
Abstract
Oviductins belong to a family of glycoproteins that have been suggested to play several roles during the early processes of reproduction. Recently, a polyclonal antibody was raised against recombinant hamster oviductin (rhaOvm). Here the anti-rhaOvm antibody was used to investigate the sites of localization of oviductin in the female golden hamster. In the hamster oviduct, immunolabeling was restricted to the content of the Golgi saccules and secretory granules of the non-ciliated oviduct cells. After its release into the lumen, oviductin becomes associated with the zona pellucida of post-ovulatory oocytes. In unfertilized oocytes, oviductin was also detected in membrane invaginations along the oolemma and in some vesicles within the ooplasm. Furthermore, oviductin was detected over the microvilli and within multivesicular bodies of uterine epithelial cells. Western blotting analysis revealed the presence of oviductin in the hamster oviduct but not in the uterus or ovary. In the oviduct, the anti-rhaOvm antibody detected a polydispersed band corresponding to native oviductin (160–350 kD) and several lower molecular weight bands (<100 kD) corresponding to nascent and partially glycosylated forms of oviductin. The anti-rhaOvm antibody provides an additional tool for investigation into the cytochemical and biochemical properties of different forms of hamster oviductin in the female reproductive tract.
T
Once released into the lumen of the oviduct, these oviduct glycoproteins are known to associate with the luminal cilia extending from the surface of the ciliated cells of the oviduct columnar epithelium (Kan et al. 1988,1989) and with various structures of the oocyte and early embryo, including the zona pellucida of hamster (Araki et al. 1987; Léveillé et al. 1987; Oikawa et al. 1988; Kan et al. 1988,1989,1990), human (O'Day-Bowman et al. 1996), baboon (Boice et al. 1990), bovine (Wegner and Killian 1991), pig (Brown and Cheng 1986; Buhi et al. 1993), sheep (Gandolfi et al. 1991), and goat (Abe et al. 1995). Oviductin has also been found in association with the perivitelline space of hamster (Abe and Oikawa 1990), mouse (Kapur and Johnson 1988), baboon (Boice et al. 1990), and pig (Buhi et al. 1993) and with microvilli of the egg plasma membrane in hamster (Kan et al. 1988,1989), baboon (Boice et al. 1990), and pig (Buhi et al. 1993). In addition, oviductin was detected within endocytic compartments, including membrane invaginations, coated vesicles, and multivesicular bodies of the fertilized egg and early embryo in some species (Kan et al. 1993; Boice et al. 1990) but not others (Gandolfi et al. 1991; Buhi et al. 1993). In the golden hamster, oviductin is associated with the uterine epithelium, which has been attributed to the flow of oviduct fluid (Martoglio and Kan, 1996). There is no expression of oviductin in ovarian (pre-ovulatory) oocytes and the surrounding granulosa cells of ovarian follicles (Araki et al. 1987; Kan et al. 1989; Abe and Oikawa 1990). Because mammalian oviductins share a common property of being highly glycosylated macromolecules, it has been suggested that different forms of oviductins, varying in number and distribution of oligosaccharide side chains, may be responsible for their diverse biological functions (Buhi 2002).
Recently, we have produced a polyclonal antibody against recombinant hamster oviductin that recognizes a peptide portion of hamster oviductin (McBride et al. 2004). The newly developed anti-rhaOvm antibody differs from the previously used monoclonal antibodies, which recognize glycosidic epitopes attached to the protein backbone (Araki et al. 1987; Kan et al. 1988,1989,1993; St-Jacques and Bleau 1988; Abe and Oikawa 1990; Martoglio and Kan 1996). The anti-rhaOvm antibody proved useful in allowing a more accurate determination of oviductin levels in the secretory granules of the non-ciliated oviduct epithelial cells when the antibody was used in conjunction with quantitative immunocytochemistry (McBride et al. 2004). The previous study revealed that there are various forms of oviductin in the hamster oviduct and that not all forms are glycosylated (McBride et al. 2004). Therefore, the objective of the current study was to investigate the sites of localization of the various forms of oviductin in the female hamster reproductive tract and post-ovulatory oocytes employing the newly developed polyclonal antibody against recombinant hamster oviductin.
Materials and Methods
Animals
Female golden hamsters (
Preparation of Tissues for Immunohistochemistry and Immunocytochemistry
For immunohistochemical (IHC) and immunocytochemical (ICC) studies, sexually mature, untreated female hamsters were selected at random. The animals were sacrificed by cervical dislocation and the ventral abdominal wall was cut open and the ovaries, uteri, and oviducts were excised and rinsed in PBS (0.01 M, pH 7.2). For IHC studies, the uterus, ovary, and oviduct from one side of each animal were fixed in 4% paraformaldehyde (in 0.2 M phosphate buffer) overnight, followed by paraffin embedding according to routine procedures. Sections of 5-μm thickness were prepared and placed on microscope slides for IHC labeling. For ICC studies, the oviducts and uteri from the opposite sides of each animal used above were isolated and processed as described previously (Roux and Kan 1995). The tissues were embedded in Lowicryl K4M (J.B.E.M.; Montreal, Quebec, Canada) according to routine procedures. Ultrathin sections of a straw interference color were cut and mounted on formvar-coated nickel grids for immunolabeling.
