cells and connective tissue mast cells was suggested by the greater diffusion of the mucosal enzyme in the absence of aldehyde fixation, although this may also result from differences in the secretory granule content of other molecules, including heparin sulfate. A recent study by Friend et al. (1998) demonstrated that paraformaldehyde-fixed connective tissue mast cells expressing mouse mast cell protease 6 and/or 7 cleaved the substrate z-AAK-mna. In the same study, mast cells showed altered protease expression, including the presence of the tryptases mast cell proteases 6 and 7, as they migrated through the jejunal mucosa during recovery from Trichinella spiralis infection. However, such tryptase-expressing mast cells were absent from jejunal mucosa of normal control mice. The present enzyme histochemical study suggests that a population of tryptase-positive but heparin sulfate-negative mast cells is present in normal mouse stomach mucosa. Further characterization of the enzyme will be required to determine if it is one of the identified mouse tryptases, mouse mast cell protease 6 or 7, or another member of this family.
Footnotes
Acknowledgements
Supported by Grants from the International Journal of Experimental Pathology and the European Federation of Experimental Morphology.
We thank Katherine Paterson and Robert Hartley for their assistance.