Preparation of Post-ovulatory Oocytes for ICC
For the collection of post-ovulatory oocytes, sexually mature female golden hamsters (Charles River) were superovulated by administration of an IP injection of 25IU PMSG, followed by 25IU hCG 48 hr later, as described previously (Kan et al. 1989). The hamsters were sacrificed by cervical dislocation 17 hr after hCG injection. The cumulus masses containing the post-ovulatory oocytes were collected from the ampullary portion of the oviducts and rinsed in PBS. Cumulus masses containing post-ovulatory oocytes were processed and embedded in Lowicryl K4M as described above.
Polyclonal Antibody
A polyclonal antibody prepared against recombinant hamster oviductin (anti-rhaOvm antibody; McBride et al. 2004) was used in the present study for immunolocalization studies and Western blotting analysis. The development and specificity of this polyclonal antibody have been recently described (McBride et al. 2004). In brief, the polyclonal antibody was raised against recombinant hamster oviductin (haOvm) prepared from a cDNA fragment coding for the region encompassing nucleotides 1140–2276 that contain the mucin-like domain (McBride et al. 2004). The nucleotide sequence of the haOvm fragment is unique to hamster oviductin and does not resemble any other glycoproteins. The specificity of the affinity-purified anti-haOvm antibody has been established by Western blotting analysis (McBride et al. 2004).
IHC Detection of Hamster Oviductin
Paraffin-embedded sections were subjected to IHC staining for oviductin by application of the avidin-biotin complex technique. Sections were deparaffinized and rehydrated through a graded series of ethanol solutions in decreasing concentrations. The antigenic sites were enhanced by incubation of sections in 0.1% CaCl2 containing 0.1% pepsin for 15 min at 37C. Sections were blocked with 10% normal goat serum (NGS in PBS) for 20 min. Next, endogenous avidin- and biotin-like substances were blocked using the Avidin/Biotin Blocking kit (Zymed; San Francisco, California). Sections were incubated with anti-rhaOvm antibody (1:100 dilution in PBS containing 2.5% NGS) for 1.5 hr. After labeling with primary antibody, the sections were washed with PBS to remove unbound antibody and incubated with biotinylated goat anti-rabbit IgG (1:100 dilution in PBS containing 2.5% NGS) for 10 min. Endogenous peroxidase activity was blocked by incubating sections for 45 min in 1% H2O2 in H2O. Then the sections were washed with PBS and incubated with streptavidin-horseradish peroxidase conjugate (1:300 dilution) for 10 min. The immunoperoxidase color reaction was developed by incubation of sections in diaminobenzidene (DAB) substrate chromogen solution (Zymed). The sections were counterstained with hematoxylin (Counterstain Panel; Zymed) and further processed according to routine procedure. To assess the specificity of the anti-rhaOvm antibody, control experiments were performed by incubation of sections with diluted preimmune serum instead of with primary antibody. All other steps and conditions remained the same.
Immunogold Labeling
Ultrathin Lowicryl sections of hamster oviducts, uteri, and cumulus masses were immunolabeled for oviductin by incubating sections with anti-rhaOvm antibody (diluted 1:5 in PBS), followed by protein A-colloidal gold complex (diluted 1:10 in PBS) as described previously (Roux and Kan 1995). The sections were counterstained with uranyl acetate and lead citrate before examination on a Hitachi 7000 electron microscope operated at 75 kV. Control experiments were performed using preimmune serum instead of anti-rhaOvm antibody.
Isolation of Total Protein from Oviduct Tissue
The oviducts used for isolation of total protein were collected from sexually mature, untreated female golden hamsters. The oviducts were separated and then rapidly frozen and stored in liquid nitrogen until use. The oviducts were homogenized at 4C in homogenization buffer (50 mM Tris-HCl/150 mM NaCl, pH 8.0, containing 0.02% NaN3, 35 μg/ml aprotinin, 0.2 mM phenylmethanesulphonyl fluoride (PMSF), 1 μM leupeptin, 100 μM EDTA, and 0.2 mM benzamidine) as described previously (Malette and Bleau 1993). The homogenates were transferred to a microfuge tube and centrifuged at 44,000 rpm at 4C for 1 hr. The supernatants were transferred to new tubes and stored at –70C. Oviductin was further purified from samples of oviduct total protein homogenates using
SDS-PAGE, Transfer, and Immunodetection of Hamster Oviductin
Protein samples containing 25 μg total protein from hamster oviduct, as well as 3 μg of purified hamster oviductin, were prepared and separated by one-dimensional SDS-PAGE under reducing conditions as described by Laemmli (1970). The separated proteins were electrophoretically transferred to PVDF membranes under cooling conditions according to the procedure described by Towbin et al. (1979). After transfer of proteins, the membranes were blocked with 10% BSA in 10 mM TBS buffer (pH 8.0), followed by incubation with diluted anti-rhaOvm antibody (1:1000 in TBS). The membranes were washed in TBST buffer (10 mM Tris-HCl, 0.9% NaCl, 0.2% Tween-20; pH 8.0) before incubation with alkaline phosphatase-conjugated goat anti-rabbit IgG (1:10,000) for 1 hr. For color development, the membranes were equilibrated in detection buffer (0.1 M Tris-HCl, 0.1 M NaCl, pH 9.5), followed by incubation with NBT and BCIP (Roche Diagnostics; Laval, Quebec, Canada).
Results
IHC and ICC Detection of Hamster Oviductin
The newly developed anti-rhaOvm antibody was used to examine the distribution of oviductin in the female hamster reproductive tract and in post-ovulatory oocytes. As revealed by results obtained with IHC, the anti-rhaOvm antibody strongly labeled the oviduct epithelium on immunoperoxidase-labeled tissue sections (Figure 1A). At lower magnification, the oviduct epithelium exhibited an intense immunoreaction (not shown), whereas at higher magnification a heterogeneous labeling pattern was apparent, demonstrating that not all oviduct cells were labeled; strongly immunoreactive cells were observed interspersed with non-immunoreactive cells (Figure 1A). The ovary (Figure 1B) was unlabeled. However, faint immunostaining was detected over the uterine epithelial cell surface (Figure 1C). Immunoreaction was absent in control sections labeled with preimmune serum (Figure 1D).
To precisely locate the sites of immunoreactivity in the oviduct and uterus, an ICC study was performed. Application of the anti-rhaOvm antibody to thin sections of Lowicryl-embedded hamster oviduct, followed by incubation with protein A–gold complex, resulted in specific labeling. In the non-ciliated secretory cells of the oviduct epithelium, labeling was observed over the content of the secretory granules located in the apical portion of the cells (Figure 2B) as well as those positioned adjacent to the Golgi region (Figure 2B). The contents of the Golgi saccules were also immunolabeled (Figure 2B), whereas the microvillar and lateral cell membranes (Figure 2B) were essentially devoid of immunolabeling. The ciliated cells of the epithelium were not labeled with the anti-rhaOvm antibody (Figure 2A). Therefore, the labeling specificity of the anti-rhaOvm antibody for oviductin was established. In the uterine sections, labeling was mainly restricted to the apical cell surface, specifically over the microvilli of the uterine epithelium (Figure 3A). In addition, labeling was detected in a number of multive-sicular bodies within the uterine epithelial cells (Figure 3B). Labeling was not detected in the Golgi apparatus of the uterine epithelial cells (Figure 3C). Control sections of hamster oviduct and uterus exhibited very few, randomly distributed gold particles over the oviduct sections (Figure 3D).
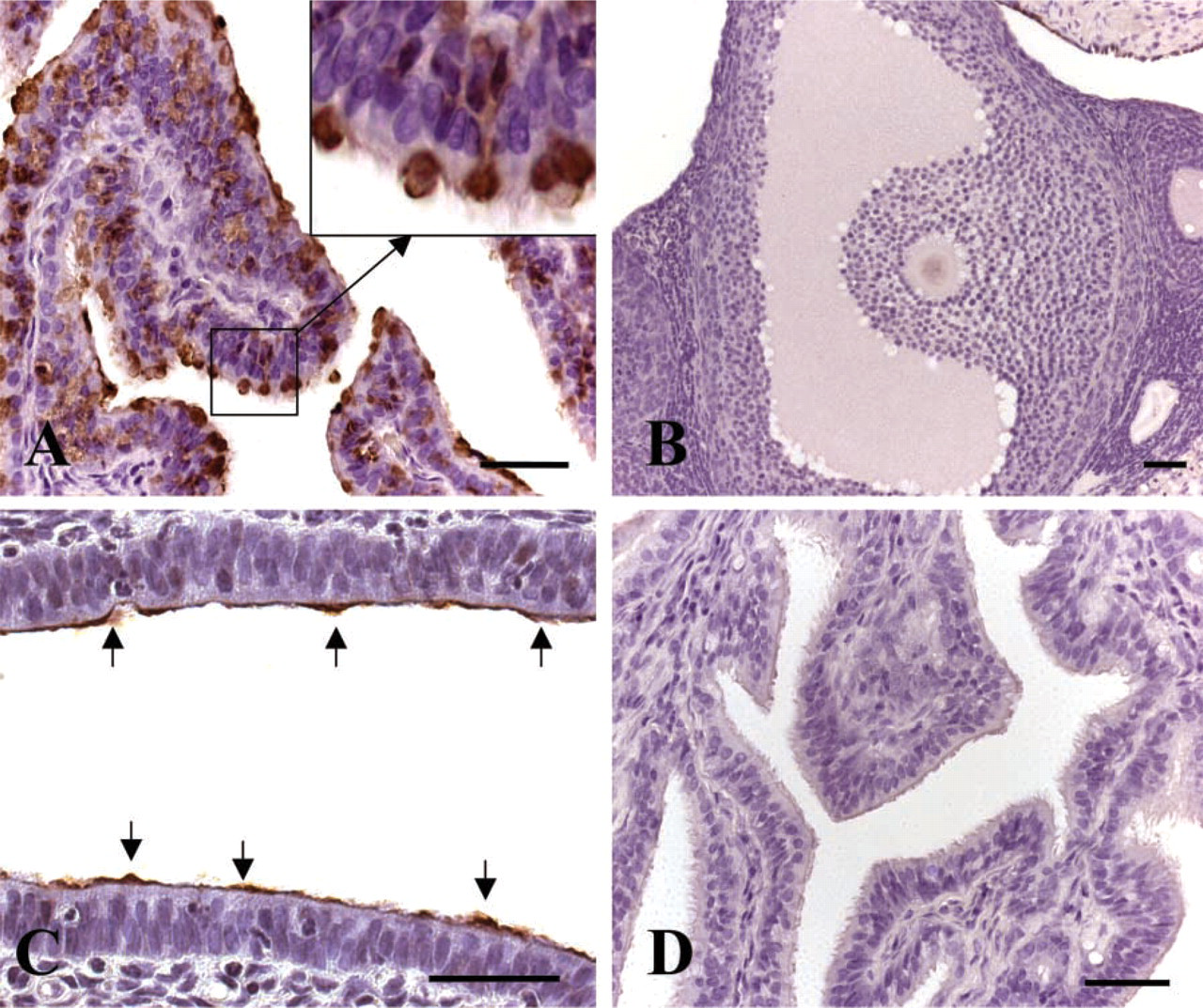
Light microscope photographs of paraffin sections of the female hamster reproductive tract. (
Application of the anti-rhaOvm antibody to sections of post-ovulatory oocytes resulted in an intense immunoreaction over the zona pellucida surrounding the oocyte (Figures 4A and 4B). The distribution of gold particles within the zona pellucida was uniform and homogeneous and the labeling appears to be associated with dense filamentous structures of the zona matrix (Figure 4B). Gold particles representing anti-genic sites were detected in the perivitelline space and in association with the microvilli of the oocyte plasma membrane (Figure 4A). Gold particles were also observed in association with membrane invaginations along the oolemma and in some vesicles of the post-ovulatory oocytes (Figure 4A). Control sections showed an absence of labeling over the zona pellucida, perivitelline space, and microvilli of the oocytes (Figure 4C).
Examination of Oviductin Protein Expression in the Reproductive Tract Tissues
To test the specificity of the anti-rhaOvm antibody against purified hamster oviductin and to examine the protein expression of oviductin in the female hamster reproductive tract, purified oviductin samples and oviduct, uterine, and ovarian total protein samples were prepared and analyzed by Western blotting. The anti-rhaOvm antibody detected a broad band extending from ≃160–350 kD in the lane containing purified hamster oviductin (Figure 5, Lane 1), further demonstrating the specificity of the newly developed antibody for hamster oviductin. The anti-rhaOvm antibody detected oviductin in the oviduct protein lysates (Figure 5, Lane 4) but not in the uterine or ovarian protein lysates (Figure 5, Lanes 2 and 3, respectively). In the oviduct total protein samples, the anti-rhaOvm antibody reacted with the polydispersed band ranging from 160 to 350 kD, corresponding to the fully glycosylated form of oviductin (Figure 5, Lane 4). In addition, the antibody was immunoreactive with several bands between 70 and 90 kD (Figure 5, Lane 4) corresponding to the nascent glycoprotein and various forms of glycosylated hamster oviductin.
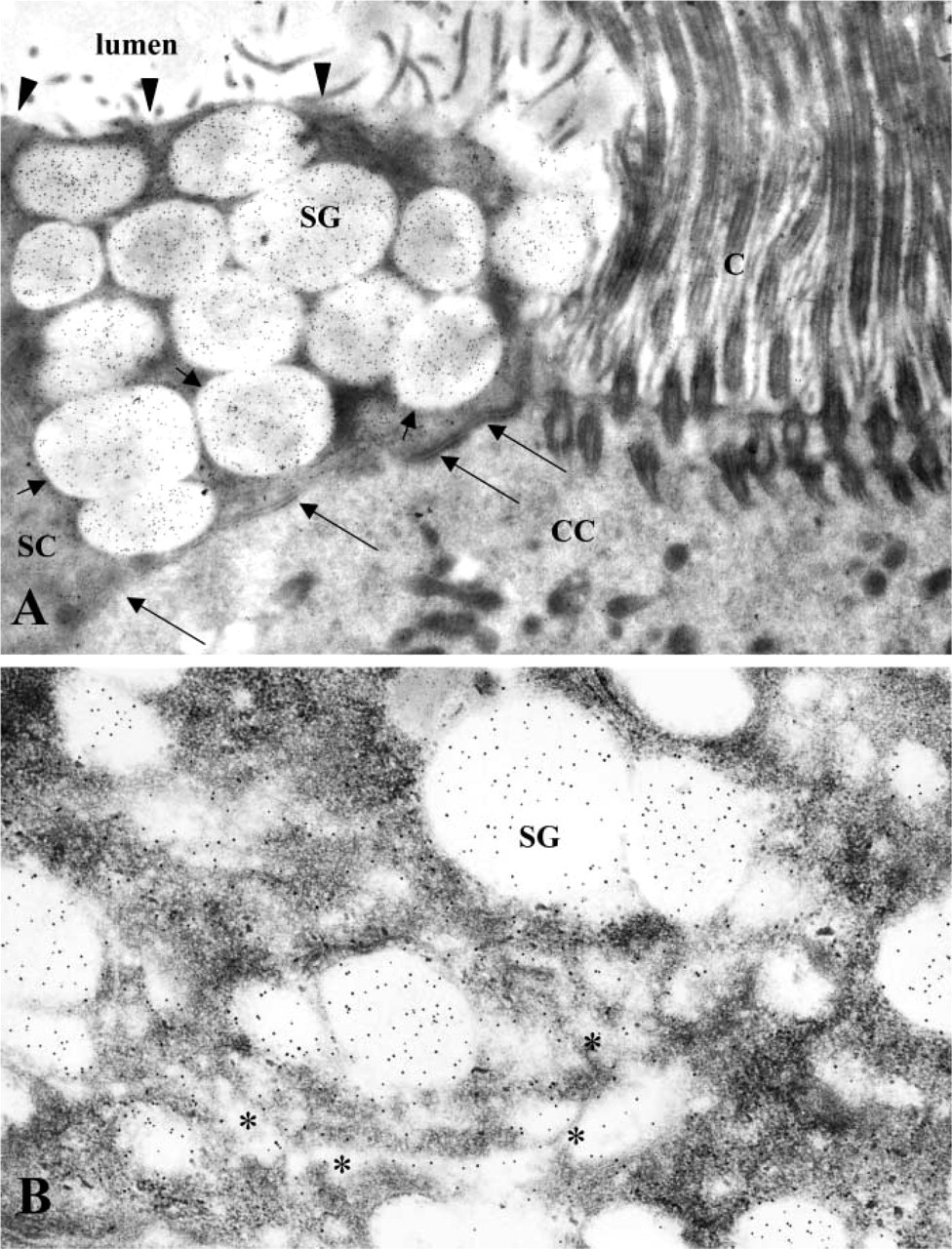
Electron photomicrographs of ultrathin Lowicryl sections of hamster oviduct showing immunolabeling for hamster oviductin by anti-rhaOvm antibody. (
Discussion
In the present study, the recently developed anti-rhaOvm antibody (McBride et al. 2004) was used to investigate the presence of oviductin in the female golden hamster reproductive tract. The epitope recognized by the anti-rhaOvm antibody is located on the polypeptide core within the mucin domain. The peptide sequence recognized by the anti-rhaOvm antibody is encoded by nucleotides 1140–2276 of the complete cDNA sequence of hamster oviductin (McBride et al. 2004). Therefore, it is expected that the anti-rhaOvm antibody is capable of labeling the fully glycosylated, partially glycosylated, and nascent forms of the glyco-protein.
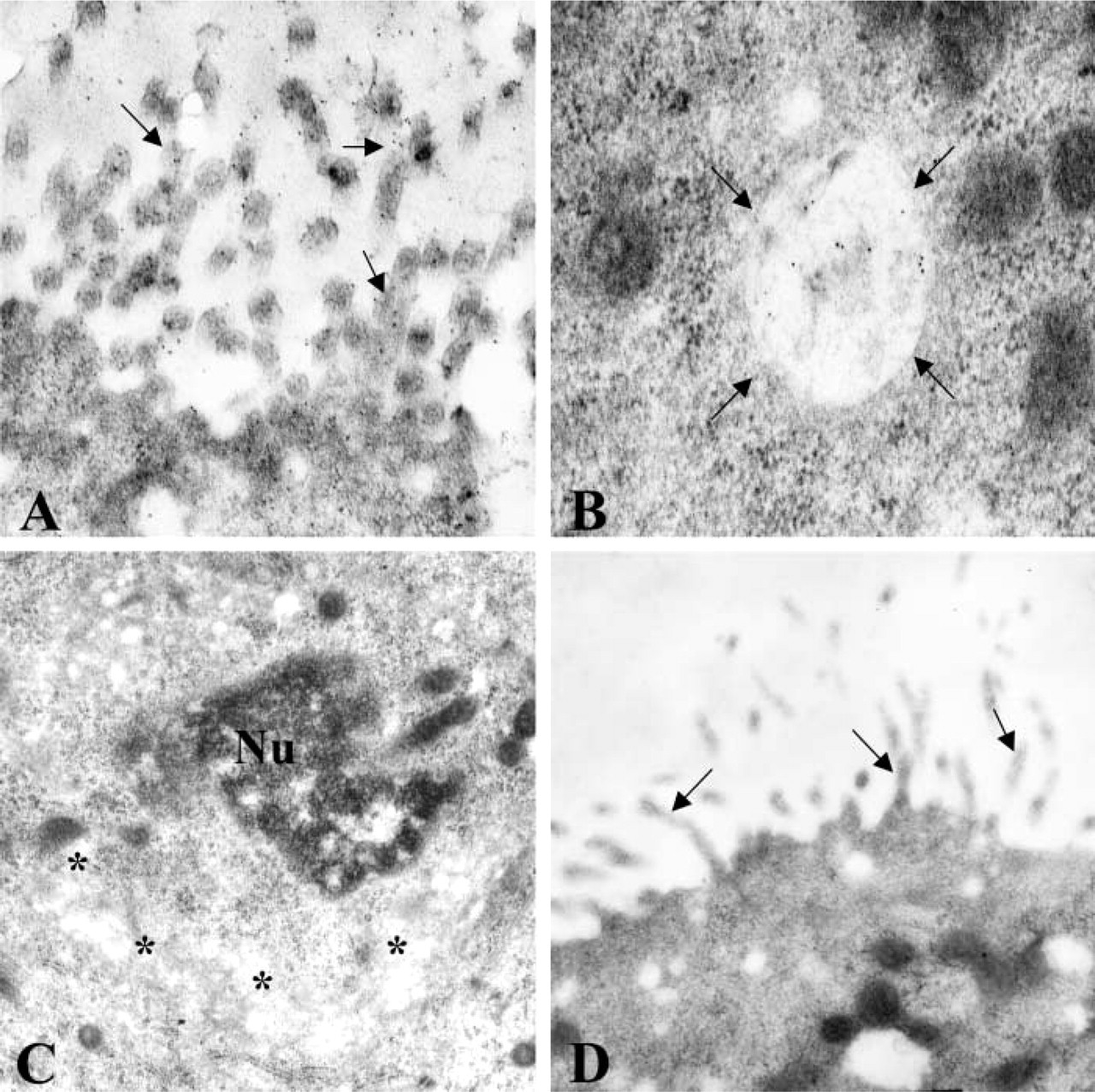
Electron micrographs of ultrathin Lowicryl sections of hamster uterus after immunolabeling with anti-rhaOvm antibody. (
IHC labeling using the anti-rhaOvm antibody produced an intense and heterogeneous labeling pattern in the oviduct epithelium, revealing the presence of oviductin only in the non-ciliated secretory cells and not in the ciliated cells of the columnar epithelium lining the oviduct lumen. This was confirmed by results obtained with immunolabeling at the electron microscopic level. Application of the anti-rhaOvm antibody to ultrathin sections of hamster oviduct demonstrated an intense labeling in the non-ciliated oviductal cells at the ultrastructural level. The anti-rhaOvm antibody labeled the content of the secretory granules and the interior of the Golgi saccules but not the granule membrane, Golgi membrane, or lateral cell membrane, indicating that the antibody recognizes a secretory protein and not a membrane-bound protein, and further demonstrating the specificity of the newly developed antibody.
With the immunoperoxidase labeling technique, the glycoprotein was not detected in the ovary, but a faint immunoreaction was noted at the cell surface of the epithelial lining of the endometrium. At the electron microscopic level, application of the anti-rhaOvm antibody to Lowicryl-embedded sections of uterus revealed the presence of gold particles over the microvilli and in multivesicular bodies of the uterine epithelial cells. Oviductin was not detected in the Golgi complex of these cells, indicating that the uterine epithelium does not synthesize oviductin. These findings represent the first to demonstrate the presence of oviductin in endocytic vesicles in the uterine epithelial cells of the non-pregnant hamster at the electron microscopic level. A previous study carried out in our laboratory, using a monoclonal antibody that recognizes a glycosidic epitope of hamster oviductin (St-Jacques et al. 1992), reported the localization of oviductin in the endometrial lining (Martoglio and Kan 1996). The presence of oviductin on the surface of the uterine epithelium was attributed to the flow of oviduct fluid from the oviduct into the lumen of the uterus. A subsequent study on the fate of hamster oviductin carried out in our laboratory using the monoclonal antibody showed that, during early gestation, immunoreactivity to oviductin in the uterus diminished to an almost total disappearance at the time of implantation (Roux et al. 1997). The current results suggest that oviductin is taken up by the uterine epithelium, although the reason for this is unknown and requires further investigation. However, the present results, taken together with our previous findings of the decrease of oviductin along the uterine epithelium at the time of blastocyst attachment and its final disappearance at implantation (Roux et al. 1997), suggest that oviductin could be a potential modulator of uterine receptivity.
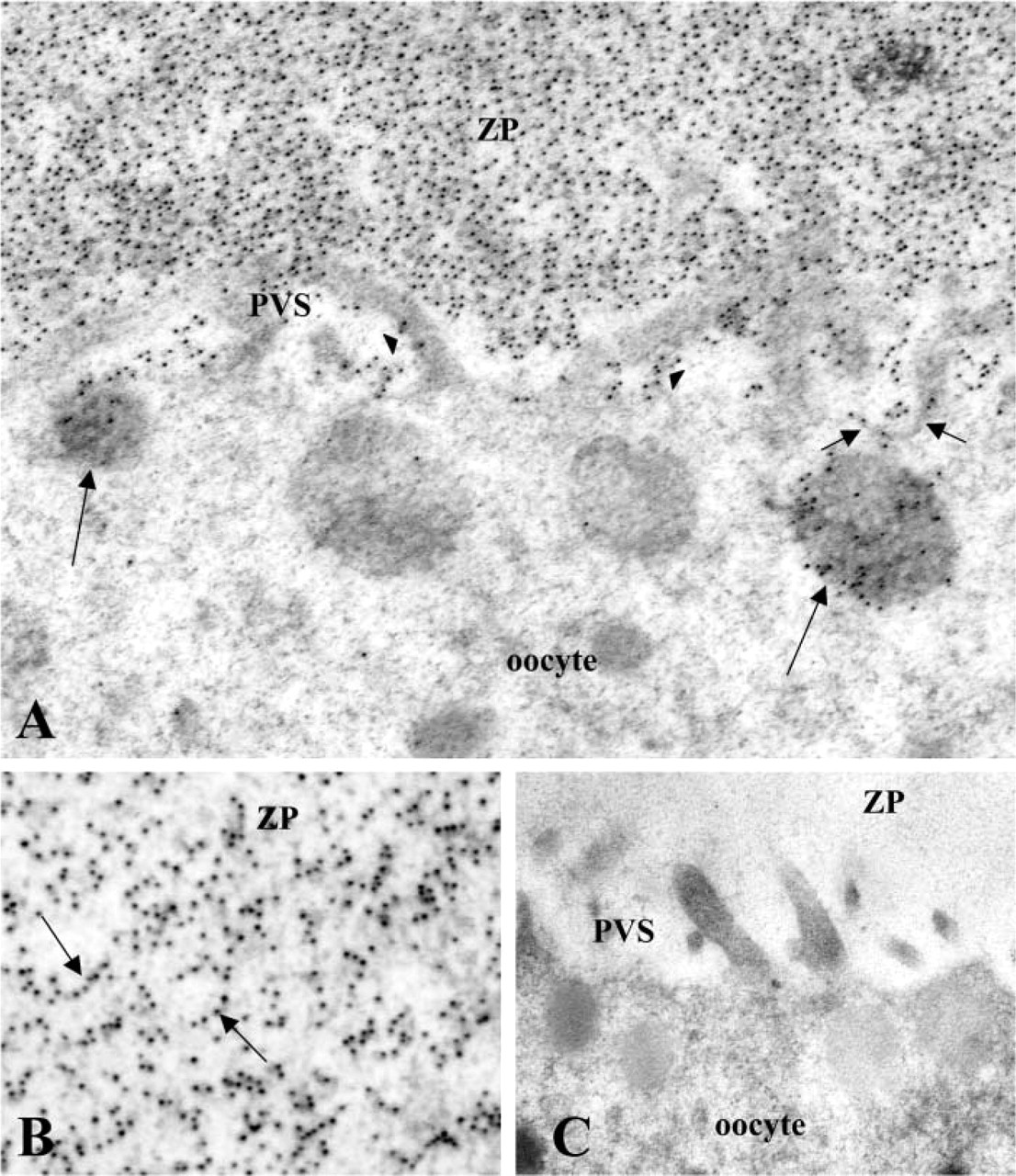
Electron micrographs of ultrathin Lowicryl sections of post-ovulatory oocytes incubated with anti-rhaOvm antibody. (
In the present study, oviductin was uniformly distributed throughout the zona pellucida of post-ovulatory oocytes and was found to be specifically associated with dense filamentous structures comprising the zona matrix. The presence of the glycoprotein was also detected on the microvilli of the post-ovulatory oocyte, in the perivitelline space, and in various intra-cellular compartments, including membrane invaginations and some vesicles. These labeled vesicles might be of the endocytic type because neighboring cortical granules were unlabeled. Our study is the first to describe the endocytosis of oviductin by unfertilized, post-ovulatory hamster oocytes. Previous studies using the monoclonal antibody showed an association of oviductin with the zona pellucida and the microvilli of the oocyte (Kan et al. 1988,1989) and in the perivitelline space (Abe and Oikawa 1990). The endocytosis of oviductin by fertilized oocytes and early embryos but not unfertilized oocytes has been reported (Kan et al. 1993; Kan and Roux 1995). The differences in the localization of oviductin observed between the current and previous studies may be attributed to the epitopes recognized by the different antibodies used. At this time, it is not clear as to the biological significance of the endocytosis of oviductin by the oocyte. It has been proposed that the endocytosis of oviductin may support the development and differentiation of early embryos (Kan and Roux 1995). Previous studies have shown the beneficial effects of oviductin in the enhancement of embryonic development (Nancarrow and Hill 1995; Kouba et al. 2000). A most recent study carried out in the pig demonstrated the embryotrophic effects of porcine oviductin when the protein was added to embryo culture medium after fertilization had occurred (McCauley et al. 2003). Future studies using specific isoforms of oviductins will be necessary to determine the mechanisms that regulate embryonic development and/or embryotrophic effects of oviductins.
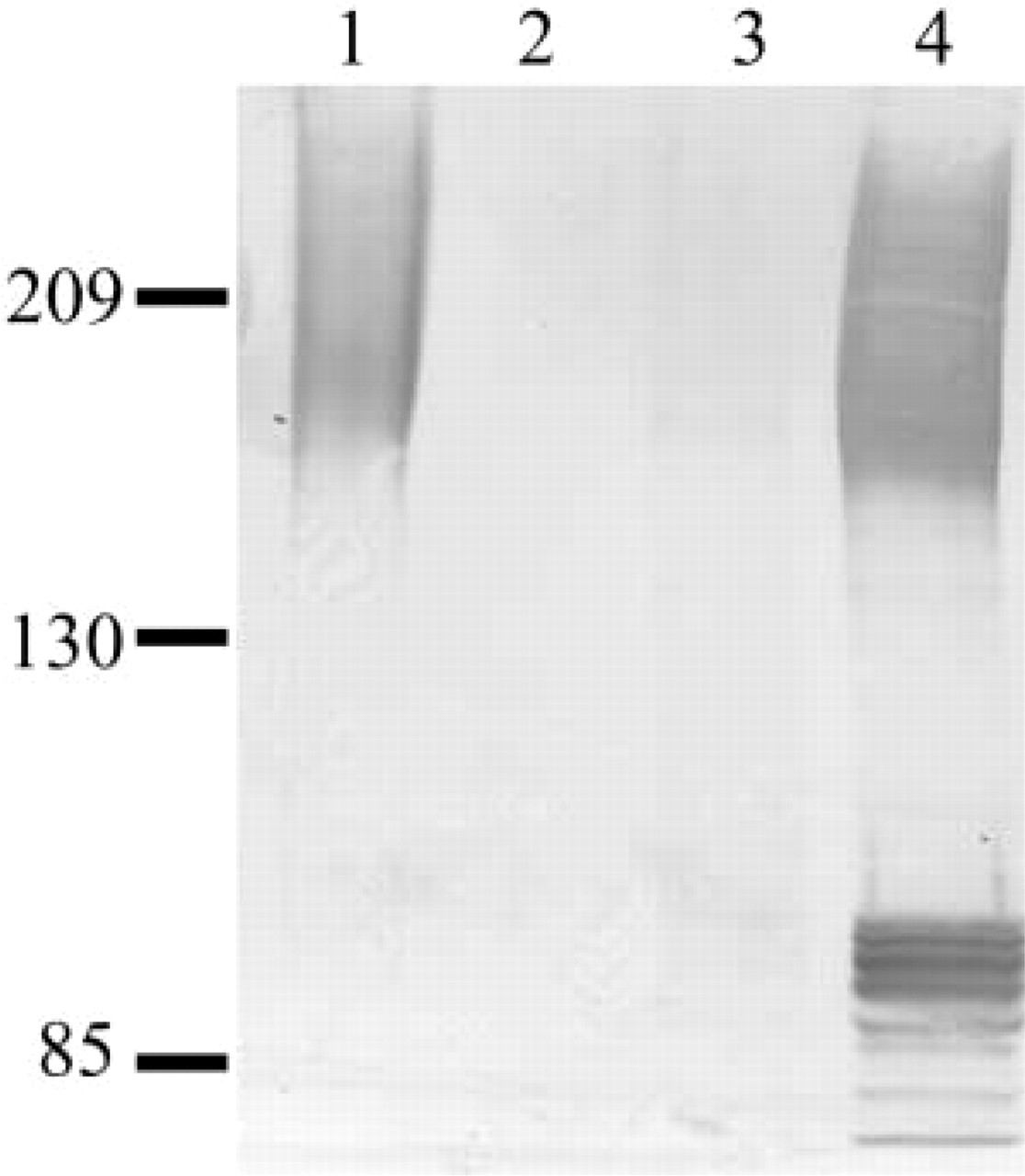
Western blot examining oviductin protein expression in the female hamster reproductive tract using anti-rhaOvm antibody. Oviductin is present in the oviduct but absent from the ovary and uterus. Lane 1, purified oviductin from the hamster oviduct; Lane 2, total protein from uterus; Lane 3, total protein from ovary; Lane 4, total protein from oviduct.
The presence of various forms of oviductin in the hamster oviduct was examined by Western blotting analysis. After immunoblotting, the anti-rhaOvm antibody detected a broad band starting at ≃160 kD in the HPA-purified hamster oviductin sample. This band corresponds to the mature, fully glycosylated form of oviductin as previously reported after detection with a monoclonal antibody (Robitaille et al. 1988; St-Jacques and Bleau 1988; Malette and Bleau 1993). There were several bands observed in the oviduct total protein homogenates, including the high molecular weight polydispersed band beginning at ≃160 kD corresponding to the band observed with HPA-purified oviductin. In addition, several lower molecular weight bands were detected, ranging from 70 to 90 kD. The lower bands most likely correspond to the nascent (non-glycosylated) form and partially glycosylated forms of the protein core. In the hamster, the nascent protein has an estimated molecular mass of 70.89 kD (Suzuki et al. 1995). Previously, the lower molecular weight forms of oviductin were not detected with the monoclonal antibody (Malette and Bleau 1993), suggesting that the anti-rhaOvm antibody can detect various forms of oviductin including nascent, partially glycosylated, and fully glycosylated forms.
In summary, we have used the polyclonal anti-rhaOvm antibody to locate the polypeptide in the female hamster reproductive tract. We have demonstrated the presence of oviductin in endocytic compartments of both unfertilized oocytes and uterine epithelial cells, reinforcing the notion that mammalian oviductins play a functional role in fertilization and early embryonic development. More importantly, the newly developed antibody is able to detect the nascent and partially glycosylated precursor forms of hamster oviductin, which was unattainable with the monoclonal antibody previously used in our laboratory. Therefore, the development of the anti-rhaOvm antibody provides an additional tool for studying the intracellular behavior of hamster oviductin and will aid in elucidating the functions of hamster oviductin in the intricate process of reproduction.
Footnotes
Acknowledgements
Supported by grants from the Canadian Institutes of Health Research (MOP-44043 to FWKK and MA 11684 to GB).
The authors would like to express gratitude to Dr Yat Tse, Ms Hong Mei Gu, Ms Verna Norkum, and Mr Bob Temkin for their technical assistance and reproduction of the original photomicrographs.